Resumos
No presente estudo foram comparadas as respostas metabólicas agudas ao exercício em ratos alimentados com dieta padrão e à base de espirulina. Ratos Wistar jovens foram divididos em dois grupos de acordo com a dieta: controle (C) (dieta padrão) e espirulina (S) (dieta à base de espirulina). Ao final do período experimental (cinco semanas) os animais foram submetidos a uma sessão aguda de exercício de natação (20 minutos, suportando sobrecarga equivalente a 5% do peso corporal) para avaliação do lactato sanguíneo, glicose, insulina, proteínas, albumina e ácidos graxos livres (AGL) séricos. Amostras do músculo gastrocnêmio e fígado foram utilizadas para determinação dos teores de glicogênio e lipídeos. Ambos os grupos C e S apresentaram aumento da glicemia e dos AGL, queda da insulinemia e redução dos teores de glicogênio muscular e hepático pós-exercício. A lactacidemia durante o exercício foi superior no grupo S em relação ao C. Conclui-se que o padrão de respostas ao exercício agudo dos grupos C e S foi semelhante. Contudo, a proteína da dieta pareceu influenciar aspectos do metabolismo glicídico.
Espirulina; Metabolismo; Exercício; Ratos; Crescimento corporal
En el presente estudio fueron comparadas las respuestas metabólicas agudas al ejercicio en ratones alimentados con dieta padrón y a base de Spirulina. Ratones Wistar jovenes fueron divididos en dos grupos de acuerdo con la dieta: control (C) (dieta padrón) y Spirulina (S) (dieta a base de Spirulina). Al final del período experimental (5 semanas) los animales fueron sometidos a una sesión aguda de ejercício de natación (20 minutos, soportando sobrecarga equivalente a 5% de su peso corporal) para evaluación del lactato sanguíneo, glucosa, insulina, proteínas, albumina y ácidos grasos libres (AGL) séricos. Las muestras del músculo gastrocnemio y fígado fueron utilizadas para la determinación de los tenores de glicogeno y de lípidos. Ambos grupos C e S presentaron un aumento de la glicemia y de los AGL, mantenimiento de la insulinemia y reducción de los tenores de glicogeno muscular y hepático post-ejercicio. La lactacidemia durante el ejercicio fué superior en el grupo S en relación a C. Se concluye que el padrón de respuestas al ejercicio agudo de los grupos C e S fué semejante. Con todo, la proteína de la dieta parece influir en los aspectos del metabolismo glicídico.
Spirulina; Metabolismo; Ejercicio; Ratones; Crecimiento corporal
In this study we compared the metabolic response to acute exercise among rats fed on a standard diet or on a spirulina diet. Young Wistar rats were divided into two groups according to diet: control (C) (standard diet) and spirulina (S) (spirulina diet). At the end of experimental period (5 weeks) rats were submitted to an acute exercise session of swimming (20 minutes, supporting a load corresponding to 5% of body weight) to determine blood lactate and serum glucose, insulin, proteins, albumin and fatty free acids (FFA). Gastrocnemius and liver samples were used to determine glycogen and lipids tenors. Both C and S groups showed increase in serum glucose and FFA, a drop in serum insulin and a decrease of muscle and liver glycogen contents after acute exercise. Blood lactate during exercise was higher in S than C rats. It was concluded that the response pattern to acute exercise was similar for C and S rats. However, diet protein seemed to influence aspects of glucose metabolism.
Spirulina; Metabolism; Exercise; Rats; Body growth
ARTIGO ORIGINAL
Influência da ingestão de espirulina sobre o metabolismo de ratos exercitados
Influencia de ingestión de Spirulina sobre el metabolismo de ratones exercitados
Gustavo Puggina Rogatto; Camila Aparecida Machado de Oliveira; Júlio Wilson dos Santos; Fúlvia de Barros Manchado; Fábio Yuzo Nakamura; Camila de Moraes; Alessandro de Moura Zagatto; Marcel Cardoso Faria; Márcia Afonso; Maria Alice Rostom de Mello
Universidade Estadual Paulista, Rio Claro-SP
Endereço para correspondência Endereço para correspondência Gustavo Puggina Rogatto Rua Padre Ferraz, 427 13970-250 - Itapira, SP Tels.: (65) 9976-2499/(19) 3863-2328 e-mail: grogatto@hotmail.com
RESUMO
No presente estudo foram comparadas as respostas metabólicas agudas ao exercício em ratos alimentados com dieta padrão e à base de espirulina. Ratos Wistar jovens foram divididos em dois grupos de acordo com a dieta: controle (C) (dieta padrão) e espirulina (S) (dieta à base de espirulina). Ao final do período experimental (cinco semanas) os animais foram submetidos a uma sessão aguda de exercício de natação (20 minutos, suportando sobrecarga equivalente a 5% do peso corporal) para avaliação do lactato sanguíneo, glicose, insulina, proteínas, albumina e ácidos graxos livres (AGL) séricos. Amostras do músculo gastrocnêmio e fígado foram utilizadas para determinação dos teores de glicogênio e lipídeos. Ambos os grupos C e S apresentaram aumento da glicemia e dos AGL, queda da insulinemia e redução dos teores de glicogênio muscular e hepático pós-exercício. A lactacidemia durante o exercício foi superior no grupo S em relação ao C. Conclui-se que o padrão de respostas ao exercício agudo dos grupos C e S foi semelhante. Contudo, a proteína da dieta pareceu influenciar aspectos do metabolismo glicídico.
Palavras-chave: Espirulina. Metabolismo. Exercício. Ratos. Crescimento corporal.
RESUMEN
En el presente estudio fueron comparadas las respuestas metabólicas agudas al ejercicio en ratones alimentados con dieta padrón y a base de Spirulina. Ratones Wistar jovenes fueron divididos en dos grupos de acuerdo con la dieta: control (C) (dieta padrón) y Spirulina (S) (dieta a base de Spirulina). Al final del período experimental (5 semanas) los animales fueron sometidos a una sesión aguda de ejercício de natación (20 minutos, soportando sobrecarga equivalente a 5% de su peso corporal) para evaluación del lactato sanguíneo, glucosa, insulina, proteínas, albumina y ácidos grasos libres (AGL) séricos. Las muestras del músculo gastrocnemio y fígado fueron utilizadas para la determinación de los tenores de glicogeno y de lípidos. Ambos grupos C e S presentaron un aumento de la glicemia y de los AGL, mantenimiento de la insulinemia y reducción de los tenores de glicogeno muscular y hepático post-ejercicio. La lactacidemia durante el ejercicio fué superior en el grupo S en relación a C. Se concluye que el padrón de respuestas al ejercicio agudo de los grupos C e S fué semejante. Con todo, la proteína de la dieta parece influir en los aspectos del metabolismo glicídico.
Palabras-clave: Spirulina. Metabolismo. Ejercicio. Ratones. Crecimiento corporal.
INTRODUÇÃO
A desnutrição protéica é um problema de saúde pública que há tempos acomete grande parcela da população mundial, principalmente nos países em desenvolvimento(1-4). No Brasil, até regiões tidas como ricas como, por exemplo, a região Sudeste, apresentam significativa prevalência de desnutrição (6%)(5). Em Estados pertencentes às regiões Norte e Nordeste os índices são ainda mais preocupantes, chegando a atingir 17% da população com menos de cinco anos de idade.
O comprometimento do crescimento é uma conseqüência freqüentemente observada em organismos em fase de desenvolvimento submetidos a situações em que haja carência de nutrientes, principalmente de origem protéica(6). Contudo, outros efeitos deletérios que aumentam a taxa de mortalidade infantil e comprometem o desenvolvimento mental e a maturação demonstram o risco que a deficiência protéico-calórica pode gerar sobre crianças desnutridas(2).
Seja qual for a causa da desnutrição, o organismo gera respostas adaptativas aos longos períodos em que é submetido a esta condição. No que diz respeito à utilização de substratos energéticos, estudos relatam diminuição do gasto energético quando a ingestão de energia é reduzida(2). Esta automodulação entre ingestão e gasto é mantida até um momento em que o gasto energético não pode ser compensado pela ingestão insuficiente. Dessa forma, o organismo lança mão de suas reservas de gordura, o que resulta em diminuição da adiposidade e perda de peso. Os metabolismos de carboidratos e proteínas também sofrem influência da carência de proteínas na dieta, apresentando redução dos estoques de glicogênio e proteínas no músculo e no fígado(7-11). Ao mesmo tempo, em indivíduos desnutridos, podem ser observadas maiores concentrações de lipídeos hepáticos em função da deficiência do transporte de gorduras para fora do fígado(12,13). Estes e outros efeitos negativos apontam para a necessidade de novas alternativas para o controle e combate a este grave problema que é a desnutrição protéico-calórica (DPC).
A agricultura convencional, apesar da tecnologia disponível, possui limitações na sua capacidade de fornecer proteína necessária à população mundial(14). O problema da desnutrição protéica vem preocupando as autoridades ligadas aos setores de saúde pública há várias décadas, especialmente a Organização Mundial da Saúde (OMS), cujos técnicos têm debatido novas fontes protéicas alimentares bem como normas a serem adotadas para seu uso na alimentação humana. Nesse sentido, microorganismos têm recebido atenção especial como fonte alternativa de proteína na dieta.
Entre os vários microorganismos que vêm sendo estudados, a alga verde-azulada espirulina é considerada um microorganismo promissor, devido ao seu alto teor protéico (65 a 70% de seu peso seco) e abundância em vitaminas e minerais(14,15).
Espirulina é uma alga verde azulada de forma helicoidal, com comprimento de 0,2-0,5mm(16). A alga cresce normalmente em águas de lagos naturalmente alcalinos, localizados em zonas áridas. Embora a água desses lagos não possa ser usada para irrigação, pode ser utilizada para o cultivo de espirulina(17). Uma vez que essa alga tem rápida taxa de reprodução, dividindo-se três vezes ao dia, uma área devotada totalmente ao crescimento de espirulina pode produzir 125 vezes mais proteína que área de mesma dimensão voltada ao cultivo de milho; ou 70 vezes mais do que se voltada à criação de gado(17). Além disso, a espirulina possui algumas vantagens sobre as outras algas, dentre as quais podemos citar paladar agradável, não apresenta problemas na sua digestão e nem toxicidade aparente aos humanos, o que não acontece com outras algas tais como Chlorella e Scenedesmus(14). Apresenta, ainda, ações antioxidantes e hipocolesterolêmica(18).
Assim como a desnutrição, o exercício físico é uma condição em que são observadas modificações do perfil metabólico geral, ocorrendo mobilização das reservas de substratos energéticos e alterações nas secreções hormonais(19,20). As respostas agudas ao estímulo aeróbio por meio do exercício físico têm sido reportadas na literatura específica, registrando manutenção ou decréscimo dos estoques de substrato lipídico e glicídico durante o exercício em diferentes modelos experimentais (sedentarismo e treinamento, diabetes, obesidade) em humanos ou animais de laboratório(21-23). São escassas na literatura pesquisas sobre os efeitos da utilização de espirulina como fonte de proteínas, principalmente relacionando com exercício físico. Dessa forma, o presente estudo foi delineado para avaliar os efeitos da ingestão desta algacomo única fonte protéica alimentar sobre o crescimento somático e respostas metabólicas ao exercício aeróbio agudo de ratos jovens.
MÉTODOS
Animais
Para o desenvolvimento deste estudo foram utilizados ratos machos jovens (aproximadamente 30 dias) Wistar (Rattus norvegicus albinus, Wistar). Os animais provenientes do Biotério Central da Unesp-Botucatu foram mantidos no Biotério do Laboratório de Biodinâmica do Departamento de Educação Física do Instituto de Biociências-Unesp-Rio Claro. Os ratos foram mantidos em gaiolas coletivas (com cinco ratos por gaiola) à temperatura ambiente controlada de 25ºC e fotoperíodo de 12h claro/12h escuro (7:00/19:00h) e água ad libitum. Todos os procedimentos aos quais os animais foram submetidos estão de acordo com as normas para experimentação animal propostas pelo "International Guiding Principles for Biomedical Research Involving Animals".
Grupos experimentais
Os animais foram distribuídos aleatoriamente nos seguintes grupos experimentais:
Controle (C): Animais alimentados com dieta balanceada padrão (caseína como fonte protéica) durante cinco semanas.
Espirulina (S): Animais alimentados com dieta manipulada (espirulina como fonte protéica) durante cinco semanas.
A composição das dietas está apresentada no quadro 1.
Procedimentos
Durante todo o período experimental os animais foram pesados e medidos semanalmente. Avaliou-se a ingestão alimentar diária e calculou-se a eficiência alimentar através da seguinte equação:

Onde:
EA: eficiência alimentar
PF: peso corporal ao final do experimento (g)
PI: peso no início do experimento (g)
TA: quantidade total de alimento ingerido durante o experimento
Na última semana os animais dos dois grupos foram novamente divididos em subgrupos: CR (controle repouso); CE (controle exercício agudo); SR (espirulina repouso); SE (espirulina exercício agudo), de acordo com a realização ou não de uma sessão aguda de exercício imediatamente antes do sacrifício.
Exercício físico agudo
Após um período de adaptação ao meio líquido (cinco minutos de imersão parcial em água sem a utilização de carga, nos três dias que antecederam o teste), os animais dos grupos CE e SE foram submetidos a uma sessão de exercício físico que consistiu na realização de 20 minutos de natação em tanque com água, suportando sobrecarga equivalente a 5% do peso corporal de cada animal acoplada ao tórax.
A temperatura da água foi mantida entre 30ºC e 32ºC por ser considerada termicamente neutra em relação à temperatura corporal do rato(25).
Durante a realização da sessão de natação, foram coletadas amostras de sangue através de corte na extremidade distal da cauda dos animais em repouso e aos 5, 10, 15 e 20 minutos de exercício para a determinação das concentrações de lactato. O lactato foi determinado em analisador eletroquímico YSL 2300 STAT (Yellow Springs, Inc., EUA).
Sacrifício dos animais e análises bioquímicas
Ao final do período experimental, os animais foram sacrificados por decapitação sem jejum prévio. O sacrifício dos ratos dos grupos controle e espirulina ocorreu nas condições de repouso ou após a realização aguda de exercício de natação.
Amostras de sangue do tronco foram coletadas em tubos de vidro sem anticoagulante para avaliação da glicose (método enzimático da glicose-oxidase)(26), albumina, proteínas, ácidos graxos livres(27) e insulina sérica (radioimunoensaio - Kit Coat-A-Count, EUA).
Foi realizada uma incisão mediana na pata traseira direita para a retirada de amostra do músculo gastrocnêmio. A amostra foi pesada e utilizada para a análise das concentrações de glicogênio por método colorimétrico(28,29). Amostras do fígado foram utilizadas para dosagem do glicogênio(28,29) e lipídeos(27), também por colorimetria.
Todas as determinações bioquímicas foram efetuadas por um técnico especializado que não tinha acesso às informações quanto à alocação dos animais aos respectivos grupos.
Análise estatística
Os resultados foram expressos como média ± desvio padrão e analisados estatisticamente por teste t-Student para amostras independentes, análise de Variância e teste post-hoc de Bonferroni. Em todos os casos, o nível de significância foi prefixado em 5%.
RESULTADOS
Parâmetros avaliados durante o período experimental
Na tabela 1 encontram-se os resultados referentes ao peso corporal, comprimento corporal e ingestão alimentar dos animais dos grupos controle e espirulina durante as cinco semanas de tratamento. Ambos os grupos apresentaram significativo aumento do peso corporal após a primeira semana do estudo, não sendo observadas diferenças entre grupos. Com relação ao comprimento corporal, a partir da segunda semana os animais de ambos os grupos apresentaram crescimento significativo semanalmente até o final do experimento. Contudo, assim como o peso, o comprimento corporal não diferiu entre os grupos controle e espirulina. Não foram observadas diferenças significativas na ingestão alimentar quando comparamos os grupos. A eficiência alimentar do grupo controle também foi semelhante à do grupo espirulina (figura 1).
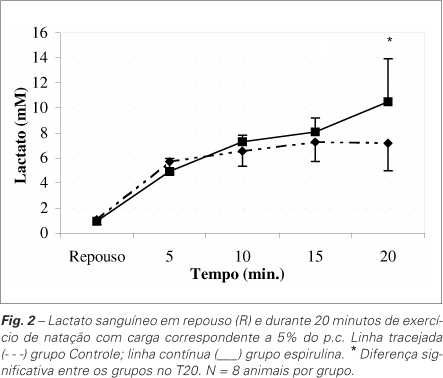
As concentrações de lactato sanguíneo estão apresentadas na figura 2. Tanto os animais do grupo controle quanto os do grupo espirulina apresentaram significativo aumento deste metabólito durante a realização do teste de carga. Contudo, nos animais do grupo espirulina este aumento foi mais acentuado no 20o minuto (p < 0,05).
Parâmetros avaliados após o sacrifício
No presente estudo, 30 animais de cada grupo foram considerados para a avaliação dos parâmetros pré-sacrifício (peso, comprimento e ingestão e eficiência alimentar). Entretanto, em função da continuidade do experimento e de outras avaliações, somente oito animais de cada grupo foram sacrificados para a obtenção de material biológico para o presente estudo.
Na tabela 2 estão dispostas as concentrações séricas de glicose, insulina, proteínas totais, albumina e ácidos graxos livres (AGL) dos animais dos grupos controle e espirulina nas condições de repouso e após a realização de uma sessão de 20 minutos de exercício de natação. Foi observado um aumento da glicemia pelo efeito agudo do esforço, sendo este menos evidente no grupo espirulina. A sessão única de exercício resultou em redução das concentrações séricas de insulina em ambos os grupos experimentais. Não foram observadas diferenças significativas nas concentrações de proteínas tanto pela ação da dieta, quanto pela realização do exercício agudo. Ao término da série de esforço, ambos os grupos experimentais apresentaram modificações da albumina do soro, sendo que no grupo controle foi observada redução e no grupo espirulina, aumento. No que diz respeito aos ácidos graxos livres, somente o exercício agudo causou alterações significativas, resultando em aumento deste substrato lipídico.
Os teores de glicogênio hepático e muscular e as concentrações de lipídeos do fígado estão registrados na tabela 3. Tanto o conteúdo de carboidrato do fígado quanto do músculo apresentaram comportamentos semelhantes pela realização do exercício agudo e pela modificação de dieta. Após a realização da série de natação, ambos os órgãos apresentaram significativa queda das reservas deste substrato energético, sendo que no grupo espirulina esta redução foi mais acentuada. Não foram observadas modificações nos teores de lipídeos hepáticos.
DISCUSSÃO
A utilização de espirulina como alimento tem sido bastante discutida na literatura específica(14,30-32). Além de ser boa fonte de proteínas, minerais e vitaminas, a espirulina não apresenta toxicidade ao organismo, além de não comprometer o desenvolvimento de órgãos e tecidos(14,31). Em nosso estudo, não observamos alterações no peso e no comprimento corporal dos animais alimentados com dieta à base de espirulina quando comparados com o grupo controle. Além disso, os valores de ingestão e eficiência alimentar foram também semelhantes entre as duas condições nutricionais (controle e espirulina), indicando que esta fonte alternativa de proteína proveniente de biomassa não compromete o crescimento de animais em fase de desenvolvimento. Estes resultados corroboram relatos da literatura que compararam dietas à base de espirulina e caseína(14) e espirulina e "pellet"(30). A utilização de animais jovens (30 dias, no início do experimento) pode explicar a redução da quantidade de alimento ingerido ao longo do estudo, uma vez que à medida que estes envelheceram, a necessidade de nutrientes para o crescimento somático pode ter sido diminuída. Através das avaliações feitas no presente estudo, não foi possível determinar as razões que levaram os animais do grupo controle a apresentarem redução da quantidade de alimento ingerido ao longo do experimento antecipadamente aos animais do grupo espirulina.
Em nosso trabalho, a realização de exercício físico promoveu alterações das concentrações de lactato sanguíneo em ambos os grupos, indicando uma condição de estresse metabólico agudo. Durante a realização do exercício físico ocorre um aumento da exigência de diversos sistemas orgânicos, o que resulta na ativação de mecanismos relacionados com a mobilização de substratos energéticos. Tal resposta facilita a redistribuição destes combustíveis no organismo, visando a manutenção do trabalho muscular. Quando esta demanda excede o suprimento de oxigênio ou sua taxa de utilização, a contribuição energética pela via anaeróbia do metabolismo de carboidratos torna-se maior, o que produz um excesso de íons hidrogênio que se combinam com o ácido pirúvico e formam ácido lático. Posteriormente, este se difunde para o sangue, sendo convertido em lactato(33). O lactato produzido tem importante papel no fornecimento de energia, já que pode ser reconvertido em glicose pelo fígado(34). As concentrações de lactato sanguíneo durante os primeiros 15 minutos de natação (teste agudo) foram semelhantes entre os grupos controle e espirulina (figura 2). Apenas no 20o minuto houve uma elevação mais pronunciada no grupo alimentado com dieta à base de espirulina. Provavelmente, os grupos exercitaram-se em domínios de esforço diferentes. Segundo Gaesser e Poole(35), as intensidades de exercício em que se detecta uma elevação da concentração de lactato sanguíneo acima dos valores de repouso, porém, com estabilização em algum momento da carga, pertencem ao domínio intenso de esforço. O limite superior desse domínio é a intensidade correspondente ao máximo estado estável de lactato, ou limiar anaeróbio. As intensidades de esforço superiores ao limiar anaeróbio provocariam um aumento contínuo da concentração sanguínea de lactato, precipitando a fadiga decorrente da acidose metabólica. Essas intensidades são enquadradas no domínio severo de esforço. Acredita-se que nesse domínio de esforço o  atingiria invariavelmente seu valor máximo. Dessa forma, de acordo com os resultados da figura 2, poderíamos supor que os ratos pertencentes ao grupo controle estariam se exercitando no domínio intenso de esforço. Ou seja, a natação com sobrecarga adicional de 5% do peso corporal não seria suficientemente intensa a ponto de provocar uma acidose elevada, tampouco uma resposta cardiorrespiratória máxima (inferida pela constatação do
atingiria invariavelmente seu valor máximo. Dessa forma, de acordo com os resultados da figura 2, poderíamos supor que os ratos pertencentes ao grupo controle estariam se exercitando no domínio intenso de esforço. Ou seja, a natação com sobrecarga adicional de 5% do peso corporal não seria suficientemente intensa a ponto de provocar uma acidose elevada, tampouco uma resposta cardiorrespiratória máxima (inferida pela constatação do  ). Já os animais pertencentes ao grupo espirulina estariam, durante o teste agudo, realizando exercício severo, pois não há indicação de estabilização do lactato sanguíneo, particularmente nos cinco minutos finais de esforço, quando a concentração do metabólito sofreu elevação de 2,4mM. Autores(36) aceitam que uma elevação inferior a 1mM durante os últimos 20 minutos de carga retangular de 30 minutos seria um bom indicador de ocorrência de estado estável de lactato. Segundo essa interpretação, os animais alimentados com espirulina teriam um prejuízo no desenvolvimento de sua capacidade aeróbia em comparação com os do grupo controle no decorrer de seu crescimento e desenvolvimento, pois, ao serem submetidos a uma mesma carga relativa de nado, apresentam respostas fisiológicas que indicam maior sobrecarga orgânica (acidose).
). Já os animais pertencentes ao grupo espirulina estariam, durante o teste agudo, realizando exercício severo, pois não há indicação de estabilização do lactato sanguíneo, particularmente nos cinco minutos finais de esforço, quando a concentração do metabólito sofreu elevação de 2,4mM. Autores(36) aceitam que uma elevação inferior a 1mM durante os últimos 20 minutos de carga retangular de 30 minutos seria um bom indicador de ocorrência de estado estável de lactato. Segundo essa interpretação, os animais alimentados com espirulina teriam um prejuízo no desenvolvimento de sua capacidade aeróbia em comparação com os do grupo controle no decorrer de seu crescimento e desenvolvimento, pois, ao serem submetidos a uma mesma carga relativa de nado, apresentam respostas fisiológicas que indicam maior sobrecarga orgânica (acidose).
A resposta glicêmica durante e após a realização aguda de exercício em ratos tem sido bastante variada em função das diferenças nas atividades empregadas. O desempenho de exercício de natação com cargas que variaram de 0 a 8% em relação ao peso corporal por 30 ou 60 minutos, em alguns casos não resultou em modificações no perfil glicêmico na condição aguda(37-39). Reduções da glicemia também têm sido observadas após a realização aguda de esforço aeróbio como reportado por Nardo e Luciano(40). Nestes casos o aumento da captação de glicose pelos tecidos periféricos durante o exercício pode ser o fator responsável pela queda glicêmica(41). Por outro lado, em nosso trabalho verificamos aumentos das concentrações séricas de glicose, o que coincide com achados anteriores(19,20,37). Esta resposta hiperglicêmica foi observada tanto no grupo controle, quanto no espirulina e possivelmente está relacionada com o aumento da atividade simpática com conseqüente aumento da atividade glicogenolítica muscular e hepática. Mecanismos envolvidos com a neoglicogênese também podem estar contribuindo com o incremento do aporte glicêmico em função de elevações na secreção de hormônios glicocorticóides(42). Além disso, vários estudos têm reportado aumentos da resposta secretória de outros hormônios contra-regulatórios como hormônio do crescimento, catecolaminas, entre outros, após exercício agudo(43-50).
A baixa na concentração sérica de insulina induzida pela realização aguda de esforço, observada em ambos os grupos de nosso estudo, está de acordo com trabalhos anteriores que utilizaram exercícios contínuos e intermitentes(19,20,48). A estimulação simpática decorrente da realização aguda de esforço pode ter resultado na inibição da liberação insulínica, o que, somado ao aumento da secreção dos hormônios contra-regulatórios, contribuiu com a elevação da glicemia em ambos os grupos experimentais.
O metabolismo lipídico também pode sofrer interferência da atividade física, já que a mobilização deste tipo de substrato pode encontrar-se aumentada. Em nosso estudo as concentrações de AGL apresentaram significativo aumento após a realização da sessão única de esforço tanto no grupo alimentado com dieta padrão, quanto no com espirulina. Este resultado pode indicar aumento da mobilização dos triglicerídeos estocados nos adipócitos, o que coincide com trabalhos anteriores(51) e pode estar relacionado com elevações das secreções dos hormônios contra-regulatórios. A qualidade da proteína ingerida parece não influenciar as concentrações deste substrato lipídico.
A avaliação das proteínas totais e da albumina sérica pode ser utilizada como indicador da presença de desnutrição protéica, já que organismos com Kwashiorkor apresentam significativa redução destes componentes(4). Dessa forma, o fato de não termos constatado redução nas proteínas sanguíneas no grupo alimentado com a dieta espirulina é mais uma evidência de que esta última consiste numa fonte protéica adequada ao desenvolvimento de animais jovens. Foram observadas influências da atividade física aguda sobre o perfil da albumina, que sofreram redução e aumento nos grupos controle e espirulina, respectivamente. Uma hipótese para este tipo de resposta ao exercício pode estar relacionada a modificações da atividade catabólica do organismo frente à atividade aguda, geradas pela ingestão de espirulina, no sentido de acelerar a mobilização das proteínas musculares e hepáticas, aumentando a disponibilidade de albumina na circulação.
A esteatose hepática, ou o acúmulo de lipídeos no fígado é uma alteração metabólica que também tem sido observada em organismos submetidos à desnutrição protéico-calórica crônica(13,52). Este aumento da concentração de gordura no fígado acontece principalmente em função de incremento dos triglicerídeos(13,53). Em nosso estudo, não observamos qualquer diferença nos teores hepáticos de lipídeos em ambas as condições nutricionais e de exercício. Em estudos anteriores foi relatado aumento na atividade da enzima lipaselipoprotéica no fígado de animais alimentados com espirulina(54).
O metabolismo intermediário tem importante papel na manutenção da homeostase orgânica, contribuindo com o fornecimento de energia em condições normais ou em situações em que a exigência metabólica encontra-se aumentada. A atividade física aguda é um tipo de estimulo que exige do organismo maior mobilização de substrato energético, inclusive das reservas de carboidratos(33,55). Em nosso estudo observamos que as reservas de glicogênio hepático e muscular apresentaram significativa redução após a realização do exercício agudo tanto no grupo submetido à dieta controle, quanto no alimentado com espirulina. Este tipo de comportamento metabólico tem sido freqüentemente observado em vários estudos que utilizaram diferentes formas de atividades físicas(16). Este decréscimo dos estoques de glicogênio está relacionado com a elevação da atividade glicogenolítica causada pelo aumento agudo dos hormônios catabólicos(56) e foi responsável pelo acréscimo da glicemia pós-exercício em ambos os grupos. Não temos explicações consistentes para a observação de maior mobilização de glicogênio muscular e hepático nos animais do grupo espirulina submetido ao exercício agudo, mas mecanismos relacionados com a glicogenólise, como a atividade das enzimas glicogenolíticas bem como atividade adrenérgica, que podem estar aumentadas no grupo alimentado com esta alga, são possíveis explicações para tal resposta. Os estoques de glicogênio muscular e hepático, significativamente maiores nos animais alimentados com espirulina em repouso em comparação com os controles, podem ser de fundamental importância na manutenção da homeostase durante o exercício nesses animais. O conjunto desses resultados aponta para a necessidade de novas pesquisas, a fim de esclarecer pontos ainda obscuros sobre os efeitos da utilização de espirulina como fonte alternativa de proteína para organismos em fase de crescimento.
Concluímos em nosso estudo que a espirulina não causou prejuízo no crescimento dos roedores, bem como na maioria dos parâmetros bioquímicos analisados, e apresentou boa aceitação pelos animais. Isso sugere que esta alga possa ser uma boa fonte protéica alternativa. Entretanto, os elevados teores de lactato sanguíneo observados durante o teste de esforço podem induzir a fadiga precoce no exercício, o que poderia comprometer o desempenho físico. Mais estudos sobre o tema são necessários para elucidar essa e outras questões que permanecem sem resposta.
Recebido em 6/2/04. 2a versão recebida em 26/4/04. Aceito em 2/5/04
Todos os autores declararam não haver qualquer potencial conflito de interesses referente a este artigo.
- 1. Castro J. Geopolítica da fome. 7a ed. Rio de Janeiro: Brasiliense, 1965.
- 2. Torun B, Chew F. Protein energy malnutrition. In: Shils M, Olson JA, Shike M, editors. Modern nutrition in health and disease. Philadelphia: Lea & Febiger, 1994;950-76.
- 3. Uvin P. The state of world hunger. Nutr Res 1994;52:151-61.
- 4. Dâmaso A. Nutrição e exercício na prevenção de doenças. Rio de Janeiro: MEDSI, 2001.
-
5Ministério da Saúde. Pesquisa nacional sobre demografia e saúde 1996: relatório preliminar. Brasília: Ministério da Saúde, 1996.
- 6. Frank S. Metabolic adaptation in protein-energy malnutrition. J Am Coll Nutr 1986;5:371-81.
- 7. Alleyne GAO, Scullard GH. Alterations in carbohydrate metabolism in Jamaican children with severe malnutrition. Clin Sci 1969;37:631-42.
- 8. Alleyne GAO, Millward DJ, Scullard GH. Total body potassium muscle electrolytes and glycogen in malnourished children. J Pediatr 1970;76:75-81.
- 9. Waterlow JC, Mendes CB. Composition of muscle in malnourished infants. Nature 1957;180:1361.
- 10. Waterlow JC. The nature and significance of nutritional adaptation. Eur J Clin Nutr 1998;53:52-5.
- 11. Waterlow JC, Weisz J. The fat protein and nucleic acid of liver in malnourished human infant. J Clin Invest 1956;35:346-54.
- 12. Flores H, Pak N, Maccioni A, Monckeberg F. Lipid transport in kwashiorkor. Br J Nutr 1970;24:1005-11.
- 13. Kumar V, Deo MG, Ramalingaswami V. Mechanism of fatty liver in protein deficience: an experimental study in the rhesus monkey. Gastroenterology 1972; 62:445-51.
- 14. Contreras A, Herbert DC, Grubbs BG, Cameron IL. Blue-green alga, spirulina, as the sole dietary source of protein in sexually maturing rats. Nutr Rep Int 1979; 19:749-63.
- 15. Kay RA. Microalgae as food and supplement. Food Sci Nutr 1991;30:555-73.
- 16. Hadenshog G, Haffen AV. The ultrastructure of Spirulina plantensis. Physiol Plant 1970;23:209.
- 17. Furst PT. Spirulina a nutricious alga, once a staple of Aztec diet, could feed many of the world hungry people. Human Nature 1978;3:60.
- 18. Rodriguez-Hernandez A, Blé-Castillo JL, Juárez-Oropeza MA, Diaz-Zagoya JC. Spirulina maxima prevents fatty liver formation in CD-1 male and female mice with experimental diabetes. Life Sciences 2001;69:1029-37.
- 19. Rogatto GP, Luciano E. Efeitos do treinamento físico intenso sobre o metabolismo de carboidratos. Revista Brasileira de Atividade Física & Saúde 2001;6:39-46.
- 20. Rogatto GP, Luciano E. Influência do treinamento físico intenso sobre o metabolismo de proteínas. Motriz 2001;7:75-82.
- 21. Green HJ, Ball-Brunett M, Symon S, Grant S, Jamieson G. Short-term training, muscle glycogen and cycle endurance. Can J Appl Physiol 1995;20:315-24.
- 22. Nakamura M, Brown J, Miller WC. Glycogen depletion patterns in trained rats adapted to a high-fat or high-carbohydrate diet. Int J Sports Med 1998;19:419-24.
- 23. Romijn JA, Coyle EF, Sidossis LS, Rosenblatt J, Wolfe RR. Substrate metabolism during different exercise intensities in endurance-trained women. J Appl Physiol 2000;88:1707-14.
- 24. Reeves PG, Nielsen FH, Fahey GC Jr. AIN-93 purified diets for laboratory rodents: final report of the American Institute of Nutrition ad hoc writing committee on the reformulation of the AIN-76 rodent diet. J Nutr 1993;123:1939-51.
- 25. Azevedo JRM. Determinação de parâmetros bioquímicos em ratos sedentários e treinados, durante e após exercício agudo de natação. Tese de Doutorado. Campinas: Unicamp, 1994.
- 26. Henry RJ, Cannon DC, Wilkeman J. Clinical chemistry, principles and techniques. 2nd ed. New York: Harper and Harper Row Publishes, 1974.
- 27. Nogueira DM, Strufaldi B, Hirata MH, Abdalla DSP, Hirata RDC. Métodos de bioquímica clínica: técnico-interpretação. São Paulo: Pancasat, 1990.
- 28. Sjörgreen B, Nordenkjold T, Holmgren H, Wollerstrom J. Bertrag zur kuntnis des leberrhythmik. Pflügers Arch Gesamte Physiol Menschen Tiere 1938;240:247.
- 29. Dubois B, Gilles KA, Hamilton JK, Rebers PA. Colorimetric method for determination of sugar and related substances. Anal Chem 1965;28:350-6.
- 30. Tranquille N, Emeis JJ, Chambure D, Binot R, Tamponnet C. Spirulina acceptability trials in rats. A study for the "Melissa" life-support system. Adv Space Res 1994;14:167-70.
- 31. Salazar M, Martinez E, Madrigal E, Ruiz LE, Chamorro GA. Subchronic toxicity study in mice fed Spirulina maxima. J Ethnophamacol 1998;62:235-41.
- 32. Grinstead GS, Tokach MD, Dritz SS, Goodband RD, Nelssen JL. Effects of Spirulina plantensis on growth performance of weanling pigs. Anim Feed Sci Technol 2000;83:237-47.
- 33. McArdle WD, Katch FI, Katch VL. Fisiologia do exercício: energia, nutrição e desempenho humano. Rio de Janeiro: Guanabara Koogan, 1996.
- 34. Brooks GA. Ácido láctico no sangue: o "vilão" dos esportes torna-se bom. Sports Science Exchange 1995;2:1-6.
- 35. Gaesser GA, Poole DC. The slow component of oxygen uptake kinetics in humans. Exerc Sport Sci Rev 1986;24:35-70.
- 36. Heck H, Mader A, Hess G, Mücke S, Hollman W. Justification of the 4-mmol/l lactate threshold. Int J Sports Med 1985;6:117-30.
- 37. Prada FJA, Carneiro EM, Azevedo JRM, Luciano E. Respostas endócrino-metabólicas em ratos diabéticos: efeito estressor do exercício agudo. Revista Brasileira de Atividade Física & Saúde 1997;2:22-9.
- 38. Felício PFV, Drigo AJ, Azevedo JRM. Efeito da administração de ascorbato (vitamina C) associada ao exercício agudo de natação sobre os níveis de substratos energéticos em cobaias. Motriz 1999;5:92.
- 39. Luciano E, Guerino MR, Luciano EA. Influências da atividade física e da aplicação do ultra-som sobre o reparo ósseo em ratos diabéticos. Rev Bras Cien Mov (Edição Especial) 1999;142.
- 40. Nardo N Jr, Luciano E. Influências do treinamento físico sobre a resposta ao estresse agudo em ratos normais e diabéticos experimentais. Anais do XXI Simpósio Internacional de Ciências do Esporte. p. 105. São Paulo: CELAFISCS, 1998.
- 41. Luciano E, Carneiro EM, Reis MAB, Peres SB, Velloso LA, Boschero AC, Saad MJA. Endurance training modulates early steps of insulin signaling in rat muscle. Med Sci Sports Exerc 1998;30:S24.
- 42. Rogatto GP, Luciano E. Hormonal and metabolic response to acute exercise: effects of high intensity training. Med Sci Sports Exerc 2000;32:S227.
- 43. Jurimae T, Karelson K, Smirnova T, Viru A. The effect of a single-circuit weight-training session on the blood biochemistry of untrained university students. Eur J Appl Physiol 1990;61:344-8.
- 44. Tabata I, Atomi Y, Mutoh Y, Miyashita M. Effect of physical training on the responses of serum adrenocorticotropic hormone during prolonged exhausting exercise. Eur J Appl Physiol 1990;61:188-92.
- 45. Häkkinen K, Pakarinen A. Acute hormonal response to heavy resistance loading in men and women at different ages. Int J Sports Med 1995;16:507-13.
- 46. Gotshalk LA, Loebel CC, Nindl BC, Putukian M, Sebastianelli WJ, Newton RU, et al. Hormonal responses of multiset versus single-set heavy-resistance exercise protocols. Can J Appl Physiol 1997;22:244-55.
- 47. Ortega E, Rodriguez MJ, Barriga C, Forner MA. Corticosterone, prolactin and thyroid hormones as hormonal mediators of the stimulated phagocytic capacity of peritoneal macrophages after high-intensity exercise. Int J Sports Med 1996; 17:149-55.
- 48. Kraemer WJ, Hakkinen K, Newton RU, McCormick M, Nindl BC, Volek JS, et al. Acute hormonal responses to heavy resistance exercise in younger and older men. Eur J Appl Physiol 1998;77:206-11.
- 49. Viru A, Laaneots L, Karelson K, Smirnova T, Viru M. Exercise-induced hormone responses in girls at different stages of sexual maturation. Eur J Appl Physiol 1998;77:401-8.
- 50. Zouhal H, Rannou F, Gratas-Delamarche A, Monnier M, Bentué-Ferrer D, Delamarche P. Adrenal medulla responsiveness to the sympathetic nervous activity in sprinters and untrained subjects during a supramaximal exercise. Int J Sports Med 1998;19:172-6.
- 51. Stevanato E. Efeitos do jejum sobre a interação entre o metabolismo de ácidos graxos livres e glicose em músculos esqueléticos de ratos treinados. Dissertação de Mestrado. Rio Claro: Unesp, 1999.
- 52. Ferro-Luzzi A, Spadoni MA. Protein-energy malnutrition. Prog Food Nutr Sci 1978; 2:515-41.
- 53. Taylor GO, Ziboh VA. Liver lipid changes in experimental protein malnutrition. Am J Clin Nutr 1972;25:285-90.
- 54. Iwata K, Inayama T, Kato T. Effects of Spirulina plantensis on plasma lipoprotein lipase activity in fructose-induced hyperlipidemic rats. J Nutr Sci Vitaminol 1990; 36:165-71.
- 55. Fox EL, Bowers RW, Foss ML. Bases fisiológicas de educação física e dos desportos. 4a ed. Rio de Janeiro: Guanabara Koogan, 1991.
- 56. Guyton AC, Hall JE. Tratado de fisiologia médica. 9a ed. Rio de Janeiro: Guanabara Koogan, 1997.
Datas de Publicação
-
Publicação nesta coleção
09 Nov 2004 -
Data do Fascículo
Ago 2004
Histórico
-
Aceito
02 Maio 2004 -
Revisado
26 Abr 2004 -
Recebido
06 Fev 2004