Resumos
O treinamento esportivo provoca adaptações neuromusculares e alterações metabólicas visando a performance durante a competição. Nas competições de judô, o número de lutas a que os atletas são submetidos e suas respectivas durações e intervalos são aleatórios, fatores que podem influenciar a performance objetivada no treinamento. O presente estudo investigou a hipótese de que diferentes durações de lutas, 90s, 180s e 300s, poderiam influenciar a atividade enzimática, elétrica muscular e a produção do pico de torque. Antes e após cada luta, foram coletadas amostras sanguíneas dos atletas; em seguida, os mesmos realizaram cinco contrações dinâmicas (90º/s) com a utilização de um dinamômetro isocinético (Biodex System 3). Simultaneamente registrou-se o sinal eletromiográfico dos músculos agonista, antagonista e sinergista do movimento avaliado. Não se verificou alteração no torque. As enzimas AST e ALT apresentaram aumento na atividade, nas lutas de 90s (p = 0,0033/p = 0,00059), 180s (p = 0,0044/p = 0,0033) e 300s (p = 0,0044/p = 0,0033). Aumento (p = 0,0180) da atividade da CK após a luta de 300s foi verificado. A LDH diminuiu após a luta de 90s (p = 0,0392). Na análise intermuscular observou-se após a luta de 90s aumento do sinal eletromiográfico do agonista (p = 0,005); na luta de 180s, aumento do antagonista (p = 0,0129) e na luta de 300s, diminuição (p = 0,0137) da atividade do músculo agonista. Observou-se que os esforços da luta de 300s podem ter induzido lesões no tecido muscular caracterizadas pela elevação da CK plasmática, embora a lesão não tenha sido suficiente para detectar fadiga através da dinamometria isocinética. Conclui-se que o protocolo proposto foi suficiente para alteração enzimática e eletromiográfica, sugerindo adaptações metabólicas e neurais a partir do estresse das lutas de judô.
Eletromiografia; Enzimas; Fadiga; Torque; Suporte Financeiro: UNIVAP
El entrenamiento deportivo provoca adaptaciones neuromusculares y alteraciones metabólicas que buscan una actuación durante la competición. En las competencias de judo, el número de luchas a que los atletas se someten así como la duración respectiva y los intervalos son al azar, factores estos que influencian en la actuación y en el entrenamiento. El estudio presente investigó la hipótesis de que las duraciones diferentes de luchas, 90s 180s y 300s, podrían influir en la actividad enzimático, eléctrico muscular y la producción del pico del cambio. Antes y después de cada lucha, fueron obtenidas las muestras sanguíneas de los atletas, tan pronto después de que los mismos lograron cinco reducciones dinámicas (90º/s) con el uso de un dinamômetro isocinético (Biodex Sistema 3). Simultáneamente se consignaron la señal electromiográfica de los músculos agonistas, antagonistas y sinérgica de los movimientos estimados. El estudio no verificó si hubo una alteración en el torque. Las enzimas AST y ALT presentaran aumento en la actividad, en las luchas de 90s (p = 0,0033/p = 0,00059), 180s (p = 0,0044/p = 0,0033) y 300s (p = 0,0044/p = 0,0033). Un aumento (p = 0,0180) de la actividad de CK después de que la lucha de 300s fue verificada. LDH disminuyó después de la lucha de 90s (p = 0,0392). En el análisis intermuscular se observó después de la lucha 90s un aumento de la señal electromiográfica del agonista (p = 0,005), en la lucha 180s un aumento de la del antagonista (p = 0,0129) y en la lucha 300s una disminución (p = 0,0137) de la actividad del músculo agonista. Fue observado, además, que los esfuerzos de la lucha de 300s podrían haber inducido lesiones en el tejido muscular caracterizado por una elevación de CK plasmático, aunque la lesión no ha sido bastante para descubrir la fatiga a través de la dinamometría isocinética. Ha mostrado que el protocolo propuesto era bastante para determinar las alteraciones enzimáticas y eletromiográficas, mientras hace pensar en las adaptaciones metabólicas y neurológicas que ocurren por la tensión de las luchas del judo.
Electromiografía; Enzimas; Fatiga; Cambio; Apoyo financiero: UNIVAP
The sportive training causes neuromuscular adaptations and metabolic alterations aiming the competition performance. In judo competitions, the number of fights to what athletes are submitted, as well as their respective endurance and intervals are randomized, and these factors may influence the aimed training performance. This study investigated the hypothesis that different fight endurances, 90s, 180s, and 300s could influence the enzymatic and muscular electrical activity, as well as the torque peak production. Before and after each fighting, a blood sampling was collected from each athlete. After and before each fight, they performed five dynamic contractions (90º/s) using an isokinetic Dynamometer (Biodex System 3). Simultaneously, it was recorded the electromyographic signal of the agonist, antagonist and synergistic muscles of the movement assessed. It was observed no alterations in the torque. The AST and ALT enzymes presented an increasing activity in the 90 sec. (p = 0.0033/p = 0.00059), 180 sec. (p = 0.0044/p = 0.0033), and in the 300 sec. (p = 0.0044/p = 0.0033) fights. It was verified an increase (p = 0.0180) in the CK activity after the 300 sec. fight. LDH decreased after the 90 sec. fight (p = 0.0392). Upon the intermuscular analysis, it was observed an increase in the electromyographic signal of the agonist muscle after the 90 sec. fight (p = 0.005), an increase of the antagonist muscle in the 180 sec. fight (p = 0.0129), and a decrease (p = 0.0137) in the activity of the agonist muscle in the 300 sec. fight. It was observed that the strength in the 300 sec. fight might reduced the injuries in the muscular tissue characterized by a raise in the plasmatic CK, although the injury was not sufficient to detect the fatigue through the isokinetic dynamometry. It can be concluded that the proposed protocol was sufficient to the enzymatic and electromyographic alteration, suggesting metabolic and neural adaptations from stress caused by the judo fights.
Electromyography; Enzymes; Fatigue; Torque; Financial support: UNIVAP
ARTIGO ORIGINAL
Efeitos de diferentes esforços de luta de judô na atividade enzimática, atividade elétrica muscular e parâmetros biomecânicos de atletas de elite
Los efectos de diferentes esfuerzos de lucha del judo en la actividad enzimática, actividad eléctrica muscular y biomecánica de los parámetros de atletas de élite
Silvia Regina RibeiroI; Carlos Julio Tierra-CriolloII; Rodrigo Álvaro Brandão Lopes MartinsIII
ILaboratório de Biodinâmica do Movimento Humano, Faculdade de Ciências da Saúde, Universidade do Vale do Paraíba UNIVAP, São José dos Campos, SP, Brasil, Laboratório de Fisiologia e Farmacodinâmica Instituto de Pesquisa e Desenvolvimento II IP&D II, Universidade do Vale do Paraíba, São José dos Campos, SP, Brasil
IIGrupo de Engenharia Biomédica (GENEBIO), Cento de Pesquisa e Desenvolvimento em Engenharia Elétrica, Universidade Federal de Minas Gerais, Belo Horizonte, MG, Brasil
IIIDepartamento de Farmacologia, Instituto de Ciências Biomédicas I. Universidade de São Paulo, Cidade Universitária, Butantã, São Paulo, SP, Brasil
Endereço para correspondência Endereço para correspondência: Silvia Regina Ribeiro Instituto de Pesquisa & Desenvolvimento, Universidade do Vale do Paraíba UNIVAP, Av. Shishima Hifumi, 2.911, Urbanova 12244-000 São José dos Campos, SP Tel.: (12) 3947-1000 (2032) E-mail: sribeiro@univap.br
RESUMO
O treinamento esportivo provoca adaptações neuromusculares e alterações metabólicas visando a performance durante a competição. Nas competições de judô, o número de lutas a que os atletas são submetidos e suas respectivas durações e intervalos são aleatórios, fatores que podem influenciar a performance objetivada no treinamento. O presente estudo investigou a hipótese de que diferentes durações de lutas, 90s, 180s e 300s, poderiam influenciar a atividade enzimática, elétrica muscular e a produção do pico de torque. Antes e após cada luta, foram coletadas amostras sanguíneas dos atletas; em seguida, os mesmos realizaram cinco contrações dinâmicas (90o/s) com a utilização de um dinamômetro isocinético (Biodex System 3). Simultaneamente registrou-se o sinal eletromiográfico dos músculos agonista, antagonista e sinergista do movimento avaliado. Não se verificou alteração no torque. As enzimas AST e ALT apresentaram aumento na atividade, nas lutas de 90s (p = 0,0033/p = 0,00059), 180s (p = 0,0044/p = 0,0033) e 300s (p = 0,0044/p = 0,0033). Aumento (p = 0,0180) da atividade da CK após a luta de 300s foi verificado. A LDH diminuiu após a luta de 90s (p = 0,0392). Na análise intermuscular observou-se após a luta de 90s aumento do sinal eletromiográfico do agonista (p = 0,005); na luta de 180s, aumento do antagonista (p = 0,0129) e na luta de 300s, diminuição (p = 0,0137) da atividade do músculo agonista. Observou-se que os esforços da luta de 300s podem ter induzido lesões no tecido muscular caracterizadas pela elevação da CK plasmática, embora a lesão não tenha sido suficiente para detectar fadiga através da dinamometria isocinética. Conclui-se que o protocolo proposto foi suficiente para alteração enzimática e eletromiográfica, sugerindo adaptações metabólicas e neurais a partir do estresse das lutas de judô.
Palavras-chave: Eletromiografia. Enzimas. Fadiga. Torque. Suporte Financeiro: UNIVAP.
RESUMEN
El entrenamiento deportivo provoca adaptaciones neuromusculares y alteraciones metabólicas que buscan una actuación durante la competición. En las competencias de judo, el número de luchas a que los atletas se someten así como la duración respectiva y los intervalos son al azar, factores estos que influencian en la actuación y en el entrenamiento. El estudio presente investigó la hipótesis de que las duraciones diferentes de luchas, 90s 180s y 300s, podrían influir en la actividad enzimático, eléctrico muscular y la producción del pico del cambio. Antes y después de cada lucha, fueron obtenidas las muestras sanguíneas de los atletas, tan pronto después de que los mismos lograron cinco reducciones dinámicas (90o/s) con el uso de un dinamômetro isocinético (Biodex Sistema 3). Simultáneamente se consignaron la señal electromiográfica de los músculos agonistas, antagonistas y sinérgica de los movimientos estimados. El estudio no verificó si hubo una alteración en el torque. Las enzimas AST y ALT presentaran aumento en la actividad, en las luchas de 90s (p = 0,0033/p = 0,00059), 180s (p = 0,0044/p = 0,0033) y 300s (p = 0,0044/p = 0,0033). Un aumento (p = 0,0180) de la actividad de CK después de que la lucha de 300s fue verificada. LDH disminuyó después de la lucha de 90s (p = 0,0392). En el análisis intermuscular se observó después de la lucha 90s un aumento de la señal electromiográfica del agonista (p = 0,005), en la lucha 180s un aumento de la del antagonista (p = 0,0129) y en la lucha 300s una disminución (p = 0,0137) de la actividad del músculo agonista. Fue observado, además, que los esfuerzos de la lucha de 300s podrían haber inducido lesiones en el tejido muscular caracterizado por una elevación de CK plasmático, aunque la lesión no ha sido bastante para descubrir la fatiga a través de la dinamometría isocinética. Ha mostrado que el protocolo propuesto era bastante para determinar las alteraciones enzimáticas y eletromiográficas, mientras hace pensar en las adaptaciones metabólicas y neurológicas que ocurren por la tensión de las luchas del judo.
Palabras-clave: Electromiografía. Enzimas. Fatiga. Cambio. Apoyo financiero: UNIVAP.
INTRODUÇÃO
O treinamento físico visa à manutenção da performance esportiva durante o período competitivo. A plasticidade do tecido muscular esquelético permite sua adaptação a vários estados de demandas funcionais(1), o que com o treinamento corresponde ao aumento da tolerância ao exercício desencadeando processos adaptativos de caráter mecânico, metabólico e eletrofisiológico(2) de acordo com as especificidades de cada modalidade. A fadiga, caracterizada pela incapacidade de manter o torque, é um fator limitante para a performance nas competições.
A mensuração de variáveis indicativas de fadiga no esporte tem sido fortemente investigada, entre elas os níveis séricos plasmáticos de enzimas musculares utilizados como indicadores do estado de lesão e/ou prejuízo do tecido muscular, após períodos de treinamento(3,4), ou indicadores de intensidades de exercício(5,6).
As alterações na atividade elétrica das fibras musculares submetidas a contrações isométricas em protocolos de fadiga têm sido investigadas(7-9). Porém, protocolos com contrações dinâmicas, principalmente na área esportiva, necessitam de outros estudos(10-12), quando se pretende investigar as adaptações eletromiográficas do gesto esportivo a treinamentos específicos(13-17), a performance(18) e análise quantitativa da fadiga muscular(17).
Apesar de a fadiga ser uma experiência comum na área esportiva, o processo envolvido neste mecanismo ainda precisa de maiores estudos devido à divergência de resultados principalmente na análise dos sinais eletromiográficos(13,19).
A performance do gesto esportivo a priori é dependente do treino de força. Uma otimização da força objetiva ativações musculares agonistas sem ou com baixas co-ativações antagonistas(14,20,21), de forma a poupar gastos energéticos desnecessários. Dessa forma, a co-ativação muscular sugere debilidade na performance de ativação dos músculos para a realização da ação motora(7), sugerindo prejuízos na performance funcional(17).
Numa competição de judô, o número de lutas a que os atletas são submetidos e suas respectivas durações e intervalos são aleatórios, fatores que podem influenciar a performance objetivada em todas as lutas(22).
É aceito que o aumento da produção de força é devido a fatores neurais, o que sustenta a hipótese de que ocorrem adaptações na atividade elétrica muscular com o esforço físico. Na literatura essas alterações em estado de fadiga aparecem de formas divergentes. Por outro lado, já está bem estabelecida a relação entre o aumento da atividade de enzimas musculares plasmáticas com o aumento dos esforços causados pelos exercícios ou com os estados de lesões musculares.
O estresse de diferentes durações de luta de judô poderia ser suficiente para causar alterações enzimáticas indicativas de aumento dos esforços causados pelos exercícios e/ou estados de lesões musculares. Esses mesmos esforços poderiam alterar a atividade eletromiográfica dos músculos analisados, alteração esta caracterizada por uma co-ativação muscular, que talvez comprometesse a produção de força dos membros superiores dos atletas.
O aumento dos níveis enzimáticos no plasma poderia alterar a atividade elétrica do músculo, e talvez estes parâmetros bioquímicos teriam correlação com os sinais elétricos indicativos de fadiga muscular. O que poderia caracterizar um estado deficiente de preparação física dos atletas para a competição, por falta ou excesso de treinamento.
O objetivo deste estudo foi verificar se as diferentes durações de lutas de judô, respectivamente de 90s, 180s e 300s, poderiam alterar o metabolismo enzimático e a atividade eletromiográfica dos atletas, prejudicando a manutenção do torque, indicativa de fadiga muscular.
MÉTODOS
Sujeitos
Doze atletas de elite de judô do sexo masculino, com idade de 22 ± 4,5 anos, assinaram voluntariamente o termo de consentimento da pesquisa. Este estudo foi aprovado pelo Comitê de Ética em Pesquisa da UNIVAP sob o protocolo nº A017/2003/CEP.
Protocolo de lutas
Os sujeitos foram divididos em duplas de acordo com o peso corporal (kg), mensurado com uma balança Filizola. Três lutas com as durações de 90s, 180s e 300s foram realizadas com os pares, nos respectivos horários: 8h, 13h e 18h, em dias alternados com intervalo de 72 horas. Os atletas foram incentivados oralmente pelo técnico da equipe a lutar com a máxima intensidade. Os sujeitos foram avaliados, imediatamente antes e após as lutas.
Protocolo do EMG
Foi utilizado um eletromiógrafo da marca EMG System do Brasil com oito canais, e coletados os sinais EMG dos músculos: deltóide anterior, peitoral maior e redondo maior, respectivamente: agonista, antagonista e sinergista do movimento. Realizou-se uma tricotomia e assepsia local na pele com algodão, álcool e gel. Um eletrodo terra foi posicionado no braço não dominante, e eletrodos bipolares de superfície fixados na porção média dos músculos. O intervalo de tomada dos sinais foi de 30s, a partir do comando de execução do movimento.
Protocolo do dinamômetro isocinético
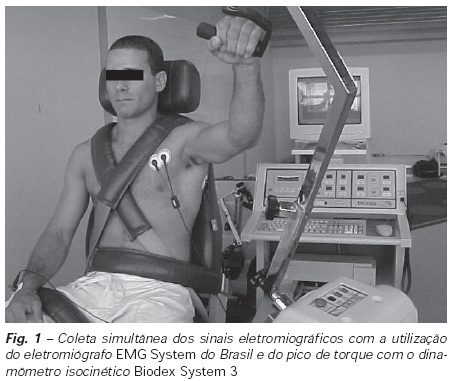
Para mensuração do pico de torque utilizou-se um dinamômetro isocinético Biodex System 3. Os sujeitos permaneceram sentados na cadeira com o eixo de rotação alinhado à região glenoumeral. Os sujeitos realizaram cinco movimentos de flexão e extensão de ombro numa velocidade de 90o/s, otimizando a captação do sinal eletromiográfico em protocolos dinâmicos(18). O movimento iniciou-se com o braço dominante estendido anterior ao corpo num ângulo de 90o a partir da posição fundamental anatômica seguido de uma flexão de ombro abduzido até atingir um ângulo de 70o. Anterior ao experimento, uma adaptação ao equipamento foi realizada durante três dias. A ação motora foi baseada no movimento da puxada de manga do judô, realizado com freqüência pelo atleta. Para sincronização dos equipamentos foi utilizada uma chave acoplada ao braço do dinamômetro, como um dispositivo de tempo de início da execução do movimento, permitindo a coleta simultânea dos sinais eletromiográficos e do dinamômetro (figura 1).
Protocolo do processamento dos sinais
Os sinais foram processados com a utilização do MATLAB Math Works 6.1. Os dados brutos foram coletados num intervalo de 15s, a 2KHz de freqüência de amostragem, filtrados por um passa alta de 20Hz e um passa baixa de 250Hz e retificados pela FFT (Fast Fourier Transformation). Para obter a mesma resolução temporal dos sinais EMG (0,05ms) e do torque (10ms), o sinal do torque foi interpolado com a utilização do spline cúbico em intervalos de 0,05ms. Foi analisada a 1ª fase de cada contração concêntrica a partir do início do movimento. Os sinais EMG foram normalizados pela linha de base e analisados no domínio do tempo, através do cálculo da integral do valor absoluto do sinal EMG (iEMG) de cada contração, com a utilização da seguinte equação:

onde T = tempo de duração da contração.
Protocolo da coleta de amostra sanguínea e dosagem bioquímica
Foram coletados 10ml de sangue dos sujeitos, com a utilização de seringa e agulha descartáveis. As amostras foram imediatamente centrifugadas, com a rotação de 10.000RPM durante 10min, com a utilização da centrífuga JOUAN CR3I, refrigerada a 4oC. Verificou-se a atividade das enzimas LDH, CK, AST e ALT, com a utilização do espectofotômetro HITACHI UV-2001 e do KIT Analisa Diagnóstica, pelo método colorimétrico.
Análise estatística dos dados
Para análise estatística dos dados utilizou-se o teste não paramétrico Wilcoxon para dados pareados, com um nível de significância de a = 0,05.
RESULTADOS
Nenhuma diferença foi encontrada no pico de torque (PT) antes e após as diferentes durações de luta, como podemos verificar nos valores expressos na tabela 1.
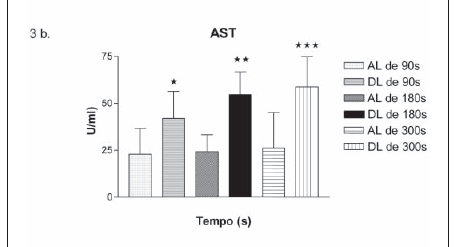
Na atividade da AST e da ALT verificou-se um aumento, relacionado ao aumento da duração das lutas de 90s, 180s e 300s, como observado nos valores da tabela 2 e figura 3.

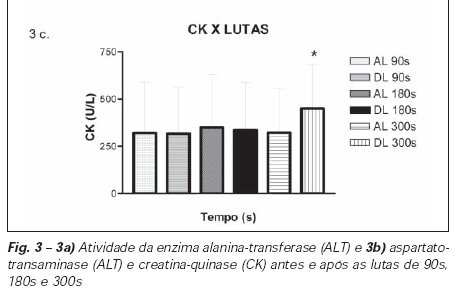
Observou-se, na atividade da LDH, uma diminuição apenas após a luta de 90s como verificada na tabela 2 e, na atividade da CK, um aumento após a luta de 300s, conforme a figura 3c.
Na luta de 90s observou-se um aumento no sinal iEMG (mV) após todos os músculos. Porém esse aumento só foi significativo (p = 0,005) no músculo agonista, como observado na tabela 3.
Após a luta de 180s, o músculo agonista não apresentou alterações, mantendo os valores de repouso; observou-se um aumento (p = 0,0129) do sinal iEMG (mV) no músculo antagonista e um aumento, apesar de não significativo, no sinergista.
Com a luta de 300s verificou-se uma diminuição da atividade do músculo agonista (p = 0,0137) e uma diminuição (p = 0,05), apesar de não significativa, na ativação do antagonista, com o músculo sinergista mantendo seus valores médios de repouso.
Podemos observar uma diminuição na intensidade da atividade elétrica dos músculos analisados a partir da duração das lutas, como observado na figura 4.

DISCUSSÃO
Os valores médios da tabela 1 sugerem que o estresse das lutas ao qual os sujeitos foram submetidos não foi suficiente para alterar o pico de torque dos músculos e/ou o tempo de descanso entre o esforço e a tomada dos sinais foi suficientemente recuperador para a capacidade de gerar força nos músculos analisados.
Os principais fenômenos biomecânicos foram verificados com a análise simultânea dos músculos envolvidos no movimento, já observado por outros autores(23).
Apesar de a luta de 90s não ter sido suficiente para diminuir o torque, provocou adaptações neurais observadas no aumento do sinal iEMG do agonista, que pode representar uma dinâmica de adaptação verificada em outros estudos com protocolos de fadiga(18).
Ao contrário de Kellis(17), pôde ser verificada na luta de 180s um aumento na amplitude do sinal iEMG do antagonista, sugerindo que este músculo desempenhou importante papel na estabilidade da articulação, como controle motor de regulação e proteção das articulações em processo de fadiga(24). Uma explicação para esse aumento pode ser atribuída a uma tentativa do sistema nervoso central de compensar a fadiga muscular através do aumento do número de unidades motoras disponíveis(25).
Se o músculo antagonista é capaz de aumentar a força durante esforços repetidos, ele poderia representar uma carga adicional para atender o músculo agonista e manter o esforço, contribuindo no momento para a diminuição do resultante na articulação envolvida, protegendo-a, devendo ser, portanto, um parâmetro considerado, quando se objetiva a performance no treinamento de gestos esportivos.
Por outro lado, essa co-ativação sugere uma debilidade na performance de ativação dos músculos para a realização da ação motora, o que poderia significar um prejuízo para a performance funcional(24). A otimização do treino de força visando performance de atletas objetiva ativações agonistas musculares sem ou com baixa co-ativação antagonista(16,22).
As adaptações neuromusculares relacionadas ao tempo de duração das lutas sugerem que se necessitou de uma demanda energética maior para realização do mesmo esforço, observada com a atividade da AST e ALT que foi regulada pela demanda energética necessária aos esforços. Mesmo com o predomínio da via glicolítica, nas lutas de judô, as proteínas musculares, alanina e glutamina, foram recrutadas para a síntese de ATP, sugerindo que a AST e ALT, neste estudo, foram biomarcadoras do nível de intensidade do exercício(6).
O aumento do iEMG (mV) do agonista após a 1ª luta e a co-ativação após a 2ª luta podem ser explicados por uma dinâmica de adaptação neuromuscular para realização do trabalho, que acompanha um aumento gradativo de demanda energética caracterizada pelas atividades da AST e ALT, conforme figuras 3a e 3b.
Na análise da LDH, verificou-se uma diminuição em todos os protocolos de lutas, porém só foi significativa após a luta de 90s, que pode ser explicada pela capacidade potencializada de resistência aeróbia, em virtude do treinamento a que os atletas são submetidos. É aceito que o treino de resistência e reduzida intensidade provoca uma acentuada redução na atividade da LDH(26,27). Dessa forma, o fator que regulará o aumento da LDH é a oferta de piruvato e NADH. Talvez a condição de treinamento dos sujeitos fez com que os níveis desses substratos não excedessem a capacidade do PDH em metabolizar piruvato e/ou do sistema alfa-glicerofosfato suprir as demandas energéticas(28). Uma vez que a LDH é uma enzima reguladora, talvez um modulador negativo, que poderia ser a redução de NADH citossólico, provocaria essa inibição.
Nas lutas de 180s e 300s, o aumento potencializado do lactato em virtude da característica intermitente da luta de judô talvez tenha sido suficiente para não permitir uma diminuição significativa da atividade da LDH(29).
Nas lutas de 300s o sinal iEMG (mV) apresentou uma diminuição da atividade agonista e antagonista, ao mesmo tempo em que manteve a ativação sinergista como nos valores de repouso. A ativação diminuída do agonista pode sugerir uma deficiência na ação motora, combinada com a atuação do sinergista, que deveria estar estabilizando o movimento, e não agindo como motor primário.
Esses dados mostram a importância dos estudos na atividade sinergista do movimento, de forma que para um treinamento de força efetivo, além da ativação do agonista e redução da co-ativação, uma otimização do sinergista poderia exercer um importante papel no aumento do sistema de torque produzido sobre as articulações(12).
Verificou-se que o protocolo da luta de 300s provocou adaptações fisiológicas com o aumento da CK, que prejudicou a ativação das unidades motoras e/ou a propagação do sinal elétrico muscular.
A não alteração na atividade da CK nas duas primeiras lutas pode ser explicada pelas variações interindividuais expressas pelos valores elevados de desvio-padrão, dificultando a análise das alterações causadas pelo exercício.
O aumento da CK após as lutas de 300s pode ser resultado do esforço submáximo intermitente(6) a que os sujeitos foram submetidos, com aumento de temperatura e taxa metabólica e predomínio de contração excêntrica, aumentando liberação desta enzima(30). A intensidade do exercício e sua duração são variáveis que estão diretamente relacionadas à alteração das concentrações plasmáticas de CK(30). À medida que o esforço aumenta, a porosidade do sarcolema e/ou o rompimento da membrana provoca a passagem destas proteínas celulares para o plasma, fenômeno observado neste estudo, após a luta de 300s.
A diminuição do iEMG do músculo motor primário do movimento, associada ao aumento da CK plasmática durante a luta de 300s, sugere que o esforço foi suficiente para causar um aumento na permeabilidade e/ou microlesões no sarcolema que provocou uma alteração na polaridade, prejudicando a ativação e propagação dos sinais elétricos, resultando na diminuição da amplitude do sinal eletromiográfico.
Esse fenômeno pode ser resultado da presença de subprodutos metabólicos decorrentes do exercício, provocando uma alteração no potencial de membrana, pela diminuição do pH, decorrente da acidificação causada pela lesão na fibra muscular, que pode ter levado a prejuízos na ativação celular(31).
As lutas às quais os atletas foram submetidos neste estudo constituem-se num fator limitante, visto que, apesar de estimuladas pelo treinador e obedecer a todas as normas de uma competição oficial, foram simuladas para a realização do experimento. De forma que os atletas poderiam apresentar respostas fisiológicas diferenciadas e individuais numa competição oficial.
Apesar de verificado aumento de intensidades dos esforços através da atividade das enzimas AST e ALT, a mecânica de ações musculares utilizadas nas diferentes durações de luta é outro aspecto a ser considerado, uma vez que as lutas divergem nas ações, limitando a avaliação funcional das ações motoras usadas pelos atletas durante as lutas.
As adaptações no sinal EMG observadas neste estudo evidenciam a necessidade da análise simultânea da coordenação intermuscular. Os resultados da atividade da AST e ALT relacionada com as respectivas durações de lutas confirmam a utilização destas enzimas como biomarcadoras de intensidade de esforços, indicando uma dinâmica de adaptação metabólica aos esforços que foram gradativamente aumentados.
As alterações nos parâmetros elétricos e bioquímicos no protocolo proposto sugerem que a intensidade do esforço está relacionada com a alteração da atividade elétrica do tecido muscular esquelético.
E que os efeitos da luta de 300s provocaram aumento da atividade da CK e diminuição da atividade elétrica do músculo agonista. Sugerindo que a diminuição da amplitude do sinal no músculo motor primário está associada a possíveis microlesões na membrana muscular que prejudicam a ativação e propagação do sinal eletromiográfico.
CONCLUSÃO
Conclui-se que o estresse das lutas de 90s, 180s e 300s, apesar de não alterar a capacidade de torque, foi estímulo suficiente para provocar alterações na atividade enzimática e elétrica muscular que podem ser prejudiciais à performance dos atletas.
Recebido em 5/4/05. Versão final recebida em 15/9/05. Aceito em 19/9/05.
Todos os autores declararam não haver qualquer potencial conflito de interesses referente a este artigo.
- 1. Mujica I, Padilla S. Muscular characteristics of the training in humans. Med Sci Sports Exerc 2000;33:1297-3.
- 2. Coyle EF. Detraining and retention of training-induced adaptations. Med Sci Sports Exerc 1990;2:1-5.
- 3. Apple FS, Hellsten Y, Clarkson PM. Early detection of skeletal muscle injury by assay creatine kinase MM isoforms in serum after acute exercise. Clin Chem 1998;32:41-4.
- 4. Volfinger L, Lassorourd V, Michaux JM, Braun JP, Tiurtain PL. Kinetic evaluation of muscle damage during exercise by calculation of amount of creatine kinase released. Am J Physiol 1994;266:434-1.
- 5. Meulen Van Der JH, Kuipers H, Drukker J. Relationship between exercise-induced muscle damage and enzyme release in rats. The American Physiological Society 1991;161:999-4.
- 6. Lijnen P, Hespel P, Fagard R, Lysens R, Van Den Eynde E, Goris M, et al. Indicators of cell breakdown in plasma of men during and after a marathon race. J Sports Med 1988;9:108-13.
- 7. Masuda T, Tizuca T, Zhe JY, et al. Influence of contraction force and speed on muscle fiber conduction velocity during dynamic voluntary exercise. J Electromyogr Kinesiol 2001;1:85-4.
- 8. Perry SR, Housh TJ, Weir JP, Johnson GO, Bull A, Ebersole KT. Mean power frequency and amplitude of the mechanomyographic and electromyographic signals during incremental cycle ergometry. J Electromyogr Kinesiol 2001;11:299-5.
- 9. Gabriel DA, Basford JR, Kai-Nan AN. Neural adaptations to fatigue: implications for muscle strength and training. Med Sci Sports Exerc 2001;33:1354-69.
- 10. Hakkinen K, Kramer WJ, Newton RU, Alen M. Changes in electromyographic activity muscle fibre and force production characteristics during heavy resistance/power strength training in middle-aged and older men and women. Scandinavian Physiological Society 2001;171:51-62.
- 11. Kell RT, Bell G, Quinney A. Muscle skeletal fitness, health outcomes and quality of life. Sports Med 2001;12:863-73.
- 12. Ross A, Leveritt M, Riek S. Neural Influences on sprint running. Sports Med 2001;31:409-25.
- 13. Hawley JA, Stepto NK. Adaptations to training in endurance cyclists. Implications for performance. Sports Med 2001;31:511-0.
- 14. Clarys JP, Cabri J. Electromyography and study of sports movements: a review. J Sports Sci 1993;11:379-48.
- 15. Kazumi M, Tadashi M, Tsugutake S, Mitsuharu I, Shigeru K. Changes in surface EMG parameters during static and dynamic fatiguing contractions. J Electromyogr Kinesiol 1999;9:39-6.
- 16. Vollestad NK. Measurement of human muscle fatigue. J Neurosci Methods 1997; 74:219-7.
- 17. Kellis E. The effects of fatigue on the resultant joint moment, agonist and antagonist electromyographic activity at different angles during dynamic knee extension efforts. J Electromyogr Kinesiol 1999;9:191-9.
- 18. Gerdle B, Larsson B, Karlsson S. Criterion validation of surface EMG variables as fatigue indicators using peak torque. J Electromyogr Kinesiol 2000;10:225-2.
- 19. Lamontagne A, Richards CL, Malouin F. Coactivation during gait as an adaptive behavior after stroke. J Electromyogr Kinesiol 2000;10:407-5.
- 20. Hautier CA, Arsac LM, Deghdegh K. Influence of fatigue on EMG/force ratio and co-contraction in cycling. Med Sci Sports Exerc 2000;32:839-43.
- 21. Gerdle B, Wretling ML, Henriksson-Larsén K. Do the fibre-type proportion and angular velocity influence the mean power frequency of the electromyogram? Acta Physiol Scand 1988;134:341-6.
- 22. Franchini E. Características fisiológicas em testes laboratoriais e resposta da concentração de lactato sanguíneo de três lutas em judocas das classes juvenil-A, Júnior e Sênior. Revista Paulista de Educação Física 1998;12:5-16.
- 23. Berger W, Trippl M, Discher M, Dietz V. Influence of subjects height on the stabilization of posture. Acta Otolaryngol 1992;30:112-22.
- 24. Van Handel PJ, Watson P, Troup J, Plyley M. Effects of treadmill running on oxidative capacity of regenerated skeletal muscle. Clin J Sport Med 1981;2:92-6.
- 25. Spriet LL, Heigenhairser GJF. Regulation of Pyruvate Dehydrogenase (PDH) activity in human skeletal muscle during exercise. Exerc Sport Sci Rev 2002;30: 91-5.
- 26. Almada A, Mitchell T, Earnest C. Impact of chronic creatine supplementation on serum enzyme concentrations. FASEB J 1996;10:45-67.
- 27. Noakes TD. Effect of exercise on serum activities in humans. Sports Med 1987; 4:245-67.
- 28. Bourdon L, Stieglitz P, Pouzeratte N, Curé M. Effect of incubation temperature on the creatine kinase release from in vitro rat skeletal muscle preparation. J Therm Biol 1996;21:109-13.
- 29. Hortobagyi T, Denahan D. Variability in creatine kinase: methodological, exercise and clinically related factors. Int J Sports Med 1989;10:69-80.
- 30. Janssen GM, Kuipers H, Willems GM, Does RJ, Janssen, EP, Geurten P. Plasma activity of muscle enzymes. Quantification of skeletal muscle damage and relationship with metabolic variables. J Sports Med 1989;3:160-8.
- 31. Dimitrova NA, Dimitrov GH. Interpretation of EMG changes with fatigue: facts, pitfalls, and fallacies. J Electromyogr Kinesiol 2003;13:13-26.
Endereço para correspondência:
Datas de Publicação
-
Publicação nesta coleção
02 Maio 2007 -
Data do Fascículo
Fev 2006
Histórico
-
Aceito
19 Set 2005 -
Revisado
15 Set 2005 -
Recebido
05 Abr 2005