Resumos
As glândulas mandibulares de operárias recém-emergidas, nutridoras e campeiras, rainhas virgens e fisogástricas, e machos maduros de Scaptotrigona postica Latreille foram estudadas histológica e morfometricamente. Estas glândulas apresentam diferenças histológicas características para cada idade e/ou função do indivíduo. De acordo com a morfometria conclui-se que as glândulas mandibulares possuem um ciclo secretor determinado, no qual a secreção começa a ser produzida nas operárias recém-emergidas, e acumula-se no citoplasma e reservatório da glândula das operárias nutridoras. Nas operárias campeiras o citoplasma encontra-se bastante vacuolizado, o que foi interpretado como resultado da eliminação da secreção. Rainhas fisogástricas apresentaram as maiores áreas celulares e nucleares, ao oposto dos machos, os quais apresentaram as menores áreas.
Anatomia; histologia; casta; sexo; estado funcional
The mandibular glands of newly-emerged, nurse and forager workers, virgin and physogastric queens, and mature males of Scaptotrigona postica Latreille were studied under histological and morphometrical analyses. These glands present differences in their histology, according to the age and/or function of the individuals. The morphometric results showed that the mandibular glands have a very markedly secretory cicle, in which the secretion starts to be produced in newly emerged workers, being accumulated into the cytoplasm and reservoir in the nurses. In forager workers the cytoplasm presents many vacuoles, which are remaining of the secretion releasing. Physogastric queens showed the largest cellular and nuclear areas. On the contrary, the males presented the smallest ones.
Mandibular gland; anatomy; histology; caste; sex; functional state
SYSTEMATICS, MORPHOLOGY AND PHYSIOLOGY
Caracterização das glândulas mandibulares nas diferentes classes de adultos de Scaptotrigona postica Latreille (Hymenoptera: Apidae)
Characterization of the mandibular glands in different adult types of Scaptotrigona postica Latreille (Hymenoptera: Apidae)
Luciana F. Gracioli-Vitti; Fábio C. Abdalla; Carminda da Cruz-Landim
Depto. Biologia, Instituto de Biociências, UNESP, Campus de Rio Claro. Av. 24A, 1515. 13506-900, Rio Claro, SP
RESUMO
As glândulas mandibulares de operárias recém-emergidas, nutridoras e campeiras, rainhas virgens e fisogástricas, e machos maduros de Scaptotrigona postica Latreille foram estudadas histológica e morfometricamente. Estas glândulas apresentam diferenças histológicas características para cada idade e/ou função do indivíduo. De acordo com a morfometria conclui-se que as glândulas mandibulares possuem um ciclo secretor determinado, no qual a secreção começa a ser produzida nas operárias recém-emergidas, e acumula-se no citoplasma e reservatório da glândula das operárias nutridoras. Nas operárias campeiras o citoplasma encontra-se bastante vacuolizado, o que foi interpretado como resultado da eliminação da secreção. Rainhas fisogástricas apresentaram as maiores áreas celulares e nucleares, ao oposto dos machos, os quais apresentaram as menores áreas.
Palavras-chave: Anatomia, histologia, casta, sexo, estado funcional
ABSTRACT
The mandibular glands of newly-emerged, nurse and forager workers, virgin and physogastric queens, and mature males of Scaptotrigona postica Latreille were studied under histological and morphometrical analyses. These glands present differences in their histology, according to the age and/or function of the individuals. The morphometric results showed that the mandibular glands have a very markedly secretory cicle, in which the secretion starts to be produced in newly emerged workers, being accumulated into the cytoplasm and reservoir in the nurses. In forager workers the cytoplasm presents many vacuoles, which are remaining of the secretion releasing. Physogastric queens showed the largest cellular and nuclear areas. On the contrary, the males presented the smallest ones.
Key words: Mandibular gland, anatomy, histology, caste, sex, functional state
Numa colônia de abelhas eussociais, há uma divisão do trabalho reprodutivo entre as castas femininas, sendo a rainha responsável pela postura e ficando para as operárias todas as outras tarefas da colônia, entre elas a de nutrir a cria. A divisão de trabalho em abelhas está baseada na capacitação do indivíduo para exercer determinada tarefa. No caso das operárias, a capacitação é alcançada através da maturação fisiológica para seu desempenho, adquirida por etapas, com o avanço da idade, o polietismo etário, no qual as estruturas glandulares endócrinas e exócrinas desempenham importante papel.
A idade das operárias em que essas glândulas se desenvolvem ou degeneram não está rigidamente estabelecida no programa inato do indivíduo e, aparentemente, nem sequer a seqüência a seguir. Assim as operárias podem responder às necessidades da colônia em um dado momento, ou seja, são as variações das condições do seu meio com ajustes que levam a colônia a funcionar como um superorganismo. Assim, o redirecionamento fisiológico dos tecidos e órgãos acontece (Cruz-Landim & Silva de Moraes 2000)
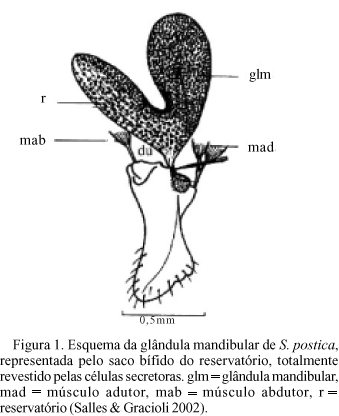
As glândulas de especial interesse neste trabalho são as glândulas mandibulares, as quais são compostas por células da classe III, segundo Noirot & Quennedey (1974, 1991). Essas glândulas, em Scaptotrigona postica Latreille, estão presentes tanto em rainhas como em operárias e machos e constituem, juntamente com as hipofaríngeas e as salivares, o sistema salivar, da mesma forma que em Apis (Cruz-Landim 1967). São estruturas pares, localizadas na cabeça das abelhas, em íntima relação com as mandíbulas e constituídas por um conjunto de células secretoras, que revestem o reservatório, o qual apresenta forma bífida. As glândulas mandibulares de rainhas de meliponíneos, de maneira geral, são menores que as das operárias (Cruz-Landim 1967).
É sabido que a glândula mandibular apresenta diferentes função nas diferentes espécies (Kerr & Cruz 1961). Em S. postica, as operárias usam a secreção para comunicar a localização de fontes de alimentos. Essas abelhas marcam o caminho da fonte de alimento até a colônia com gotículas de secreção depositadas de espaços em espaços (Lindauer & Kerr 1960). As trilhas não são colônia-específicas, ao contrário, abelhas de uma colônia podem ser atraídas por marcas deixadas por abelhas de outra colônia (Lindauer & Kerr 1960).
Além das diferenças entre as espécies, as glândulas mandibulares apresentam, também, diferenças dentro da mesma espécie, seja por meio dos produtos secretados pelas diferentes castas ou sexos, ou seja dentro da mesma casta, no decorrer da idade e/ou função por ela desempenhada. Por exemplo, em Melipona bicolor Lepeletier, diferenças etárias casta-específicas foram evidenciadas por variações histológicas e celulares entre operárias, rainhas virgens e fisogástricas (Gracioli & Silva de Moraes 2002), bem como entre os produtos secretados por elas (Gracioli et al. no prelo).
O presente trabalho tem como objetivo verificar divergências relacionadas à histologia das glândulas mandibulares de operárias (recém-emergidas, nutridoras e campeiras), rainhas (virgens e fisogástricas) e machos maduros de S. postica, bem como alterações nas áreas celulares e nucleares das células secretoras nestas classes de indivíduos.
Material e Métodos
Obtenção de Indivíduos. Para o presente experimento foram utilizadas operárias (recém-emergidas, nutridoras e campeiras), rainhas (virgens e fisogástricas) e machos maduros de S. postica, provenientes de colônias mantidas em laboratório no Instituto de Biociências de Rio Claro da UNESP - Rio Claro. Foram tomadas como operárias nutridoras aquelas trabalhando no aprovisionamento dos alvéolos de cria e como campeiras as que retornavam do forrageamento. Os machos maduros são aqueles capturados fora da colônia, nos enxames de machos.
Histologia. As glândulas mandibulares de operárias (recém-emergidas, nutridoras e campeiras), rainhas (virgens e fisogástricas) e machos maduros de S. postica foram dissecadas em salina para insetos (NaCl - 3,75g, KH2PO4 - 1,76 g, Na2HPO4 - 1,98 g para 500 ml de água destilada) e fixadas, durante 24h, em paraformoldeído 4%. Posteriormente, sofreram desidratação em uma série de alcoóis de concentração crescente e foram incluídas em resina JB - 4 (Polysciences). Os cortes, com espessura de 7mm, foram realizados em histomicrótomo BIO-RAD JB-4, recolhidos em lâminas e corados com Azul de Toluidina, durante 5 min. sendo, posteriormente, lavados em água corrente. Os cortes foram montados com Entellan e lamínula, examinados e registrados em fotomicroscópio (Leica).
Análise Morfométrica e Tratamento Estatístico dos Dados. As células secretoras e os respectivos núcleos das glândulas mandibulares dos indivíduos foram medidos com a utilização de um microscópio Leica, acoplado a um computador, provido com o programa Qwin, que permite a medição e o cálculo de áreas diretamente de cortes histológicos. Foram estudadas 10 glândulas para cada grupo (operárias recém-emergidas, nutridoras e campeiras, rainhas virgens e fisogástricas e machos maduros), com cinco medidas para cada um, obtendo-se, portanto, n = 50.
As médias e desvios padrões foram calculados e os resultados estatisticamente analisados através de análise de variância e pelo teste de Tukey, a 5% de significância (Zar 1996). A análise de variância foi utilizada para verificar diferenças entre os grupos e o teste de Tukey, indicado para amostras com tamanhos (n) iguais, para a localização das diferenças.
Resultados
Histologia. Através das dissecções pode-se observar que as glândulas mandibulares de S. postica encontram-se localizadas lateralmente, na cabeça das abelhas, com a desembocadura ligada à face interna da base da mandíbula. São, em todas as castas e sexos, formadas por um reservatório em forma de saco bífido, parcialmente revestido por células secretoras (Fig. 1).
As células secretoras têm formato de cilíndrico (Figs. 2A, B, D; 3B ) a esférico (Figs. 2C; 3A ) nas castas, entretanto, nos machos essas células, assim como seus núcleos, possuem formato irregular (Figs. 3C, D). Essas células ligam-se ao reservatório, de parede muito fina (Figs. 2D; 3A ), através de canalículos (Figs. 2B, C; 3A ). As células que delimitam o reservatório, assim como os núcleos, são achatadas e pequenas e recobertas por cutícula na face luminal (Figs. 2D; 3A ). Secreção na luz do reservatório é observada em operárias nutridoras (Fig. 2C) e em machos (Figs. 3C, D).
Comparação Entre Castas e Sexos. Células secretoras bastante íntegras de núcleos centralizados e com cromatina bastante dispersa são observadas em operárias recém-emergidas e apresentam região de canalículos coletores bastante corados e definidos (Figs. 2A, B). Nas operárias nutridoras, os núcleos das células secretoras, centralizados, apresentam cromatina bastante compactada (Fig. 2C). Há muita secreção na luz do reservatório (Fig. 2C). Nas operárias campeiras, as células secretoras das glândulas mandibulares possuem citoplasma vacuolizado, onde os núcleos têm cromatina dispersa e a região dos canalículos coletores não está muito evidenciada (Fig. 2D).
As células secretoras das glândulas mandibulares de rainhas virgens são bastante semelhantes às de operárias recém-emergidas, com citoplasma bastante íntegro, núcleos com cromatina dispersa e região de canalículos coletores bastante corados (Fig. 3A ). Entretanto, nas rainhas fisogástricas, as células secretoras adquirem citoplasma vacuolizado, os núcleos continuam apresentando cromatina dispersa e a região dos canalículos coletores apresenta-se bastante corada (Fig. 3B ).
As células secretoras das glândulas mandibulares dos machos possuem formato diferenciado das encontradas nas fêmeas, apresentando limites celulares e núcleos irregulares. A presença de secreção é observada na luz do reservatório dessas glândulas (Figs. 3C, D).
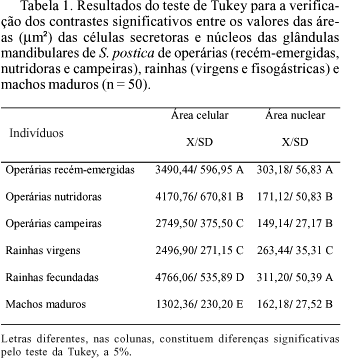
Análise Morfométrica e Tratamento Estatístico dos Dados. A análise de variância mostra diferenças significativas na área média das células secretoras das glândulas mandibulares entre castas e sexos (P > 0,05). De acordo com o teste de Tukey para localização dos contrastes significativos, a área média das células secretoras das glândulas de rainhas fisogástricas é significativamente maior do que a área celular das rainhas virgens, operárias e machos. Entre as operárias, as nutridoras foram as que apresentaram as maiores áreas celulares, seguidas pelas operárias recém-emergidas, a qual é significativamente maior que a área média de operárias campeiras. Não houve diferença significativa entre células secretoras das glândulas mandibulares de operárias campeiras e rainhas virgens. As menores áreas celulares foram encontradas nas glândulas mandibulares de machos de S. postica (Tabela 1).
Ao se aplicar a análise de variância às áreas nucleares das células secretoras das glândulas mandibulares de S. postica, observam-se também diferenças significativas, verificando-se, através do teste de Tukey, que as áreas dos núcleos das células secretoras das glândulas mandibulares de rainhas fisogástricas são significativamente maiores em relação às das rainhas virgens, operárias nutridoras e campeiras e machos, e não possuem diferenças em relação às operárias recém-emergidas (Tabela 1). A área média dos núcleos das células secretoras das glândulas mandibulares de rainhas virgens é significativamente maior que em relação às operárias nutridoras, campeiras e machos. Estes últimos, por sua vez, não possuem diferença significativa entre si (Tabela 1) e representam os menores núcleos analisados.
Discussão
Em todas as glândulas estudadas, o citoplasma apresentou-se basófilo, provavelmente devido à secreção. Portanto, embora a qualidade da secreção não tenha sido estudada no presente trabalho, observa-se, por meio das análises histológicas, que a coloração por hematoxilina-eosina não diferenciou as castas e sexos e/ou fases estudadas, embora sua concentração variada possa ser considerada um indicativo de diferentes períodos de atividade dessas glândulas. Esse fato é registrado em Apis, onde operárias jovens desempenhando tarefas dentro da colônia produzem, principalmente, o ácido 10-hidroxidecenóico em suas glândulas mandibulares, enquanto as mais velhas, trabalhando como forrageiras, produzem, sobretudo, 2-heptanona (Callow et al. 1959, Butler 1966, Crewe 1976).
O significado da presença de vacúolos citoplasmáticos observados em células secretoras das glândulas mandibulares de operárias campeiras e rainhas fisogástricas ainda é desconhecido, podendo apenas ser aventada sua relação com a desestruturação do citoplasma. Os vacúolos poderiam também ser resultado da eliminação de secreção, visto que operárias campeiras utilizam a secreção dessas glândulas para marcação de trilha de cheiro (Lindauer & Kerr 1960) e, rainhas fisogástricas talvez a utilizem para manutenção e controle da colônia, assim como em Apis, onde a rainha exerce grande influência na fisiologia e no comportamento das operárias, principalmente pela utilização da "substância de rainha", sintetizada e liberada pelas glândulas mandibulares (Butler 1957). Contudo, nos meliponíneos a dominância da rainha parece ser exercida por agressividade ritualizada (Zucchi 1993). Os vacúolos podem, também, corresponder a acúmulos lipídicos, visto que Cruz-Landim & Puga (1967) verificaram a presença de lipídeos no citoplasma das células secretoras, bem como na luz do reservatório das glândulas mandibulares de S. postica.
A secreção que aparece acumulada em torno da região da porção do canalículo coletor, representada por uma coloração forte, pode estar no citoplasma ou, à semelhança do que ocorre nas glândulas hipofaríngeas de Apis (Knecht & Kaatz 1990), no espaço entre a membrana plasmática da célula secretora envolvente e a parede do canalículo. Esse espaço, denominado espaço pericanalicular, funcionaria, então, como reservatório celular para a secreção antes de sua eliminação, o que resulta em aumento no volume citoplasmático. Secreção estocada no espaço pericanalicular também foi observada em glândulas mandibulares de M. bicolor (Gracioli & Silva de Moraes 2002).
As alterações das áreas celulares das glândulas mandibulares detectadas pelas análises morfométricas dependem de mudanças citoplasmáticas provocadas por variações no número e grau de desenvolvimento de organelas e, sobretudo, pelo acúmulo de secreção, assim como as alterações nucleares refletem seu nível de atividade (Gracioli & Silva de Moraes 2002). Esses aspectos celulares são considerados bons parâmetros para avaliação da atividade glandular por serem proeminentes e facilmente detectáveis (Knecht & Kaatz 1990). Portanto, tomando-se as variações das áreas celulares das glândulas de S. postica indicativas de variações na intensidade dos processos de síntese, os resultados morfológicos indicaram que as células secretoras de rainhas fisogástricas têm nível mais alto de atividade, refletido pelas maiores áreas celulares, seguidos pelas operárias nutridoras e, posteriormente, pelas recém-emergidas, campeiras e machos.
Cruz-Landim & Ferreira (1968) verificaram que essas glândulas, em operárias de S. postica, acumulam secreção em seus reservatórios até a meia idade, tornando-se vazias em operárias velhas e, embora tenham funções distintas daquelas de Apis, o mecanismo de funcionamento dessas glândulas em ambas as espécies deve ser o mesmo, com base nos dados morfológicos. Francke et al. (1983) analisaram extratos da cabeça de operárias de S. postica, em diferentes idades, onde observaram a presença de bouquets específicos típicos para cada idade.
Assim, as células secretoras das glândulas mandibulares de S. postica aumentam de tamanho até a função de nutridoras, regredindo, posteriormente, nas campeiras. Do mesmo modo, rainhas virgens possuem glândulas mandibulares menores que as encontradas em rainhas fisogástricas, as quais utilizam a secreção na comunicação com as operárias dentro da colônia (Velthuis 1997).
Da mesma forma, as alterações encontradas nos núcleos refletem alta atividade destes em operárias recém-emergidas, as quais apresentaram núcleos maiores, o que indica, em sua próxima etapa da vida (função de nutridora), alta atividade celular e, conseqüentemente, maiores áreas celulares. Rainhas fisogástricas também apresentaram núcleos em alta atividade, o que se interpreta como concomitante síntese celular. Assim, as variações no tamanho dos núcleos devem refletir variações na atividade de síntese, e as variações no tamanho da célula devem refletir o acúmulo de material nestas. A variabilidade das duas partes representa momentos diferentes de atividade celular.
Em rainhas virgens de S. postica, o produto secretado pelas glândulas mandibulares serve de comunicação com os machos durante o vôo nupcial, enquanto, mais tarde, as glândulas têm função de comunicação com as operárias. Isto é resultado da rápida mudança de sua composição, como conseqüência do acasalamento (Velthuis 1997).
Nos machos, como não foi evidenciada secreção na região dos canalículos, pode-se aventar que a mesma é liberada no reservatório concomitantemente ao processo de síntese, permanecendo, então, estocada na luz do reservatório. Esta não volta a ser sintetizada, pois os machos a utilizam quando estão maduros e saem para fazer o vôo do acasalamento, liberando essa secreção como feromônio sexual. Ao liberarem os espermatozóides, durante a fecundação da rainha, eles perdem a genitália externa e, às vezes, parte do intestino, morrendo logo após o ato. Em vista disto, após o uso da secreção das glândulas mandibulares, o destino desses indivíduos é a morte, caso eles consigam fecundar alguma rainha; assim, não há necessidade de continuidade de síntese da secreção, o que justifica a ausência de secreção na região dos canalículos.
A ocorrência dos altos desvios padrões detectados nos grupos de indivíduos estudados pode ser atribuída à plasticidade morfo-funcional das glândulas em geral e, em especial da mandibular, que se mostra reduzida ou hipertrofiada ao longo do desenvolvimento, de acordo com as necessidades ambientais e com a idade e/ou atividade do indivíduo dentro da colônia (Cruz-Landim 1994).
Os resultados histológicos e morfométricos evidenciam que as operárias recém-emergidas possuem células secretoras das glândulas mandibulares mais íntegras quando comparadas às outras operárias, e alta atividade nuclear, o que sugere futura atividade de síntese celular. As operárias nutridoras parecem estar em alta atividade secretora e acumulam secreção no reservatório, enquanto as campeiras, que possuem células em estado de desintegração, com grandes e numerosos vacúolos, parecem refletir certo envelhecimento celular (Cruz-Landim & Silva de Moraes 2000). As rainhas virgens possuem células bastante íntegras e alta atividade nuclear, pois, provavelmente, estão se preparando para o vôo do acasalamento, onde, talvez, utilizarão a secreção aí produzida para a comunicação com os machos; já as rainhas fisogástricas, embora apresentem citoplasma vacuolizado, possuem área celular e nuclear em alta atividade, pois, eventualmente, estão utilizando a secreção glandular para a comunicação com as operárias dentro da colônia. As glândulas mandibulares dos machos representam as menores áreas celulares e nucleares, o que sugere que utilizem a secreção destas glândulas exclusivamente como feromônio sexual durante o vôo de acasalamento.
Agradecimentos
À Fundação de Amparo a Pesquisa do Estado de São Paulo (Proc.02/08625-5).
Literatura Citada
Received 10/08/04. Accepted 15/08/04.
- Butler, C.G. 1957. The process of queen supersedure in colonies of honeybees (Apis mellifera). Insects Sociaux 4: 211-223.
- Butler, C.G. 1966. Mandibular gland pheromone of worker honeybee. Nature 212: 530.
- Callow, R.K., N.C. Johnston & J. Simpson. 1959. 10-hydroxy 2-decenoic acid in the honeybee (Apis mellifera). Experientia 15: 421.
- Crewe, R.M. 1976. Agressiveness of honeybees and their pheromone production. South African. J. Sci. 72: 209-212.
- Cruz-Landim, C. 1967. Estudo comparativo de algumas glândulas das abelhas (Hymenoptera, Apoidea) e respectivas implicações evolutivas. Arq. Zool. São Paulo 15: 177-290.
- Cruz-Landim, C. 1994. Polimorfismo na ocorrência das glândulas exócrinas nas abelhas (Hymenoptera, Apoidea). In Encontro sobre abelhas, 1, Ribeirão Preto. Anais. Ribeirão Preto (SP). 118-129p.
- Cruz-Landim, C. & A. Ferreira. 1968. Mandibular gland development and communication in field bees of Trigona (Scaptotrigona) postica (Hymenoptera: Apidae). J. Kansas Entomol. Soc. 41: 474-481.
- Cruz-Landim, C. & F.R. Puga. 1967. Presença de substâncias lipídicas nas glândulas do sistema salivar de Trigona (Hym., Apoidea). Papéis Av. Zool. 20: 65-74.
- Cruz-Landim, C. & R.L.M. Silva de Moraes. 2000. Morte celular programada em abelhas como uma forma de redirecionar a morfologia e a fisiologia adaptativa. Editora e Tipografia Costa. Rio Claro, SP, 48p.
- Francke, W., W. Schroder, E. Engels & W. Engels. 1983. Variation in cephalic volatile substances in relation to worker age and behavior in the stingless bee, Scaptotrigona postica Z. Naturforsch. 38c: 1066-1068.
- Gracioli, L.F. & R.L.M. Silva de Moraes. 2002. Histological and morphometric comparisons of worker and queen mandibular glands of Melipona bicolor bicolor (Hymenoptera, Meliponini). Sociobiology 40: 449-456.
- Gracioli, L.F., R.L.M. Silva de Moraes & C. Cruz-Landim. No prelo. Ultrastructural aspects of the mandibular gland of Melipona bicolor Lepeletier, 1836 (Hymenoptera: Apidae, Meliponini) in the castes. Micron. 35: 331-336.
- Kerr, W.E. & C.C. Cruz. 1961. Funções diferentes tomadas pela glândula mandibular na evolução das abelhas em geral e em "trigona (oxytrigona) tataira" em especial. Rev. Bras. Biol. 21: 1-16.
- Knecht, D. & H.H. Kaatz. 1990. Patterns of larval food prodution by hypopharyngeal glands in adult woker honey bees. Apidologie 27: 457- 468.
- Lindauer, M. & W.E. Kerr. 1960. Communication between the workers of stingless bees. Bee World 41: 29-41.
- Noirot, C. & A. Quennedey. 1974. Fine structure of insect epidermal glands. Annu. Rev. Entomol. 19: 61-80.
- Noirot, C. & A. Quennedey. 1991. Glands, gland cells, glandular units: Some comments on terminology and classification. Ann. Soc. Entomol. Fr. (N.S.) 27: 123-128.
- Salles, H.C. & L.F. Gracioli. 2002. Glândulas mandibulares e intramandibulares. 71-90p. In C. Cruz-Landim & F.C. Abdalla (eds.), Glândulas exócrinas das abelhas. FUNPEC, 181p.
- Velthuis, H.H.W. 1997. Biologia das abelhas sem ferrão. Utrecht, Holanda, Universidade de Utrecht, Departamento de Etologia, São Paulo, Universidade de São Paulo, Departamento de Ecologia, 33p.
- Zar, J.H. 1996. Biostatistical analysis, p.226-229. 3° ed. New Jersey, Prentice-Hall.
- Zucchi, R. 1993. Ritualized dominance, evolution of queen-worker interactions and related aspects in stingless bees (Hymenoptera: Apidae), p.207-249 In T. Inoue & S. Yamane (eds.), Evolution of insect societies, Hakuhin-sha, Tokio, 300p.

Datas de Publicação
-
Publicação nesta coleção
18 Mar 2005 -
Data do Fascículo
Dez 2004
Histórico
-
Recebido
10 Ago 2004 -
Aceito
15 Ago 2004