Resumos
Foi testada a preferência para oviposição (com chance de escolha) de Bemisia tabaci (Genn.) biótipo B em 20 genótipos de feijoeiro. Os materiais foram divididos em dois lotes, utilizando-se em cada um deles delineamento em blocos ao acaso, com 20 repetições. Folhas recém-expandidas de cada genótipo foram mantidas em tubos de vidro contendo água destilada, colocadas em uma gaiola acrílica, onde foram liberados cerca de 500 insetos. Após 24h, contou-se o número total de ovos na superfície abaxial e o número de tricomas aciculares e unciformes em duas áreas de 4,9 mm² da superfície abaxial. Mediu-se também a área foliar dos feijoeiros e o comprimento máximo e mínimo dos tricomas, que foram classificados em curto, médio ou longo. Não foram constatadas correlações significativas entre o número de ovos e a densidade de tricomas total e de tricomas aciculares e unciformes. Foram observadas correlações significativas entre o número de ovos e a densidade de tricomas aciculares longos tipo A1 (r = 2,85) e a densidade de tricomas unciformes curtos tipo B3 (r = -2,44). Assim, pode-se inferir que a preferência para oviposição da mosca-branca pela cultivar Bolinha (4,8 ovos/cm²) pode estar relacionada ao grande número de tricomas aciculares longos neste material tipo A1 (31,7 tricomas/4,9 mm²). O contrário pode ser considerado quanto ao genótipo menos preferido, G13028 (1,1 ovos/cm²), que possui principalmente tricomas unciformes curtos tipo B3 (64,1 tricomas/4,9 mm²) em suas folhas.
Insecta; Phaseolus vulgaris; mosca-branca; antixenose
The oviposition preference (free-choice tests) of Bemisia tabaci (Genn.) Biotype B for 20 bean genotypes was tested. The genotypes were divided into two plots and set up in a block-randomized design with 20 replications. New expanded bean leaves were sectioned and placed in glass tubes containing distilled water and disposed in acrylic cages containing approximately 500 insects. After 24h, the total number of eggs laid on the abaxial surface of leaves and the density of straight and hooked trichomes in two areas of 4.9 mm² on the abaxial surface were counted. The leaf area was measured as well as the maximum and minimum length of hooked and straight trichomes, which were classified in short, medium or long. No significant correlation was found between the number of eggs and the density of total trichomes (straight + hooked) or between hooked or straight trichomes. Significant correlation was detected between the number of eggs and long straight trichomes type A1 (r = 2.85) and short hooked trichomes type B3 (r = -2.44). In this way, the ovipositional preference of B. tabaci for Bolinha genotype (4.8 eggs/cm²) in a free-choice test could be associated to the larger number of long straight trichomes type A1 (31.7 trichomes/4.9 mm²) on its leaves surface. In addition, the whitefly ovipositional nonpreference for G13028 wild genotype (1.1 eggs/cm²) could be related with the numerous short hooked trichomes type B3 (64.1 trichomes/4.9 mm²) on its leaves surface.
Insecta; Phaseolus vulgaris; whitefly; antixenosis
CROP PROTECTION
Influência dos tricomas na preferência para oviposição de Bemisia tabaci (Genn.) biótipo B (Hemiptera: Aleyrodidae) em genótipos de feijoeiro
Influence of trichomes on ovipositional preference of Bemisia tabaci (Genn.) biotype B (Hemiptera: Aleyrodidae) for bean genotypes
Maria A. de G. OrianiI; José D. VendramimII; Rogério BrunherottoIII
IDepto. Ciências Biológicas, UNIFEOB, Av. Octávio da Silva Bastos, S/N, 13870-000, São João da Boa Vista SP, mariaoriani@hotmail.com
IIDepto. Entomologia, Fitopatologia e Zoologia Agrícola, ESALQ/USP, C. postal 9, 13418-900, Piracicaba, SP
IIIFESB, Av. Francisco S. Lucchesi Filho, 770, C. postal 183, 12900-000, Bragança Paulista, SP
RESUMO
Foi testada a preferência para oviposição (com chance de escolha) de Bemisia tabaci (Genn.) biótipo B em 20 genótipos de feijoeiro. Os materiais foram divididos em dois lotes, utilizando-se em cada um deles delineamento em blocos ao acaso, com 20 repetições. Folhas recém-expandidas de cada genótipo foram mantidas em tubos de vidro contendo água destilada, colocadas em uma gaiola acrílica, onde foram liberados cerca de 500 insetos. Após 24h, contou-se o número total de ovos na superfície abaxial e o número de tricomas aciculares e unciformes em duas áreas de 4,9 mm2 da superfície abaxial. Mediu-se também a área foliar dos feijoeiros e o comprimento máximo e mínimo dos tricomas, que foram classificados em curto, médio ou longo. Não foram constatadas correlações significativas entre o número de ovos e a densidade de tricomas total e de tricomas aciculares e unciformes. Foram observadas correlações significativas entre o número de ovos e a densidade de tricomas aciculares longos tipo A1 (r = 2,85) e a densidade de tricomas unciformes curtos tipo B3 (r = -2,44). Assim, pode-se inferir que a preferência para oviposição da mosca-branca pela cultivar Bolinha (4,8 ovos/cm2) pode estar relacionada ao grande número de tricomas aciculares longos neste material tipo A1 (31,7 tricomas/4,9 mm2). O contrário pode ser considerado quanto ao genótipo menos preferido, G13028 (1,1 ovos/cm2), que possui principalmente tricomas unciformes curtos tipo B3 (64,1 tricomas/4,9 mm2) em suas folhas.
Palavras-chave: Insecta, Phaseolus vulgaris, mosca-branca, antixenose
ABSTRACT
The oviposition preference (free-choice tests) of Bemisia tabaci (Genn.) Biotype B for 20 bean genotypes was tested. The genotypes were divided into two plots and set up in a block-randomized design with 20 replications. New expanded bean leaves were sectioned and placed in glass tubes containing distilled water and disposed in acrylic cages containing approximately 500 insects. After 24h, the total number of eggs laid on the abaxial surface of leaves and the density of straight and hooked trichomes in two areas of 4.9 mm2 on the abaxial surface were counted. The leaf area was measured as well as the maximum and minimum length of hooked and straight trichomes, which were classified in short, medium or long. No significant correlation was found between the number of eggs and the density of total trichomes (straight + hooked) or between hooked or straight trichomes. Significant correlation was detected between the number of eggs and long straight trichomes type A1 (r = 2.85) and short hooked trichomes type B3 (r = -2.44). In this way, the ovipositional preference of B. tabaci for Bolinha genotype (4.8 eggs/cm2) in a free-choice test could be associated to the larger number of long straight trichomes type A1 (31.7 trichomes/4.9 mm2) on its leaves surface. In addition, the whitefly ovipositional nonpreference for G13028 wild genotype (1.1 eggs/cm2) could be related with the numerous short hooked trichomes type B3 (64.1 trichomes/4.9 mm2) on its leaves surface.
Key words: Insecta, Phaseolus vulgaris, whitefly, antixenosis
O Brasil é considerado um dos maiores produtores mundiais de feijão, Phaseolus vulgaris L., que é uma das principais fontes protéicas da dieta do brasileiro. A mosca-branca Bemisia tabaci (Genn.) é um dos insetos que mais prejudica esta cultura.
Grandes perdas na produção de hortaliças, feijão, soja, amendoim, algodão e várias plantas ornamentais (Lourenção & Nagai 1994, França et al. 1996) têm sido associadas ao ataque do biótipo B de B. tabaci.
O melhoramento de germoplasmas visando à obtenção de cultivares resistentes a Bemisia spp. é um dos mais promissores campos de pesquisa para reduzir as perdas associadas à mosca-branca (McAuslane et al. 1996). Várias características fisiológicas e morfológicas têm sido relacionadas com a incidência de mosca-branca, entre as quais incluem-se os tricomas presentes na superfície foliar das plantas, os quais podem interferir na oviposição, fixação e alimentação dos insetos. Geralmente, a densidade de tricomas tem sido associada com a preferência para oviposição de B. tabaci em diversas culturas, incluindo-se o feijoeiro (Peña et al. 1992, Peña et al. 1993, Oriani & Lara 2000).
Em feijoeiro, são encontrados três diferentes tipos de tricomas: o acicular (em forma de agulha) que é longo, ereto, distalmente estreitado e formado de duas células basais e uma célula terminal; o unciforme (em forma de gancho) que é menor e possui duas células basais e uma célula terminal; e o glandular que é curto. Tais características foram primeiramente descritas por Moutt, em 1932, de acordo com Dahlin et al. (1992).
A densidade de tricomas em feijoeiro pode variar com a sua localização na folha (em geral, a região periférica é mais pilosa), com a superfície foliar (abaxial e adaxial), com o tipo de folha (primária, secundária, terciária, quaternária) e com o genótipo (Quiring et al. 1992).
Quanto à preferência para oviposição de B. tabaci em feijoeiro, Peña et al. (1992) constataram que os genótipos 27-R e PC-50, com maior densidade de tricomas unciformes e com tricomas aciculares mais longos, foram mais ovipositados que os genótipos A-429 e DOR-303, que possuem tricomas aciculares mais curtos e menor densidade de tricomas unciformes. No entanto, Peña et al. (1993) relataram que os tricomas aciculares e unciformes não constituem uma defesa efetiva para o feijoeiro contra a oviposição de B. tabaci, devendo existir outros fatores que atuam na maior preferência desse inseto pelos genótipos 27-R e PC-50. Oriani & Lara (2000) mencionaram que a preferência para oviposição de B. tabaci biótipo B pelo genótipo de feijoeiro Bolinha poderia estar relacionada ao grande número de tricomas aciculares presentes em suas folhas.
Desta forma, este trabalho tem como objetivo verificar a influência dos tricomas presentes na superfície foliar de 20 genótipos de feijoeiro na preferência para oviposição do biótipo B de B. tabaci.
Material e Métodos
Os ensaios de preferência foram conduzidos em laboratório, à temperatura de 23 ± 2ºC, umidade relativa de 70 ± 10% e fotofase de 13h.
Foram avaliados 20 genótipos de P. vulgaris, sendo 13 genótipos selvagens (Arc 1s, Arc 3s, Arc 5s, G11025C, G11056, G12856, G12856B, G12857, G12895, G13028, G23425C1, G23425C6, G23425E), quatro genótipos em melhoramento (Arc 1, Arc 2, Arc 3, Arc 4) e as cultivares (Porrillo 70, IAPAR MD 808 e Bolinha).
Os genótipos foram divididos em dois lotes: lote 1 - Arc 1s, Arc 3s, Arc 5s, Arc 1, Arc 2, Arc 3, Arc 4, Porrillo 70 e IAPAR MD 808; lote 2 - G11025C, G11056, G12856, G12856B, G12857, G12895, G13028, G23425C1, G23425C6 e G23425E. A cultivar Bolinha (testemunha) foi incluída nos dois lotes.
Para a semeadura, foram utilizados sacos plásticos, contendo três partes de terra, uma parte de areia e uma parte de composto orgânico. As plantas foram irrigadas diariamente e mantidas em estufa. A adubação foi realizada no dia da semeadura nos níveis recomendados para a cultura.
A criação de mosca-branca iniciou-se a partir de uma população de B. tabaci (biótipo B) adquirida no Instituto Agronômico de Campinas, sendo os insetos criados em soja, amendoim-bravo e bico-de-papagaio, dentro de estufa em telado tipo anti-afídeo.
Para a infestação dos genótipos de feijoeiro, foram utilizadas gaiolas acrílicas de 40 x 33 x 33 cm, com o fundo e as bordas laterais de alumínio. Aberturas retangulares nas laterais (10 x 10 cm), cobertas com tela anti-afídeo, permitiam a aeração no interior das gaiolas. Folhas seccionadas dos genótipos de feijoeiro foram introduzidas em canudos plásticos de 9 cm de comprimento, e então colocadas em tubos de vidro (8,5 cm de altura e 2,5 cm de diâmetro) com água destilada. Os tubos foram fechados com filme de PVC, o qual era perfurado para a introdução do sistema canudo-folha.
Os 10 tubos de vidro com as folhas de feijoeiro do primeiro lote (e os 11 do segundo) foram distribuídos de forma eqüidistante, ao acaso, em grades de arame, a qual foi colocada no interior da gaiola acrílica sobre quatro tubos de PVC (20 cm de altura e 13 cm de diâmetro), a fim de manter o feijoeiro na parte superior da gaiola. Um frasco de vidro contendo 500 insetos foi colocado no fundo da gaiola. Como a mosca-branca possui fototropismo positivo, à medida que saiam do frasco, os adultos subiam e entravam em contato com as plantas localizadas na parte superior.
Após 24h, foi contado o número total de ovos na superfície abaxial, com auxílio de microscópio estereoscópico. Mediu-se também a área foliar das folhas utilizadas.
O delineamento experimental foi em blocos ao acaso, com 20 repetições de um folíolo (genótipos em melhoramento e cultivares) e de um trifolíolo (genótipos selvagens) recém-expandido por gaiola, para todos os genótipos de feijoeiro dos dois lotes. A utilização de um trifolíolo para os genótipos selvagens teve como objetivo homogeneizar a área foliar entre os materiais avaliados.
Contagem de Tricomas. Com o auxílio de um microscópio estereoscópico com câmara clara, aumento de 80x, contou-se o número de tricomas aciculares e unciformes presentes na superfície abaxial em uma área de 4,9 mm2, em duas regiões da folha de feijoeiro: uma área proximal (eqüidistante entre a nervura principal e a margem foliar) e uma área distal em relação ao pecíolo. A densidade de tricomas foi calculada como a densidade média entre as duas regiões. O método foi adaptado daquele utilizado por McAuslane (1996).
Como não existe classificação dos tricomas presentes em feijoeiros quanto ao comprimento, está sendo proposta uma tipologia de acordo com a forma e o tamanho: tipo A para os tricomas aciculares e tipo B para os unciformes. Os tricomas foram subdivididos em: acicular longo (A1), acicular médio (A2) e acicular curto (A3) e unciforme longo (B1), unciforme médio (B2) e unciforme curto (B3). Assim, além da contagem da densidade total de tricomas aciculares e unciformes, realizaram-se contagens dos seis diferentes tipos de tricomas anteriormente descritos, em 10 das 20 repetições realizadas. Medidas máximas e mínimas de cada um dos diferentes tipos de tricomas também foram registradas.
Para a análise estatística, foram aplicados os testes F e de Tukey, a 5% de probabilidade de erro para o número ovos de B. tabaci por cm2 e correlações lineares simples entre o número de ovos/cm2 e os números de tricomas (totais, unciformes e aciculares) e os números dos diferentes tipos de tricomas (A1, A2, A3, B1, B2 e B3).
Resultados e Discussão
Com base nas medidas máximas e mínimas de cada um dos tipos de tricomas presentes na superfície foliar de feijoeiros (Tabela 1), determinou-se que o tricoma acicular tipo A1 é o mais longo, geralmente encontrado nas nervuras dos folíolos, possuindo comprimento máximo de 1.050 mm e mínimo de 790 mm, seguido do tipo A2, variando de 780 a 500 mm e do tipo A3, que é o tricoma acicular mais curto, com medida máxima de 490 mm e mínima de 360 mm. Para o tricoma unciforme, tem-se como mais longo o tipo B1, com comprimento máximo de 490 mm e mínimo de 350 mm, seguido do tipo B2, com valores variando de 340 a 210 mm e o tipo B3, o mais curto dos tricomas avaliados nos feijoeiros, com medidas máximas de 200 mm e mínimas de 90 mm.
Segundo Doutt (1932), citado por Dahlin et al. (1992), para a cultivar Black Valentine, o comprimento do tricoma acicular é de 400 mm e do tricoma unciforme, de 200 mm. Comparando-se os dados de Doutt com a classificação apresentada neste trabalho (Tabela 1), observa-se que a cultivar Black Valentine teria tricoma acicular do tipo A3 e tricoma unciforme do tipo B3, que são os mais curtos dentro de suas categorias, acicular e unciforme, respectivamente.
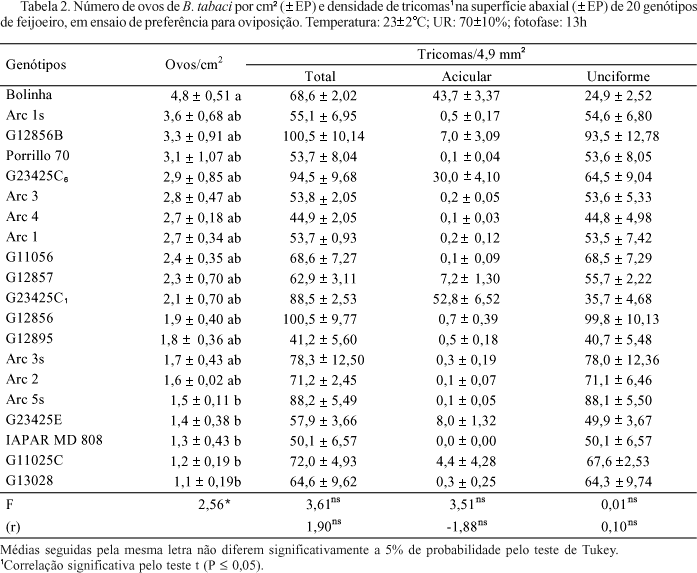
Quanto à oviposição de B. tabaci biótipo B, observa-se que o genótipo mais preferido foi o Bolinha (4,8 ovos/cm2); entre os menos preferidos podem-se relacionar G13028, G11025C, IAPAR MD808, G23425E e Arc 5s (médias variando de 1,1 a 1,5 ovos/cm2), valores estes que diferiram significativamente dos constatados na cultivar Bolinha (Tabela 2).
Oriani et al. (2005) também observaram que o genótipo selvagem G13028 mostrou-se altamente não-preferido para oviposição por B. tabaci biótipo B, em ensaios com e sem chance de escolha (0,9 e 1,0 ovos/cm2, respectivamente). Citaram também como resistentes à mosca-branca, em ensaios com e sem chance de escolha, os genótipos Arc 3s (0,9 e 1,0 ovos/cm2, respectivamente), Arc 5s (1,1 e 1,3 ovos/cm2, respectivamente) e G11056 (2,4 e 1,1 ovos/cm2, respectivamente); já o genótipo Bolinha (7,9 e 2,2 ovos/cm2, respectivamente) mostrou-se altamente suscetível.
As maiores densidades de tricomas (aciculares + unciformes) ocorreram nos genótipos G12856B, G12856, G23425C6, G23425C1 e Arc 5s (médias variando de 100,5 a 88,2 tricomas/4,9 mm2) e as menores densidades, nos genótipos G12895, Arc 4, IAPAR MD 808 (médias variando de 41,2 a 50,1 tricomas/4,9 mm2) (Tabela 2). Quanto aos tricomas aciculares, pode-se relacionar as maiores densidades para os genótipos G23425C1, Bolinha, G23425C6 (52,8, 43,7 e 30,0 tricomas aciculares/4,9 mm2, respectivamente). Nos demais materiais testados, os valores não ultrapassaram 8,0 tricomas aciculares/4,9 mm2, sendo que 13 dos 20 materiais avaliados apresentaram valores próximos a zero para os tricomas aciculares. Os tricomas unciformes são os mais comuns para todos os feijoeiros testados, sendo as maiores densidades observadas para G12956, G12956B e Arc 5s (médias variando de 99,8 a 88,1 tricomas unciformes/4,9 mm2) e as menores densidades para Bolinha, G23425C1 e G12895 (médias variando de 24,9 a 40,7 tricomas unciformes/4,9 mm2) (Tabela 2).
Não foram constatadas correlações lineares entre o número de ovos e a densidade total de tricomas (aciculares + unciformes) e de tricomas aciculares e unciformes presentes na superfície abaxial dos 20 genótipos de feijoeiros (Tabela 2). Desta forma, pode-se sugerir que a densidade de tricomas total (unciformes + aciculares) e de tricomas aciculares e unciformes não está relacionada com a preferência para oviposição da mosca-branca em feijoeiro.
Vários autores mencionaram que a alta densidade de tricomas está positivamente correlacionada com a oviposição de B. tabaci em algodoeiro (Wilson et al. 1993, Chu et al. 2000, Chu et al. 2001), em tomateiro (Heinz & Zalom 1995, Toscano et al. 2002) e em soja (McAuslane et al. 1995, McAuslane 1996, Valle & Lourenção 2002), já que genótipos altamente pilosos poderiam fornecer um microclima mais adequado para a oviposição das fêmeas e uma melhor proteção às ninfas (Butter & Vir 1989), além de as fêmeas preferirem colocar os ovos na base de inserção dos tricomas (Omran & El-Khidir 1978).
No entanto, a correlação positiva entre a oviposição da mosca-branca e a densidade de tricomas não é constante para todas as plantas hospedeiras. Assim, pubescência de folhas foi negativamente correlacionada com a população de moscas-brancas, em várias espécies de cucurbitáceas (McCreight & Kishaba 1991) e em Lagenaria siceraria (Kishaba et al. 1992). Isto também foi observado no presente trabalho em que não houve correlação positiva entre a densidade total de tricomas e a oviposição de B. tabaci biótipo B.
Os tricomas aciculares longos (tipo A1) estão presentes nos genótipos Bolinha, G23425C6 e G23425C1 (médias variando de 31,7 a 12,0 tricomas/4,9 mm2) e praticamente ausentes nos demais genótipos; 13 dos 20 genótipos testados não apresentaram nenhum tricoma acicular longo (tipo A1). Os tricomas aciculares médios (tipo A2) e os tricomas aciculares curtos (tipo A3) estão presentes em maior densidade no genótipo G23425C1 (33,2 e 19,3 tricomas/4,9 mm2, respectivamente); no entanto, os genótipos Bolinha, G23425C6, G23425E, G12957 também apresentaram estes dois tipos de tricomas (médias variando de 8,0 a 2,3 tricomas/4,9 mm2); os demais genótipos apresentaram valores próximos de zero (Tabela 3).
Quanto aos tricomas unciformes longos tipo B1 (Tabela 4), as maiores densidades foram constatadas nos genótipos G23425C6, G23425C1 e G12856B (46,9, 18,5 e 16,4 tricomas/4,9 mm2, respectivamente); os genótipos G12985, G23425E, G12857, Bolinha e G12856 apresentaram médias variando de 7,4 a 3,8 tricomas/4,9 mm2 e os demais materiais, valores próximos a zero. Para os tricomas unciformes médios (tipo B2), os genótipos G12856 e G12856B apresentaram valores próximos a 45 tricomas/4,9 mm2; sete genótipos apresentaram valores médios variando de 8,8 a 21,5 tricomas/4,9 mm2 e os demais valores próximos de zero. Os tricomas unciformes curtos (tipo B3) foram constatados em todos os materiais testados, sendo as maiores densidades observadas em Arc 5s, G11056 e G13028 (84,3, 69,7 e 64,1 tricomas/4,9 mm2, respectivamente) e as menores densidades, em G23425C1, Bolinha e G23425C6 (4,7, 9,2 e 15,1 tricomas/4,9 mm2, respectivamente); os demais materiais apresentaram valores intermediários (Tabela 4).
Foram observadas correlações significativas entre o número de ovos e a densidade de tricomas aciculares longos (tipo A1) (r = 2,85, Tabela 3) e a densidade de tricomas unciformes curtos (tipo B3) (r = -2,44, Tabela 4).
Desta forma, o fator comprimento dos tricomas deve estar influenciando a preferência para oviposição da mosca-branca, uma vez que houve correlação positiva, para tricomas aciculares longos (tipo A1) e negativa, para tricomas unciformes curtos (tipo B3). Os tricomas aciculares longos (tipo A1), possuem comprimento máximo de 1.050 mm e mínimo de 790 mm, enquanto para os tricomas unciformes curtos (tipo B3), tais valores foram 200 mm e 90 mm, respectivamente (Tabela 1). Assim, a preferência para oviposição da mosca-branca pela cultivar Bolinha (4,8 ovos/cm2) pode estar relacionada ao grande número de tricomas aciculares longos presentes neste material (31,7 tricomas/4,9 mm2) (Tabela 3). Entretanto, como a correlação positiva constatada para os tricomas aciculares longos foi proveniente de apenas três genótipos, Bolinha (31,7), G23445C6 (15,8) e G23445C1 (12,0) dos 20 testados, propõe-se que sejam testados mais materiais com tricomas aciculares longos, a fim de comprovar essa constatação. O contrário pode ser considerado quanto ao genótipo menos preferido, G13028 (1,1 ovos/cm2), uma vez que possui principalmente tricomas unciformes curtos (64,1 tricomas/4,9 mm2) (Tabela 4).
Peña et al. (1993) também verificaram que B. tabaci manifestou preferência pelos genótipos de feijoeiro com tricomas aciculares mais longos (27-R e PC-50), enquanto aqueles que possuem tricomas aciculares mais curtos (A-429 e o DOR-303) tiveram as menores oviposições. Oriani & Lara (2000) relataram que a preferência para oviposição de B. tabaci biótipo B pela cultivar Bolinha poderia estar associada ao grande número de tricomas aciculares presentes em suas folhas, porém os autores não verificaram os comprimentos destes tricomas.
Desta forma, sugere-se que, para programas de melhoramento de feijoeiro que visem obter germoplasma com resistência a B. tabaci biótipo B, sejam selecionados materiais com tricomas mais curtos, já que estes podem atuar na não-preferência para oviposição pelo referido inseto.
Agradecimentos
Os autores agradecem à Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP), pela concessão de bolsa de pós-doutorado ao primeiro autor e de Bolsa de Auxílio à Pesquisa para este Projeto; ao Dr. André Luiz Lourenção (IAC), pelo fornecimento da população inicial de mosca-branca; e ao CNPAF/EMBRAPA, pelo fornecimento dos genótipos de feijoeiro.
Literatura Citada
Received 15/XII/03. Accepted 14/XII/04.
- Butter, N.S. & B.K. Vir. 1989. Morphological basis of resistance in cotton to the whitefly Bemisia tabaci Phytoparasitica 17: 251-261.
- Chu, C.C., T.P. Freeman, J.S. Buckner, E.T. Natwick, T.J. Henneberry & D.R. Nelson. 2000. Silverleaf whitefly colonization and trichome density relationships on upland cotton cultivars. Southwestern Entomol. 25: 237-242.
- Chu, C.C., T.P. Freeman, J.S. Buckner, T.J. Henneberry, D.R. Nelson & E.T. Natwick. 2001. Susceptibility of upland cotton cultivars to Bemisia tabaci biotype B (Homoptera: Aleyrodidae) in relation to leaf age and trichome density. An. Entomol. Soc. Am. 94: 743-749.
- Dahlin, R.M., M.A. Brick & J.B. Ogg. 1992. Characterization and density of trichomes on three common bean cultivars. Econ. Bot. 46: 299-304.
- França, F.H., G.L. Villas Bôas & M. Castelo Branco. 1996. Ocorrência de Bemisia argentifolii Bellows & Perring (Homoptera, Aleyrodidae) no Distrito Federal. An. Soc. Entomol. Bras. 25: 369-372.
- Heinz, K.M. & F.G. Zalom. 1995. Variation in trichome-based resistance to Bemisia argentifolii (Homoptera, Aleyrodidae) oviposition on tomato. J. Econ. Entomol. 88: 1494-1502.
- Kishaba, A.N., S. Castle, J.D. McCreight & P.R. Desjardins. 1992. Resistance of white-flowered gourd to sweetpotato whitefly. Hortscience 27: 1217-1221.
- Lourenção, A.L. & H. Nagai. 1994. Surtos populacionais de Bemisia tabaci no estado de São Paulo. Bragantia 53: 53-59.
- McAuslane, H.J. 1996. Influence of leaf pubescence on ovipositional preference of Bemisia argentifolii (Homoptera, Aleyrodidae) on soybean. Environ. Entomol. 25: 834-841.
- McAuslane, H.J., F.A. Johnson, D.L. Colvin & B. Sojack. 1995. Influence of foliar pubescence on abundance and parasitism of Bemisia argentifolii (Homoptera, Aleyrodidae) on soybean and peanut. Environ. Entomol. 24: 1135-1143.
- McAuslane, H.J., S.E. Webb & G.W. Elmstrom. 1996. Resistance in germplasm of Cucurbita pepo to silverleaf, a disorder associated with Bemisia argentifolii (Homoptera, Aleyrodidae). Fla. Entomol. 79: 206-221.
- McCreight, J.D. & A.N. Kishaba. 1991. Reaction of cucurbit species to squash leaf curl virus and sweet-potato whitefly. J. Am. Soc. Hortic. Sci. 116: 137-141.
- Omram, H.H. & E. El-Khidir. 1978. On the preference of Bemisia tabaci Genn. (Homoptera, Aleyrodidae) on various cotton cultivars in Cukurova, Turkey. Agric. Ecosyst. Environ. 17: 83-88.
- Oriani, M.A.G. de & F.M. Lara. 2000. Oviposition preference of Bemisia tabaci (Genn.) biotype B (Homoptera: Aleyrodidae) for bean genotypes containing arcelin in the seeds. An. Soc. Entomol. Brasil 29: 565-572.
- Oriani, M.A.G. de, J.D. Vendramin & Brunherotto. 2005. Atratividade e não-preferência para oviposição de Bemisia tabaci (Genn.) biótipo b (Hemiptera: Aleyrodidae) em genótipos de feijoeiro. Neotrop. Entomol. 34(1): 105-111.
- Peña, E.A., A. Pantoja & J. Beaver. 1992. Determinación de la pubescencia de cuatro genotipos de habichuela, Phaseolus vulgaris L. J. Agric. Univ. P. R. 76: 71-82.
- Peña, E.A., A. Pantoja, J. Beaver & Armstrong, A. 1993. Oviposicion de Bemisia tabaci Genn. (Homoptera, Aleyrodidae) en cuatro genotipos de Phaseolus vulgaris L. (Leguminosae) con diferentes grados de pubescencia. Folia Entomol. Mex. 87: 1-12.
- Quiring, D.T., P.R. Timmins & S.J. Park. 1992. Effect of variations in hooked trichome densities of Phaseolus vulgaris on longevity of Liriomyza trifolii (Diptera: Agromyzidae) adults. Environ. Entomol. 21: 1357-1361.
- Toscano, L.C., A.L. Boiça Júnior & W.I. Maruyama. 2002. Nonpreference of whitefly for oviposition in tomato genotypes. Sci. Agric. 59: 677-681.
- Valle, G.E. do & A.L. Lourenção. 2002. Resistência de genótipos de soja a Bemisia tabaci (Genn.) biótipo B (Hemiptera: Aleyrodidae). Neotrop. Entomol. 31: 285-295.
- Wilson, F.D., H.M. Flint, B.R. Stapp & N.J. Parks. 1993. Evaluation of cultivars, germplasm lines, and species of Gossypium for resistance to biotype "B" of sweetpotato whitefly (Homoptera, Aleyrodidae). J. Econ. Entomol. 86: 1857-1862.
Datas de Publicação
-
Publicação nesta coleção
22 Mar 2005 -
Data do Fascículo
Fev 2005
Histórico
-
Recebido
15 Dez 2003 -
Aceito
14 Dez 2004