Abstracts
For the first time all instars of a species of the genus Belminus are described based on laboratory F1 generation specimens from adults caught in department of Cesar, Colombia. Ontogenetic morphometrics of postembryonic head development was made, suggesting that the greatest changes occur between the 1st and 2nd instars.
Chagas disease; vector; Bolboderini; systematics
Pela primeira vez todos estádios ninfais de uma espécie do gênero Belminus são descritos, baseados em espécimes da primeira geração obtida em laboratório a partir de adultos capturados em domicílios do Departamento de Cesar, Colômbia. A morfometria geométrica das cabeças sugere que as maiores mudanças no desenvolvimento pós-embrionário ocorrem entre o 1º e o 2º ínstares.
Doença de Chagas; vetor; Bolboderini; sistemática
PUBLIC HEALTH
Description and ontogenetic morphometrics of nymphs of Belminus herreri Lent & Wygodzinsky (Hemiptera: Reduviidae, Triatominae)
Descrição e morfometria ontogenética das ninfas de Belminus herreri Lent & Wygodzinsky (Hemiptera: Reduviidae, Triatominae)
Dayse da S. RochaI; James S. PattersonII; Claudia M. SandovalIII; José JurbergI; Victor M. ÂnguloIII; Lyda Esteban A.III; Cleber GalvãoI
ILab. Nacional e Internacional de Referência em Taxonomia de Triatomíneos, Instituto Oswaldo Cruz, FIOCRUZ Av. Brasil 4365, 21040-900, Rio de Janeiro, RJ, Brazil
IIPathogen Molecular Biology Unit, London School of Hygiene and Tropical Medicine, London, UK
IIICentro de Investigaciones en Enfermedades Tropicales, Universidad Industrial de Santander, CINTROP, UIS Santander, Colombia
ABSTRACT
For the first time all instars of a species of the genus Belminus are described based on laboratory F1 generation specimens from adults caught in department of Cesar, Colombia. Ontogenetic morphometrics of postembryonic head development was made, suggesting that the greatest changes occur between the 1st and 2nd instars.
Key words: Chagas disease, vector, Bolboderini, systematics
RESUMO
Pela primeira vez todos estádios ninfais de uma espécie do gênero Belminus são descritos, baseados em espécimes da primeira geração obtida em laboratório a partir de adultos capturados em domicílios do Departamento de Cesar, Colômbia. A morfometria geométrica das cabeças sugere que as maiores mudanças no desenvolvimento pós-embrionário ocorrem entre o 1º e o 2º ínstares.
Palavras-chave: Doença de Chagas, vetor, Bolboderini, sistemática
The reduviid subfamily Triatominae is organized into 18 genera and six tribes, with 136 species, excluding of the checklist of Triatominae (Galvão et al. 2003), Torrealbaia martinezi Carcavallo, Jurberg & Lent, 1998, recently synonymized with Amphibolus Klug, 1839 (Harpactorinae) by Forero et al. (2004). The majority of species are found in the Americas, and are potential or actual vectors of Trypanosoma cruzi, agent of Chagas disease (American trypanosomiasis).
After the successful large-scale campaign to control domestic populations of Triatominae in some South American countries, it is very important to maintain surveillance of species invading controlled areas. In fact, in recent years, there have been increasing reports of sylvatic species invading human dwellings and peridomestic structures in South American countries (Valente 1999, Galvão et al. 2001, Vivas et al. 2001). More surprising is the colonization of peridomestic environment or human dwellings by species previously considered exclusively sylvatic, that is the case of Microtriatoma trinidadensis (Lent, 1951) caught from peridomestic sites in Bolivia (De la Riva et al. 2001) and Belminus herreri Lent & Wygodzinsky, 1979 colonizing human dwellings in Colombia (Sandoval et al. 2000, 2004).
The genus Belminus (Bolboderini) is composed of the following six species: B. rugulosus Stål, 1859, B. costaricensis Herrer, Lent & Wygodzinsky, 1954, B. peruvianus Herrer, Lent & Wygodzinsky, 1954, B. herreri, B. pittieri Osuna & Ayala, 1993, and B. laportei Lent, Jurberg & Carcavallo, 1995. These species are restricted to Central America, Colombia, Peru, Venezuela, and northern Brazil. Species of this genus are considered sylvatic and arboreal, associated with opossums, lizards, and arthropods (Lent & Wygodzinsky 1979). B. herreri was described based on specimens collected in the Panamanian rainforest. After the initial description, this species was not reported again until 2000, when it was found for the first time in a domestic environment in Colombia (Sandoval et al. 2000). Systematic studies of this genus are restricted to adults (Lent & Wygodzinsky 1979) and male phallic structures (Lent & Jurberg 1984).
In order to obtain more understanding of immature forms of triatomine species, we describe here for the first time the five instars of B. herreri, including an ontogenetic morphometric study of head development.
Material and Methods
Insects. The specimens used in this study were laboratory F1 generation specimens from adults from dwellings from the Department of Cesar, municipality of San Martín, Colombia.
Optical Microscopy. Nymphs were warmed in a 10% solution of KOH, dehydrated in phenol, and mounted on slides. The drawings were made with a camera lucida coupled to a stereoscopic microscope.
Ontogenetic Morphometrics. The dorsal surfaces of the heads of specimens of a sample of each instar nymph and adults (three 1st, two 2nd, three 3rd, one 4th, four 5th and four adults) were photographed using a digital camera. Digital images of all specimens were subsequently digitised and seven Cartesian coordinates (landmarks) (Fig. 1) were recorded automatically using TPSdig software, version 1.39 (Rholf 2001). The x y coordinates were subjected to procrustes superimposition (Bookstein 1991) and subsequently to a Thin Plate Spline analysis using TPSrelw software,version 1.35 (Rholf 2001). The analysis produces variables subdivided into uniform and non-uniform components of shape changes. A principal component analysis of the shape components delivers tangible shape components ("relative warps"). Multivariate analyses and graphs were completed using JMP® version 4.0.5 (SAS Institute Inc. 2000) and Intercooled STATA 8.2 for Windows (CRC 1992).
Results
Ontogenetic Morphometrics.Fig. 1 shows the discrimination of the five instars and adults. The first two principal components together account for 96.% of the total heterogeneity (PC1- 92.4% and PC2 - 4.1%). PC1 represents most of the shape change and is obviously correlated to size. The thin plate spline representations a and b show that most of the shape change is associated with a relative elongation of the head and enlargement of the eye. PC2, representing a far lesser component of the shape change demonstrates that first instars and adults have similar values, separate from the other instars. The representations c and d show that this subtle secondary factor of shape change is related to the relatively longer postocular distance in adults and 1st instars. Overall, the ontogenic profile of postembryonic head development for this species suggests that the greatest change occurs between the 1st and 2nd instars.
Description
1st Instar. Total length of the body (average) 1.8 mm. Overall color brown light, abdomen brown-orange. Thorax, legs and outer border of abdomen with conspicuous setiferous tubercles and pilosity, setae longer than length of setiferous tubercle. Head short with scarce setiferous granules and more discrete pilosity, postocular region of head with sides convex, abruptly constricted before neck. Ratio of anteocular x postocular regions 1:0.8. Rostrum attaining prosternum, first and second segments subequal in length. Ratio of rostral segments 1:0.9:0.6. Antennae inserted at the apex of third basal part of anteocular region, antenniferous tubercles with apicolateral process with setae inserted, all antennal segments covered with setae. Antennal segments I, II, II, and basal portion of IV sclerotized, apical portion of IV segment more membranous. Antennal segment II with submedian trichobothria. Fourth antennal segment shorter than other three combined. Ratio of antennal segments 1:1.4:1.7:3.3. Anteclypeus rounded, reaching level of middle of the second antennal segment. Genae not reaching level of apex of anteclypeus. Postocular furrow rounded and easily visible. Pronotum the largest thoracic sclerite, with some setiferous tubercles medially and a narrow line along its middle; mesonotum smaller than pronotum but not shorter than half the length of pronotum, longer at midline than at sides; metanotum reduced to 1+1 lateral sclerotized plates with some discal setae inserted, plates narrower than half the distance between them, and with some tubercles medially. Legs with numerous and conspicous setae; underside of femora with conspicuous setiferous denticles, all legs with median rings; tarsi two-segmented. Spongy fossulae absent. Abdomen short, stout, abdominal margins unicolorous without spotted connexival marks; urotergites with tranverse row of setae, external border of connexivum with conspicuous setae (Fig. 2).
2nd Instar. Like 1st instar, except: Total length of the body (average) 2.8 mm. Overall color brown except, pro-, meso- and metanotal plates dark brown. Head slightly more elongate, with more setiferous granules in contrast to 1st instar head. Ratio of anteocular x postocular regions 1:0.6. Ratio of antennal segments 1:1.9:2:3.1. Ratio of rostral segments 1:1:0.6. (Fig. 3). Metanotal plates equal in width to half the distance between them. Setiferous tubercles very prominent and thorny on the external border of head, pronotum, metanotum, mesonotum, and abdomen, like a handsaw. Spotted connexival marks of abdomen somewhat little visible; urotergites with two tranverse rows of setae. (Fig. 4).
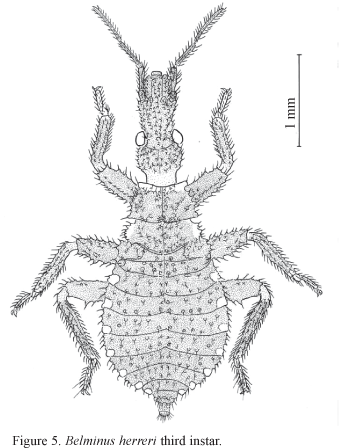
3rd Instar. Like 2nd instar, except: Total length of the body (average) 4.0 mm. Overall color brown except following areas yellow: anterior border of pronotum, outer border of mesonotum, and connexival marks. Head elongate, fusiform, with numerous setiferous granules. Ratio of anteocular x postocular regions 1:0.5. Ratio of antennal segments 1:2:2.5:3.8. Ratio of rostral segments 1:0.9:0.3. Metanotal plates slightly wider than half the distance between them (Fig. 5).
4th Instar. Like 3rd instar, except: Total length of the body (average) 6.4 mm. Ratio of anteocular x postocular regions 1:0.4. Ratio of antennal segments 1:2.2:2:2.7. Ratio of rostral segments 1:1:0.4. Genae almost attaining level of apex of anteclypeus. Metanotal plates linked (Fig. 6).
5th Instar. Like 4th instar, except: Total length of the body (average) 7.5 mm. Overall color dark brown, except following areas yellow: anterolateral and humeral angles of pronotum, apex of wing pads. Head elongate. Genae very prominent, surpassing level of apex of anteclypeus. Ratio of anteocular x postocular regions 1:0.4. Antenniferous tubercles situated almost at center of anteocular region, with conspicuous apicolateral process. Ratio of antennal segments 1:2.2:1.8:2.6. Ratio of rostral segments 1:1:0.4. (Fig. 7).
Discussion
Triatomine systematics is based mainly on exoskeletal characters of the adults and phallic structures of the males, and most species can be recognized this way (Lent & Wygodzinsky 1979, Jurberg et al. 1998). On the other hand, specific identification based on the morphological traits of nymphs of triatomine bugs is not easy. Although there are keys for generic determination of triatomine first and fifth instars (Lent and Wygodzinsky 1979, Galíndez-Girón et al. 1998), the specific identification of all instars is a challenge far from having been solved, because it is difficult to find new characters to use in the diagnosis.
A table listing specific names, used approaches and references of papers dedicated to morphology of triatomine eggs and nymphs is being published by Galvão et al. (in press). This summary shows that the morphological knowledge of eggs and immature forms of triatomine bugs is poor. Regarding eggs it is necessary to refer to the detailed studies of Barata (1980, 1981, 1998). Therefore, morphological studies of immature forms of triatomines can be useful for the early identification of the nymphs of species found in human dwellings or the peridomestic environment, and may allow the application of adequate control of these vectors. On the other hand, systematic and phylogenetic studies can be improved if new characters can be found in immature forms.
The recent finds of Sandoval (2000, 2004) show that B. herreri can adapt to human dwellings. Studies of the intestinal content of specimens caught inside dwellings showed that most of these specimens had fed on cockroaches; however, some specimens also showed the presence of fragments of DNA of T. cruzi in their intestinal content, suggesting that the dwellings are an alternative ecotope for this species and its involvement in epidemiology of Chagas disease (Sandoval et al. 2004).
The subfamily Triatominae is a group of Reduviidae defined by its members' obligatory haematophagy; however, some species are able to feed on arthropods (e.g., the case of B. herreri feeding on Blattidae). The first observations on Triatominae feeding on arthropods were made by Brumpt (1914) and Ryckman (1951). Miles et al. (1981) observed Eratyrus mucronatus Stål, 1859 feeding on spiders and, more recently, laboratory studies with T. circummaculata (Stål, 1859), T. rubrovaria (Blanchard, 1843), and T. carcavalloi Jurberg, Rocha & Lent, 1988, suggest that haemolymphagy is common in these species and probably represents an important survival strategy under natural conditions (Lorosa et al. 2000, Ruas-Neto et al. 2001). This capacity could be interpreted as an adaptation to best utilize available sources of nutrition depending on host availability in the bugs' habitats. Alternatively, according to Schofield (2000), it may suggest a recent adaptation to a blood-feeding strategy. Currently, despite ongoing debate, there is no consensus in the literature as to whether haematophagy developed independently in several lineages of Triatominae or perhaps Reduviidae, or whether a common haematophagous ancestral stock gave rise to all modern triatomine groups. For current views on the controversies of evolutionary relationships of Triatominae see Hypa et al. (2002), Galvão (2003), Schaefer (2003) and Sainz et al. (2004). Although haemolymphagy can be easier (apparently) for the young nymphs of B. herreri, our morphological results show that the reduced size of metanotal plates in young instars could facilitate the ingestion of blood meal as hypothesized by Lent & Wygodzinsky (1979).
Further detailed morphological analysis of other Bolboderini nymphs (as soon as new samples are available) is necessary to compare similarities in the ontogenetic development among the genera of this tribe, allowing not only the differentiation of related species as well as to infer on the phylogenetic relationship.
Acknowledgments
This study was supported by MCT/CNPq, FAPERJ, Chagas disease Intervention Activities (CDIA) and Fiocruz/SVS/Fiotec. An anonymous reviewer helped to improve the manuscript.
Literature Cited
Received 18/VIII/04. Accepted 04/II/05.
- Barata, J.M.S. 1980. Aspectos morfológicos de ovos de triatomíneos. I Mensuração de dez espécies do gênero Rhodnius Stål, 1859 (Hemiptera, Reduviidae). Revta. Bras. Ent. 24: 197-214.
- Barata, J.M.S. 1981. Aspectos morfológicos de ovos de triatomíneos. II Características macroscópicas e exocoriais de dez espécies do gênero Rhodnius Stal, 1859 (Hemiptera, Reduviidae). Rev. Saúde Publ. São Paulo 15: 490-452.
- Barata, J.M.S. 1998. Macroscopic and exocorial structures of Triatominae eggs (Hemiptera, Reduviidae), p.409-448. In R.U. Carcavallo, I. Galíndez-Girón, J. Jurberg & H. Lent (eds.), Atlas of Chagas disease vectors in the Americas, v. II, Editora Fiocruz, Rio de Janeiro, 733p.
- Bookstein, F.L. 1991. Morphometric tools for landmark data: Geometry and biology. Cambridge Univ. Press, New York, 80p.
- Brumpt, E. 1914. Importance du cannibalisme et de la coprofhagie chez les Réduvidés hématophages (Rhodnius, Triatoma) pour la conservation des Trypanosomes pathogénes en dehors de I'hote vertébré. Bull. Soc. Pathol. Exot. 7: 702-705.
- Computing Resource Center - CRC1992. Stata reference manual: Release 3, 5th ed. Computing resource center, Santa Monica, CA.
- Forero, D., C. Weirauch, & M. Baena. 2004. Synonymy of the reduviid (Hemiptera:Heteroptera) genus Torrealbaia (Triatominae) with Amphibolus (Harpactorinae), with notes on Amphibolus venator (Klug, 1830). Zootaxa 670: 1-12.
- Galíndez-Girón, I., D. da S.Rocha, H. Lent, R.U. Carcavallo, J. Jurberg, C. Galvão, H.S. Santos, A. Martinez, J.M.S. Barata & J.A. Rosa 1998. Nymphal stages, p.449-513. In R.U. Carcavallo, I. Galíndez-Girón, J. Jurber & H. Lent (eds.), Atlas of Chagas disease vectors in the Americas, vol. II, Editora Fiocruz, Rio de Janeiro, 733p.
- Galvão, C. 2003. A sistemática dos triatomíneos (Hemiptera, Reduviidae), de De Geer ao DNA. Entomol. Vect. 10: 511-530.
- Galvão, C., D. da S. Rocha, J. Jurberg & R.U. Carcavallo 2001. Ampliação da distribuição geográfica de Triatoma deaneorum Galvão, Souza & Lima, 1967, nova denominação para Triatoma deanei (Hemiptera, Reduviidae). Rev. Soc. Bras. Med. Trop. 34: 587-589.
- Galvão, C., F.M. McLoon, D. da S. Rocha, C.W. Schaefer, J. Patterson & J. Jurberg. Description of eggs and nymphs of Linshcosteus karupus Galvão, Patterson, Rocha, & Jurberg, 2002 (Hemiptera: Reduviidae: Triatominae). Ann. Entomol. Soc. Am 98 (in press)
- Galvão, C., R.U. Carcavallo, D.S. Rocha & J. Jurberg. 2003. A checklist of the current valid species of the subfamily Triatominae Jeannel, 1919 (Hemiptera, Reduviidae) and their geographical distribution, with nomenclatural and taxonomic notes. Zootaxa 202: 1-36.
- Hypa, V., D.F. Tietz, J. Zrzavý, R.O.M. Rego, C. Galvão & J. Jurberg. 2002. Phylogeny and biogeography of Triatominae (Hemiptera: Reduviidae): molecular evidence of a New World origin of the Asiatic clade. Mol. Phyl. Evol. 23: 447-457.
- Jurberg, J., H. Lent & C. Galvão. 1998. The male genitalia and its importance in taxonomy, p.85-106. In R.U. Carcavallo, I. Galíndez-Girón, J. Jurberg & H. Lent (eds.), Atlas of Chagas disease vectors in the Americas, vol. I, Editora Fiocruz, Rio de Janeiro, 393p.
- Lent, H. & J. Jurberg. 1984. A genitália externa na tribo Bolboderini (Hemiptera, Reduviidae, Triatominae). Mem. Inst. Oswaldo Cruz 79: 1-27.
- Lent, H. & P. Wygodzinsky. 1979. Revision of the Triatominae (Hemiptera, Reduviidae), and their significance as vectors of Chagas´disease. Bull. Am. Mus. Nat. Hist. 163: 123-520.
- Lorosa, E.S., J. Jurberg, A.L. Souza, M.C. Vinhaes & I. Nunes. 2000. Hemolinfa de Dictyoptera na manutenção do ciclo biológico silvestre de Triatoma rubrovaria (Blanchard, 1843) e Triatoma circummaculata (Stal, 1859) (Hemiptera, Reduviidae, Triatominae) Entomol. Vect. 7: 287-296.
- Miles, A., J.R. Arias, A. de Souza & M. Póvoa. 1981. Chagas disease in the Amazon basin III. Ecotopes of ten triatomine bug species (Hemiptera: Reduviidae) from the vicinity of Belém, Para state, Brazil. J. Med. Entomol. 18: 266-278.
- Riva, J. De la, A. Matias, M. Torrez, E. Martinez & J.P. Dujardin. 2001. Adults and nymphs of Microtriatoma trinidadensis (Lent, 1951) (Hemiptera:Reduviidae) caught from peridomestic environment in Bolivia. Mem. Inst. Oswaldo Cruz 96: 889-894.
- Rohlf, F.J. 2001. NTSYS. Numerical taxonomy and multivariate analysis system.Version 2.10p, Exeter software, Setauket, New York.
- Ruas-Neto, A.L., E. Corseuil & A. Cavalleri. 2001. Development of rupestrian triatomines (Hemiptera: Reduviidae: Triatominae) following hemolymphagy on Blaberids (Blattodea: Blaberidae) in Rio Grande do Sul State, Brazil. Entomol. Vect. 8: 205-216.
- Ryckman, R.E. 1951. Recent observations of cannibalism in Triatoma (Hemiptera: Reduviidae). J. Parasitol. 37: 433-434.
- Sainz, A.C., L.V. Mauro, E.N. Moriyama & B.A. García. 2004. Phylogeny of triatomine vectors of Trypanosoma cruzi suggested by mitochondrial DNA sequences. Genetica 121: 229-240.
- Sandoval, C.M., M. Joya, M. Gutiérrez & V.M. Angulo. 2000. Cleptohaemathophagia in B. herreri Med. Vet. Entomol. 14: 100-101.
- Sandoval, C.M., R. Duarte, R. Gutíerrez, D. da S. Rocha, V.M. Angulo, L. Esteban, M. Reyes, J. Jurberg & C. Galvão. 2004. Feeding sources and natural infection of Belminus herreri (Hemiptera, Reduviidae, Triatominae) from dwellings in Cesar, Colombia. Mem. Inst. Oswaldo Cruz 99: 137-140.
- SAS Institute Inc. 2000. JMP® Statistics and graphics guide. Version 4.0.4. SAS Campus Drive, Cary.
- Schaefer, C.W. 2003. Triatominae (Hemiptera: Reduviidae): Systematic Questions and Some Others. Neotrop. Entomol. 32: 1-10.
- Schofield, C.J. 2000. Biosystematics and evolution of the Triatominae. Cad. Saúde Pública 16: 89-92.
- Valente, V.C. 1999. Potential for domestication of Panstrongylus geniculatus (Latreille, 1811) (Liemiptera, Reduviidae, Triatominae) in the municipality of Muaná, Marajó Island, State of Pará, Brazil (sic). Mem. Inst. Oswaldo Cruz 94: 399-400.
- Vivas, A.S., H. Barazarte & D. Molina de Fernandéz. 2001. Primer registro de Eratyrus mucronatus Stål, 1959 (Hemiptera: Reduviidae) en el ambiente domiciliário em Venezuela (sic). Entomotropica 16: 215-217.
Publication Dates
-
Publication in this collection
12 Sept 2005 -
Date of issue
June 2005
History
-
Accepted
04 Feb 2005 -
Received
18 Aug 2004