Resumos
Bemisia tabaci (Genn.) é uma das principais pragas do tomateiro no Brasil, provocando severas perdas devido à sucção de seiva, indução do amadurecimento irregular dos frutos e transmissão de viroses. O objetivo deste trabalho foi determinar o efeito de tricomas glandulares no desenvolvimento de B. tabaci biótipo B em oito genótipos de tomateiro, em casa de vegetação. Utilizou-se o delineamento de blocos ao acaso, em arranjo fatorial (genótipo x exsudato), com três repetições, sendo os genótipos avaliados na condição com e sem exsudato. Adultos da mosca-branca foram transferidos para uma gaiola de voil, onde permaneceram durante 24h. As variáveis avaliadas foram número de ovos e viabilidade, período de incubação, número de ninfas, duração e sobrevivência da fase ninfal. Efetuou-se a contagem de tricomas e a determinação do pH e teor de acilglicose para todos os genótipos. Na presença de exsudato, LA716 proporcionou redução na oviposição do inseto, o que sugere efeito antixenótico. Resistência antixenótica em LA1739 e PI134417 não foi associada ao exsudato glandular. O efeito do exsudato na oviposição do inseto foi explicado pelo teor de acilglicose. O pH apresentou papel secundário na oviposição da mosca-branca. A densidade de tricomas favoreceu o desenvolvimento do inseto. LA1584 destacou-se pela redução na sobrevivência do inseto e prolongamento do ciclo, além de propiciar oviposição intermediária, podendo ser adequado para incorporação em programas de melhoramento. Entretanto, há necessidade de estudos adicionais para determinar o potencial de uso desse genótipo ou de seus híbridos no manejo integrado de pragas.
Lycopersicon; mosca-branca; Bemisia argentifolii
Bemisia tabaci (Genn.) is one of the most important tomato pests in Brazil and causes severe losses due to the sap suction, the induction of the irregular ripening of the fruit and virus transmission. The objective of this study was to determine the effect of exudates released by the glandular trichomes on the development of B. tabaci biotype B on eight tomato genotypes, under greenhouse conditions. The experiment was arranged as a factorial scheme (genotype x exudate) in a randomized block design, with three replications, being the genotypes evaluated in two conditions (with and without exudate). Whitefly adults were transferred to a voile cage for 24h. The variables evaluated were number of eggs and viability, incubation period, number of nymphs, duration and viability of the nymphal stage. Trichome counts and determination of pH and acylglucose content were made for all the genotypes. Intact leaflets of LA716 were less accepted for oviposition than leaflets without exudate, which suggests an antixenotic effect. Antixenotic resistance in LA1739 and PI134417 was not associated to the glandular exudate. The effect of exudates on the insect oviposition was explained by the acylglucose content. The pH showed secondary role on the whitefly oviposition. Trichomes density improved the insect development. LA1584 reduced the insect survival and increased its cycle, besides promoting an intermediate oviposition. For this reason, it can be useful in breeding programs. However, further studies are needed to determine the potential of this genotype and hybrids for future utilization in integrated pest management.
Lycopersicon; silverleaf whitefly; Bemisia argentifolii
CROP PROTECTION
Exsudato glandular de genótipos de tomateiro e desenvolvimento de Bemisia tabaci (Genn.) (Sternorryncha: Aleyrodidae) biótipo B
Glandular exudate of tomato genotypes and development of Bemisia tabaci (Genn.) (Sternorryncha: Aleyrodidae) biotype B
Marilene FancelliI; José D. VendramimII; Rosa T.S. FrighettoIII; André L. LourençãoIV
IEmbrapa Mandioca e Fruticultura, C. postal 007, 44380-000, Cruz das Almas, BA, fancelli@cnpmf.embrapa.br
IIDepto. Entomologia, Fitopatologia e Zoologia Agrícola, ESALQ/USP, C. postal 9, 13418-900, Piracicaba, SP
IIIEmbrapa Meio Ambiente, Rodovia SP 340, Campinas-Mogi Mirim, km 127,5, 13820-000, Jaguariúna, SP
IVInstituto Agronômico (IAC), C. postal 28, 13001-970, Campinas, SP
RESUMO
Bemisia tabaci (Genn.) é uma das principais pragas do tomateiro no Brasil, provocando severas perdas devido à sucção de seiva, indução do amadurecimento irregular dos frutos e transmissão de viroses. O objetivo deste trabalho foi determinar o efeito de tricomas glandulares no desenvolvimento de B. tabaci biótipo B em oito genótipos de tomateiro, em casa de vegetação. Utilizou-se o delineamento de blocos ao acaso, em arranjo fatorial (genótipo x exsudato), com três repetições, sendo os genótipos avaliados na condição com e sem exsudato. Adultos da mosca-branca foram transferidos para uma gaiola de voil, onde permaneceram durante 24h. As variáveis avaliadas foram número de ovos e viabilidade, período de incubação, número de ninfas, duração e sobrevivência da fase ninfal. Efetuou-se a contagem de tricomas e a determinação do pH e teor de acilglicose para todos os genótipos. Na presença de exsudato, LA716 proporcionou redução na oviposição do inseto, o que sugere efeito antixenótico. Resistência antixenótica em LA1739 e PI134417 não foi associada ao exsudato glandular. O efeito do exsudato na oviposição do inseto foi explicado pelo teor de acilglicose. O pH apresentou papel secundário na oviposição da mosca-branca. A densidade de tricomas favoreceu o desenvolvimento do inseto. LA1584 destacou-se pela redução na sobrevivência do inseto e prolongamento do ciclo, além de propiciar oviposição intermediária, podendo ser adequado para incorporação em programas de melhoramento. Entretanto, há necessidade de estudos adicionais para determinar o potencial de uso desse genótipo ou de seus híbridos no manejo integrado de pragas.
Palavras-chave: Lycopersicon, mosca-branca, Bemisia argentifolii
ABSTRACT
Bemisia tabaci (Genn.) is one of the most important tomato pests in Brazil and causes severe losses due to the sap suction, the induction of the irregular ripening of the fruit and virus transmission. The objective of this study was to determine the effect of exudates released by the glandular trichomes on the development of B. tabaci biotype B on eight tomato genotypes, under greenhouse conditions. The experiment was arranged as a factorial scheme (genotype x exudate) in a randomized block design, with three replications, being the genotypes evaluated in two conditions (with and without exudate). Whitefly adults were transferred to a voile cage for 24h. The variables evaluated were number of eggs and viability, incubation period, number of nymphs, duration and viability of the nymphal stage. Trichome counts and determination of pH and acylglucose content were made for all the genotypes. Intact leaflets of LA716 were less accepted for oviposition than leaflets without exudate, which suggests an antixenotic effect. Antixenotic resistance in LA1739 and PI134417 was not associated to the glandular exudate. The effect of exudates on the insect oviposition was explained by the acylglucose content. The pH showed secondary role on the whitefly oviposition. Trichomes density improved the insect development. LA1584 reduced the insect survival and increased its cycle, besides promoting an intermediate oviposition. For this reason, it can be useful in breeding programs. However, further studies are needed to determine the potential of this genotype and hybrids for future utilization in integrated pest management.
Key words:Lycopersicon, silverleaf whitefly, Bemisia argentifolii
Na cultura do tomate, além dos danos provocados pela alimentação de ninfas e de adultos, Bemisia tabaci (Genn.) biótipo B pode transmitir fitoviroses limitantes à produção, além de provocar a desordem fisiológica conhecida como amadurecimento irregular dos frutos, que deprecia o valor comercial do produto (Brown & Bird 1992).
Altamente polífago, o inseto pode ter seu desenvolvimento afetado pelo hospedeiro no qual se cria (Drost et al. 1998). Tsai & Wang (1996) avaliaram o desenvolvimento, sobrevivência e reprodução de B. tabaci biótipo B em berinjela, tomate, batata-doce, pepino e feijão. Morillo & Marcano (1997) constataram diferenças entre dez genótipos de tomateiro, com relação a todas as fases de B. tabaci, em condições de laboratório.
A resistência de plantas a insetos é uma das principais estratégias utilizadas no manejo integrado da praga. A antixenose é caracterizada pela menor utilização do hospedeiro pelo inseto, quer seja para alimentação, oviposição e/ou abrigo; a antibiose, pelos efeitos deletérios à sobrevivência e desenvolvimento do inseto e a tolerância, pela capacidade da planta de manter a produção sob ataque da praga (Lara 1991). Entre os tipos de resistência, a antixenose é a mais citada para o tomate. A presença de tricomas glandulares dos tipos IV e VI encontrados em Lycopersicon hirsutum, L. hirsutum f. glabratum e L. pennellii tem sido relatada como a causa da resistência a moscas-brancas (Channarayappa et al. 1992, Barten et al. 1994, Snyder et al. 1998). Muigai et al. (2003) verificaram uma correlação significativa e negativa entre a densidade de tricomas do tipo IV e a população de Bemisia argentifolii Bellows & Perring em acessos de Lycopersicon spp., sendo que os mais altos níveis de resistência foram observados em acessos de L. pennellii (LA716, LA1340 e LA2560).
Em estudo sobre a atratividade de genótipos de tomateiro sobre B. tabaci biótipo B, Fancelli et al. (2003) constataram que adultos que se deslocaram para LA716 (L. pennellii) foram capturados nos tricomas glandulares presentes nas folhas, não se registrando adultos vivos nos folíolos amostrados. Esse efeito foi também observado para alguns genótipos pertencentes a L. hirsutum (LA1739, LA1740 e PI127826) e L. hirtsutum f. glabratum (PI134417 e PI134418), porém em menor proporção, mas tal fato não impediu a oviposição e o desenvolvimento dos insetos. Em trabalho desenvolvido por Gentile et al. (1968), a remoção do exsudato glandular de LA716 possibilitou o desenvolvimento de Trialeurodes vaporariorum (Westwood), comprovando-se que, nesse genótipo, a resistência se deve ao efeito antixenótico exercido pelos exsudatos glandulares dos tricomas.
Assim, este trabalho foi conduzido com o objetivo de estudar o efeito do exsudato de tricomas glandulares de tomateiro no desenvolvimento de B. tabaci biótipo B em oito genótipos de Lycopersicon spp., em casa de vegetação.
Material e Métodos
O ensaio foi realizado em casa de vegetação com teto de plástico leitoso e paredes com tela anti-afídeo, com sistema de nebulização para redução da temperatura. A temperatura variou de 8ºC a 34ºC e a umidade de 40% a 100%.
Obtenção dos Tomateiros. Foram utilizados oito genótipos de tomateiro (Lycopersicon spp.), selecionados previamente (Fancelli et al. 2003) como altamente resistentes (LA716 - L. pennellii, LA1739 - L. hirsutum e PI134417 - L. hirsutum f. glabratum), moderadamente resistentes (LA462 - L. peruvianum, LA1584 - L. pimpinellifolium e Santa Clara - L. esculentum), suscetível (P25 - L. esculentum) e altamente suscetível (LA1609 - L. peruvianum). Efetuou-se a pré-germinação das sementes desses genótipos em condições de laboratório, mantendo-as em placa de Petri com alta saturação de umidade, por aproximadamente oito dias ou até o início da germinação. Posteriormente, as plântulas foram transferidas para caixa de germinação de isopor contendo substrato à base de vermiculita, apropriado para essa finalidade. Após o enraizamento, as mudas foram transplantadas em sacos plásticos com diâmetro de 15 cm e altura de 25 cm contendo mistura de solo, matéria orgânica e areia (uma planta/ saco plástico). O experimento foi instalado quando as plantas estavam com 36 dias de idade (6ª semana), na fase vegetativa (15 dias após o transplante). As plantas foram mantidas eretas com o auxílio de um tutor (vara de bambu), preso a cada planta por meio de fitilho plástico.
Criação Estoque de B. tabaci Biótipo B. Foi mantida uma criação estoque de B. tabaci biótipo B em condições de insetário, utilizando-se como hospedeiras plantas de soja e amendoim-bravo, renovadas semanalmente.
Infestação. Selecionaram-se folhas do 3º e 4º nós a partir do ápice, com remoção (ou não) do exsudato glandular de ambas as superfícies das folhas com álcool comercial a 70% e em seguida com água destilada. Utilizou-se o delineamento experimental de blocos ao acaso em esquema fatorial (presença e ausência de exsudato), com oito tratamentos (genótipos) e três repetições. A parcela foi constituída por três folíolos de cada genótipo. O confinamento dos insetos deu-se em gaiolas de voil (15 cm x 5 cm), fechadas com velcro (Souza & Vendramim 2000). Adultos da mosca-branca (50 casais) foram aspirados da criação estoque e introduzidos na gaiola, onde permaneceram durante 24h para oviposição. Os folíolos infestados, ao final do desenvolvimento das ninfas, foram novamente envolvidos pelas gaiolas de voil para confinar os adultos emergidos. As variáveis observadas foram: número e viabilidade de ovos, período de incubação, número de ninfas, duração e sobrevivência da fase ninfal (com base na presença de exúvias). Para contagem de ovos e ninfas, utilizou-se uma lupa manual com aumento de 20x. Os dados foram submetidos à análise de variância, sendo as médias comparadas pelo teste de Tukey (P < 0,05). Para número de ovos, ninfas e adultos emergidos, utilizou-se a transformação raiz (x + 1).
Contagem de Tricomas. A densidade foi determinada pela contagem de tricomas presentes em 1 mm2 na face adaxial e abaxial dos folíolos de tomateiro, sob microscópio estereoscópico, com 40x de aumento, conforme Snyder & Carter (1985). Não foi feita a identificação dos tipos de tricomas presentes. O número de repetições variou de acordo com a disponibilidade de folíolos em cada genótipo.
Convencionou-se adotar a denominação para os tratamentos como "folíolos com e sem exsudato", com base em materiais como LA716, PI134417 e LA1739. No entanto, essa terminologia pode não ser a mais adequada, em razão das diferenças entre as proporções de tricomas glandulares e não-glandulares em genótipos de tomateiro e também pelo fato de que, pelo método utilizado, não se assegure que somente o exsudato foi removido.
Determinação do pH dos Folíolos. O pH dos diferentes genótipos de tomateiro foi determinado com base em uma amostra de 2 g de massa fresca de folhas de cada genótipo, as quais foram maceradas em 5 ml de água (pH = 7,0).
Estimativa de Açúcares no Exsudato. Foram utilizadas folhas do 3º e 4º nós a partir do ápice de plantas com idades de seis e 14 semanas (idades correspondentes àquelas das plantas no início e final do experimento). Os ésteres de açúcar epicuticulares foram extraídos com diclorometano (10 ml) mergulhando-se de 3-4 folhas frescas neste solvente por cerca de 15 segundos e, em seguida, em água destilada. O excesso de água foi removido com papel-toalha e a área da superfície foliar foi medida em integrador portátil de área foliar LI-COR (LAI3000A), para estimar a quantidade de açúcar acumulada por unidade de área foliar. A solução de diclorometano foi evaporada e o resíduo foi redissolvido em 1,0 ml de metanol, logo antes das determinações de açúcares por colorimetria. O extrato em metanol (200 µl) foi hidrolisado com solução aquosa 0,04N de NaOH (200 µl), a 55ºC por 1h. A solução foi resfriada e o tubo acondicionado em gelo-água, em seguida foi adicionado 3 ml de reagente de antrona, misturando-se bem por alguns minutos, e retirado do gelo para permitir que atingisse a temperatura ambiente. A leitura foi feita em espectrofotômetro (Perkin Elmer, mod. Lambda 20) a 625 nm. O reagente de antrona foi preparado adicionando-se 0,15 g de antrona desidratada a 50 ml de ácido sulfúrico (adicionar 45 ml de ácido concentrado a 5 ml de água, com o frasco acondicionado em gelo visto que a adição de ácido gera calor), deixar repousar por cerca de 30 min para clarificar. Este reagente deve ser preparado e usado dentro de 12h após a preparação (Passos 1996). A curva de calibração foi preparada com concentrações conhecidas de glicose por meio da diluição de solução-estoque de glicose, nas concentrações de 0; 7,5; 15; 30; 75; 150 mg.ml-1 em água. O branco continha 0,2 ml de base e 0,2 ml de água, além da solução de antrona. Nessas condições, o valor de correlação positiva encontrado foi de 0,997. Os resultados referem-se às médias de três repetições por amostra.
Resultados
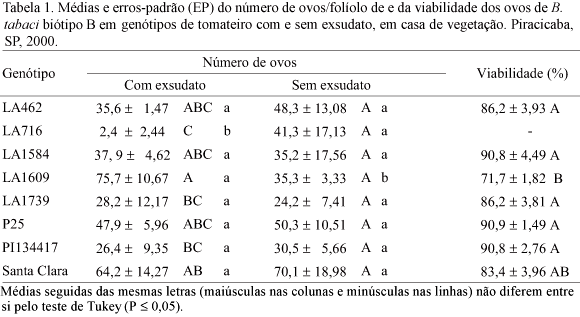
O número de ovos colocados por B. tabaci biótipo B foi afetado pelos genótipos de tomateiro nos quais os adultos foram confinados, sendo ainda significativa a interação dos fatores genótipo e exsudato (gl = 7, F = 2,83, P = 0,02) (Tabela 1). Em folíolos com exsudato, o número de ovos em LA716 foi inferior àqueles registrados em LA1609 e Santa Clara. Na ausência do exsudato, não foram constatadas diferenças significativas entre os genótipos (Tabela 1).
Para folíolos com exsudato, o número de ovos decresceu com o aumento do teor de acilglicose na 14ª semana, ajustando-se à equação y = 53,742e-0,0013x, com R2 de 0,93 (Fig. 1A).
Considerando o número de ovos e pH de folíolos de genótipos de tomateiro, observou-se que a oviposição foi estimulada pelo aumento do pH, apesar dos genótipos LA1739 e PI134417 não se adequaram à curva, visto que reduziram a oviposição do inseto, mesmo apresentando valores de pH relativamente elevados (5,97 e 5,87, respectivamente). A relação entre o número de ovos e o pH de folíolos de tomateiro foi convenientemente descrita pelo ajuste de uma equação linear aos dados (y = 83,16x 430,07, R2 = 0,80) (Fig. 1B).
Não foram observadas diferenças significativas entre os tratamentos em relação à duração do período de incubação (gl = 6, F = 1,53, P = 0,21). O exsudato, bem como a interação dos fatores genótipo e exsudato, também não influenciaram na duração dessa fase. O período de incubação médio foi de 11,3 ± 0,04 dias, com variação de 11,0 a 11,5 dias.
A viabilidade dos ovos foi afetada significativamente apenas pelos genótipos de tomateiro (gl = 6, F = 5,09, P = 0,002) (Tabela 1). Considerando-se as médias obtidas em folíolos com e sem exsudato, registrou-se o menor valor em LA1609, diferindo das médias observadas para todos os genótipos, com exceção de Santa Clara.
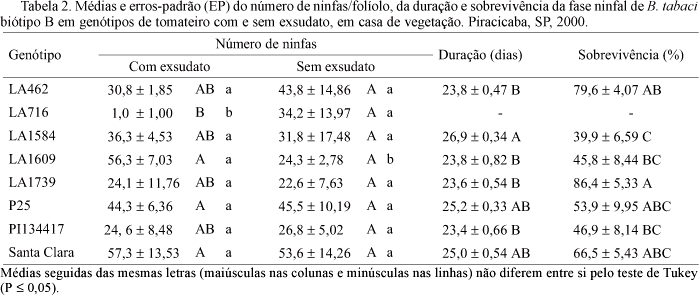
Verificou-se efeito significativo da interação dos fatores genótipo e exsudato quanto ao número de ninfas (gl = 7, F = 2,67, P = 0,03) (Tabela 2). O efeito dos genótipos foi constatado apenas em folíolos com exsudato. A presença do exsudato em folíolos de LA716 influenciou negativamente essa variável; entretanto, para LA1609, o maior número de ninfas foi registrado na condição com exsudato (Tabela 2).
A duração da fase ninfal foi afetada significativamente pela presença do exsudato (gl = 1; F = 8,42, P = 0,01), não sendo significativa a interação dos fatores genótipo e exsudato (gl = 6, F = 1,65, P = 0,17) (Tabela 2). Foi observada maior duração dessa fase em folíolos com exsudato (25,1 dias) do que naqueles sem exsudato (24,0 dias). A duração da fase ninfal em LA1584 foi significativamente superior às dos demais genótipos, excetuando P25 e Santa Clara.
Em relação ao período ninfal e densidade de tricomas, observou-se que a duração da fase foi encurtada em função do aumento do número de tricomas/mm2, seguindo uma tendência de ajuste logarítmico aos dados [y = -1,0231Ln(x) + 28,759, R2 = 0,72] (Fig 1C).
Quanto à sobrevivência ninfal, registrou-se efeito significativo apenas dos genótipos (gl = 6, F = 5,48, P = 0,001), sendo o valor verificado em LA1584 inferior aos observados em LA1739 e LA462 (Tabela 2).
Não houve efeito significativo da interação dos fatores genótipo e exsudato (presença/ausência) sobre a emergência do inseto (gl = 7, F = 1,31, P = 0,28) (Tabela 3). O número de adultos emergidos de B. tabaci biótipo B foi afetado apenas pelos genótipos de tomateiro (gl = 7, F = 3,51, P = 0,007). Desse modo, considerando-se a média entre os folíolos com e sem exsudato, o número de adultos foi significativamente inferior em LA716, PI134417 e LA1584 do que em Santa Clara.
A duração da fase pré-imaginal, à semelhança do que ocorreu para a fase ninfal, foi afetada significativamente pelos genótipos (gl = 6, F = 5,96, P = 0,00073) e pelo exsudato (gl = 1; F = 8,90; P = 0,06). Não foi significativa a interação genótipo e exsudato (gl = 6, F = 1,55, P = 0,20) (Tabela 3). O genótipo LA1584 promoveu maior duração para essa fase, sendo esse valor superior àqueles observados nos demais genótipos, com exceção de Santa Clara e P25. Folíolos com exsudato promoveram maior duração dessa fase (36,4 dias) do que folíolos sem exsudato (35,2 dias).
Com relação à sobrevivência da fase pré-imaginal, verificou-se efeito significativo apenas dos genótipos (gl = 6, F = 5,38, P = 0,0013) (Tabela 3). A sobrevivência dessa fase foi significativamente mais baixa em LA1609 e LA1584 do que em LA1739 e LA462.
Para sobrevivência pré-imaginal e densidade de tricomas, constatou-se uma tendência de aumento na sobrevivência do inseto em função da elevação do número de tricomas/mm2, sendo o ajuste explicado pela equação y = 13,692x0,3404 (R2 = 0,69) (Fig. 1D).
Discussão
A possibilidade de discriminação dos genótipos quando se avaliou a população do inseto e a sobrevivência corrobora com os dados observados para T. vaporariorum (Bas et al. 1992). Entretanto, com relação à duração do período de desenvolvimento, os resultados obtidos discordam, em parte, de Romanow et al. (1991) e Bas et al. (1992). Nesses dois trabalhos, o período de desenvolvimento não foi considerado adequado à constatação de antibiose, por serem pequenas as diferenças entre genótipos com relação a essa variável.
LA1739 e PI134417 foram materiais que se destacaram pela antixenose para oviposição, em condição de livre escolha (Fancelli et al. 2003). No presente trabalho, esse comportamento repetiu-se, mas com menor intensidade, em folíolos com exsudato (Tabela 1), evidenciando que fatores relacionados à resistência não se manifestam da mesma forma em experimentos com livre escolha e confinamento, conforme discutiram Romanow et al. (1991). No presente trabalho, a remoção do exsudato em LA1739 e PI134417 não promoveu alteração no número de ovos (Tabela 1), indicando que outros fatores podem estar relacionados à antixenose para oviposição. Dentre esses fatores, podem ser citados o fotoperíodo (Snyder et al. 1998) e a idade das plantas (Bas et al. (1992). Assim, como o número de ovos foi semelhante ao registrado em LA716, admite-se a ocorrência de antixenose em LA1739 e PI134417, sem, contudo descartar a antibiose em PI134417, pelo efeito negativo desse genótipo na sobrevivência ninfal (Tabela 2). À semelhança do que foi verificado por Romanow et al. (1991), parece que o encurtamento da fase ninfal observado para L. hirsutum f. glabratum é devido ao pequeno número de indivíduos que conseguem superar essa fase. Em LA1739, como a viabilidade de ovos e ninfas foi elevada e a duração da fase ninfal encurtada, admite-se que representa um excelente substrato nutricional para o inseto, desde que a barreira à oviposição seja vencida.
Quanto a LA1584, o número de ovos registrado para folíolos com exsudato (Tabela 1) foi intermediário, denotando certo grau de antixenose para oviposição. Registrou-se baixa sobrevivência ninfal e pré-imaginal nesse genótipo (Tabelas 2 e 3). Uma grande proporção das ninfas mortas encontrava-se nos ínstares iniciais do desenvolvimento, ao contrário do que foi observado nos demais genótipos. Pode-se, desse modo, admitir a ocorrência de antibiose, embora não possa ser descartada a antixenose, mediada por estímulos químicos, físicos ou morfológicos (Lara 1991). Verificou-se, também, o prolongamento da fase ninfal nos folíolos de LA1584 (Tabela 2), o que poderia ser uma resposta à pior qualidade desse material para alimentação do inseto, reforçando a hipótese de antibiose.
Em LA716, a redução na oviposição caracteriza um forte efeito antixenótico. Não foi possível verificar o efeito do genótipo no desenvolvimento do inseto na condição com exsudato, em razão do pequeno número de ovos e, conseqüentemente, de ninfas. O número de ovos aumentou após a remoção do exsudato glandular, indicando uma relação entre o exsudato e a oviposição no genótipo. Com base na literatura, o mecanismo de resistência nessa espécie está ligado à antixenose para oviposição, mediada por acilaçúcares presentes no exsudato glandular de tricomas tipo IV (Barten et al. 1994, Nombela et al. 2000, Muigai et al. 2003). No presente trabalho, o genótipo LA716 acumulou a maior concentração de acilglicose na superfície foliar dentre os genótipos avaliados, resultando na resposta de redução no número de ovos (Fig. 1A) para folíolos com exsudato.
Em função da oviposição intermediária em LA462 (com exsudato) (Tabela 1), admite-se preferencialmente a ocorrência de antixenose, pois o material promoveu o encurtamento da fase ninfal, associado a uma das mais altas sobrevivências nessa fase e na de ovo. LA1609, selecionado como um dos padrões suscetíveis, com base nas altas densidades populacionais de adultos (Fancelli et al. 2003), foi preferido para oviposição, na condição com exsudato. Esse comportamento pode estar associado ao pH elevado encontrado em folíolos desse genótipo, conforme discutido por Berlinger et al. (1983), para B. tabaci em folhas de algodão, porém não exclusivamente determinado por esse fator. É interessante verificar que LA1609 é relativamente pouco pubescente, o que poderia ser um fator desfavorável à seleção hospedeira. Apesar da preferência para oviposição, o genótipo interferiu negativamente na sobrevivência de ovos e ninfas, o que denota a ocorrência de antibiose. A mortalidade foi verificada em estágios mais avançados do desenvolvimento ninfal, sendo detectadas, em muitos casos, ninfas infectadas pelo fungo oportunista do gênero Cladosporium. Apesar de promover o encurtamento da fase ninfal, esse efeito pode estar relacionado ao reduzido número de indivíduos que conseguem suplantar essa fase. Quando o exsudato foi removido, o material foi menos adequado à oviposição, indicando efeito negativo do processo de remoção para o genótipo.
Quanto aos genótipos pertencentes a L. esculentum, registrou-se acentuada preferência para oviposição em Santa Clara, na condição com exsudato, verificando-se que P25 apresentou maior grau de antixenose para oviposição do que Santa Clara. Um certo grau de antibiose foi denotado pela redução na viabilidade dos ovos em Santa Clara e alongamento e redução da sobrevivência da fase ninfal para ambos os genótipos.
O efeito do exsudato na seleção hospedeira para oviposição pôde ser explicado pelos teores de acilglicose, corroborando com dados da literatura (Mutschler et al. 1996). O pH dos folíolos apresentou papel secundário na oviposição da mosca-branca, assumindo importância após a seleção dos genótipos com base nos teores de acilglicose. Independente do tipo de tricoma presente na face abaxial de folíolos de tomateiro, a densidade de tricomas favoreceu o desenvolvimento do inseto, reduzindo a duração da fase ninfal e aumentando a sobrevivência da fase pré-imaginal. A presença de exsudato determinou o prolongamento da fase ninfal e pré-imaginal do inseto, além de interagir com os genótipos para as variáveis número de ovos e ninfas. LA1584 destacou-se pela redução na sobrevivência do inseto e prolongamento do ciclo, além de propiciar oviposição intermediária, característica interessante para utilização em programas de manejo integrado de pragas. Entretanto, há necessidade de estudos adicionais para determinar o efeito desse genótipo sobre inimigos naturais da praga.
Agradecimentos
À Universidade da Califórnia e Embrapa Hortaliças pelo fornecimento das sementes de parte dos genótipos de tomateiro utilizados e à bióloga Solange Aparecida Vieira, pela identificação do fungo Cladosporium sp.
Literatura Citada
Received 12/VIII/04. Accepted 22/III/05.
- Barten, J.H.M., C.H. Thome, M.R. Stevens, D.J. Schuster, J.W. Scott & O.L. Chambliss. 1994. Evaluating resistance in tomato to the silverleaf whitefly, Bemisia argentifolii Phytoparasitica 22: 330-331.
- Bas, N., C. Mollema & P. Lindhout. 1992. Resistance in Lycopersicon hirsutum f. glabratum to the greenhouse whitefly (Trialeurodes vaporariorum) increases with plant age. Euphytica 64: 189-195.
- Berlinger, M.J., Z. Magal & A. Benzioni. 1983. The importance of pH in food selection by the tobacco whitefly, Bemisia tabaci Phytoparasitica 11: 151-160.
- Brown, J.K. & J. Bird. 1992. Whitefly-transmitted geminiviruses and associated disorders in the Americas and the Caribbean Basin. Plant Dis. 76: 220-225.
- Channarayappa, S., G., V. Muniyappa & R.H. Frist. 1992. Resistance of Lycopersicon species to Bemisia tabaci, a tomato leaf curl virus vector. Can. J. Bot. 70: 2184-2192.
- Drost, Y.C., J.C. van Lenteren & H.J.W. van Roermund. 1998. Life-history parameters of different biotypes of Bemisia tabaci (Hemiptera: Aleyrodidae) in relation to temperature and host plant: A selective review. Bull. Entomol. Res. 88: 219-229.
- Fancelli, M., J.D. Vendramim, A.L. Lourenção & C.T S. Dias. 2003. Atratividade e preferência para oviposição de Bemisia tabaci (Gennadius) (Hemiptera: Aleyrodidae) biótipo B em genótipos de tomateiro. Neotrop. Entomol. 32: 319-328.
- Gentile, A.G., R.E. Webb & A.K. Stoner. 1968. Resistance in Lycopersicon and Solanum to greenhouse whiteflies. J. Econ. Entomol. 61: 1355-1357.
- Lara, F.M. 1991. Princípios de resistência de plantas a insetos. 2 ed., São Paulo, Ed. Ícone, 336p.
- Morillo, F.E. & R.V. Marcano B. 1997. Estudio del desarrollo de la mosca blanca en diferentes genotipos de tomate. Agron. Trop. 47: 271-286.
- Muigai, S.G., M.J.Bassett, D.J. Schuster & J.W. Scott. 2003. Greenhouse and field screening of wild Lycopersicon germplasm for resistance to the whitefly Bemisia argentifolii Phytoparasitica 31: 27-38.
- Mutschler, M.A., R.W. Doerge, S.C. Liu, J.P. Kuai, B.E. Liedl & J.A. Shapiro. 1996. QTL analysis of pest resistance in the wild tomato Lycopersicon pennellii: QTLs controlling acylsugar level and composition. Theor. Appl. Gen. 92: 709-718.
- Nombela, G., F. Beitia & M. Muñiz. 2000. Variation in tomato host response to Bemisia tabaci (Hemiptera: Aleyrodidae) in relation to acyl sugar content and presence of the nematode and potato aphid resistance gene Mi. Bull. Entomol. Res. 90: 161-167.
- Passos, L.P. 1996. Carboidratos Análise quantitativa. In L.P. Passos (ed.), Métodos analíticos e laboratoriais em fisiologia vegetal. Embrapa-CNPGL, 1996, Coronel Pacheco, 223p.
- Romanow, L.R., O.M.B. de Ponti & C. Mollema. 1991. Resistance in tomato to the greenhouse whitefly: Analysis of population dynamics. Entomol. Exp. Appl. 60: 247-259.
- Simmons, A.M. 1994. Oviposition on vegetables by Bemisia tabaci (Homoptera: Aleyrodidae): Temporal and leaf surface factors. Environ. Entomol. 23: 381-389.
- Snyder, J.C., A.M. Simmons & R.R. Thacker. 1998. Attractancy and ovipositional response of adult Bemisia argentifolii (Homoptera: Aleyrodidae) to type IV trichome density on leaves of Lycopersicon hirsutum grown in three day-length regimes. J. Entomol. Sci. 33: 270-281.
- Snyder, J.C. & C.D. Carter. 1985. Trichomes on leaves of Lycopersicon hirsutum, L. esculentum and their hybrids. Euphytica 34: 53-64.
- Souza, A.P. & J.D. Vendramim. 2000. Atividade ovicida de extratos aquosos de meliáceas sobre a mosca branca Bemisia tabaci (Gennadius) biótipo B em tomateiro. Sci. Agric. 57: 403-406.
- Tsai, H. & K. Wang 1996. Development and reproduction of Bemisia argentifolii (Homoptera: Aleyrodidae) on five host plants. Environ. Entomol. 25: 810-816.
Datas de Publicação
-
Publicação nesta coleção
26 Set 2005 -
Data do Fascículo
Ago 2005
Histórico
-
Recebido
12 Ago 2004 -
Aceito
22 Mar 2005