Abstracts
Canopy arthropods, mainly from palm trees, are little known in the Brazilian Cerrado. In order to describe the arthropod community structure associated with the crown of Mauritia flexuosa (Arecaceae), we sampled 150 palm trees in six "veredas" of the Federal District, Brazil, in wild, rural and periurban areas in the rainy season. The arthropods within abandoned bird nests, mammal refuges, leaves and organic matter were manually collected, preserved in ethanol 70% and separated by order, family, morphospecies and feeding guilds. Stem height and diameter of the palm crowns were measured and leaves and bird nests were counted. We collected 3,862 arthropods, from 15 orders, 45 families and 135 morphospecies. The most abundant orders were Coleoptera (28.6%), Blattodea (21.8%), Collembola (11.4%) and Hemiptera (10.2%). The families Blaberidae, Entomobryidae, Reduviidae, Oniscidae, Staphylinidae, Carabidae and Formicidae, represented 82.1% of all individuals collected. The majority of morphospecies was not abundant, 71 (52.6%) were represented by less than 1 individual/tree. Coleopterans accounted for the highest number of morphospecies (43.7%) followed by Araneae (20.0%). The analysis of the arthropod feeding guilds showed prevalence of predatory/hematophagous ones (36.0%). Arthropod richness and abundance presented smaller values for periurban environment. The number of bird nests presented positive correlation with abundance and richness; this was not found when considering the measurements of the palm trees. The importance of M. flexuosa for the maintenance of the arthropod fauna of the "veredas" in Cerrado biome is discussed.
Arthropoda; community structure; buriti; cerrado; species richness
Artrópodos associados à copa de árvores, principalmente palmeiras, são pouco conhecidos no Cerrado. Para descrever a estrutura da comunidade de artrópodos associada à copa de Mauritia flexuosa (Arecaceae) foram amostradas 150 palmeiras em seis "veredas" do Distrito Federal, de áreas silvestres, rurais e periurbanas na estação chuvosa. Os artrópodos presentes nos ninhos abandonados de aves, refúgios de mamíferos, folhas e matéria orgânica foram coletados manualmente, fixados em etanol 70% e separados em ordem, família, morfoespécie e guildas alimentares. As características das palmeiras medidas foram altura da estipe, diâmetro da copa, número de folhas e de ninhos de aves nas palmeiras. Foram coletados 3.862 indivíduos, pertencentes a 15 ordens, 45 famílias e 135 morfoespécies. As ordens mais abundantes foram Coleoptera (28,6%), Blattodea (21,8%), Collembola (11,4%) e Hemiptera (10,2%). As famílias Blaberidae, Tenebrionidae, Entomobryidae, Reduviidae, Oniscidae, Staphylinidae, Carabidae e Formicidae representaram 82,1% de todos os indivíduos coletados. A maioria das morfoespécies foi pouco abundante, 71 (52,6%) apresentaram uma abundância média igual ou menor que 1 indivíduo/palmeira. Coleoptera compreendeu o maior número de morfoespécies (43,7%) seguida de Araneae (20,0%). A análise das guildas alimentares mostrou prevalência de predadores e hematófagos (36,0%). A riqueza e a abundância de artrópodos foram menores no ambiente periurbano. O número de ninhos de aves apresentou correlação positiva com abundância e riqueza, o que não ocorreu com as medidas das palmeiras. A importância de M. flexuosa para a manutenção da artropodofauna nas "veredas" no bioma Cerrado é discutida.
Arthropoda; estrutura da comunidade; buriti; cerrado; riqueza de espécies
ECOLOGY, BEHAVIOR AND BIONOMICS
Arthropods associated with the crown of Mauritia flexuosa (Arecaceae) palm trees in three different environments from Brazilian Cerrado
Artrópodos associados à copa da palmeira Mauritia flexuosa (Arecaceae) em três ambientes diferentes no Cerrado brasileiro
Rodrigo Gurgel-GonçalvesI, II; Alexandre R.T. PalmaI; Paulo C. MottaIII; Maria E. BarIV; Cesar A.C. CubaII
ILab. Zoologia, Curso de Ciências Biológicas, Univ. Católica de Brasília, Aguas Claras, QS 7, lote 1, sala M331, 71966-700, Brasília, DF, Brasil
IILab. Parasitologia Médica e Biologia de Vetores, Área de Patologia, Faculdade de Medicina
IIILab. Aracnídeos, Depto. Zoologia, Instituto de Biologia, Univ. Brasília, Brasília, Asa Norte, 70910-90, Brasília, DF, Brasil
IVLab Artrópodos, Facultad de Ciencias Exactas y Naturales, Univ. Nacional del Nordeste, Corrientes, Argentina
ABSTRACT
Canopy arthropods, mainly from palm trees, are little known in the Brazilian Cerrado. In order to describe the arthropod community structure associated with the crown of Mauritia flexuosa (Arecaceae), we sampled 150 palm trees in six "veredas" of the Federal District, Brazil, in wild, rural and periurban areas in the rainy season. The arthropods within abandoned bird nests, mammal refuges, leaves and organic matter were manually collected, preserved in ethanol 70% and separated by order, family, morphospecies and feeding guilds. Stem height and diameter of the palm crowns were measured and leaves and bird nests were counted. We collected 3,862 arthropods, from 15 orders, 45 families and 135 morphospecies. The most abundant orders were Coleoptera (28.6%), Blattodea (21.8%), Collembola (11.4%) and Hemiptera (10.2%). The families Blaberidae, Entomobryidae, Reduviidae, Oniscidae, Staphylinidae, Carabidae and Formicidae, represented 82.1% of all individuals collected. The majority of morphospecies was not abundant, 71 (52.6%) were represented by less than 1 individual/tree. Coleopterans accounted for the highest number of morphospecies (43.7%) followed by Araneae (20.0%). The analysis of the arthropod feeding guilds showed prevalence of predatory/hematophagous ones (36.0%). Arthropod richness and abundance presented smaller values for periurban environment. The number of bird nests presented positive correlation with abundance and richness; this was not found when considering the measurements of the palm trees. The importance of M. flexuosa for the maintenance of the arthropod fauna of the "veredas" in Cerrado biome is discussed.
Key words: Arthropoda, community structure, buriti, cerrado, species richness
RESUMO
Artrópodos associados à copa de árvores, principalmente palmeiras, são pouco conhecidos no Cerrado. Para descrever a estrutura da comunidade de artrópodos associada à copa de Mauritia flexuosa (Arecaceae) foram amostradas 150 palmeiras em seis "veredas" do Distrito Federal, de áreas silvestres, rurais e periurbanas na estação chuvosa. Os artrópodos presentes nos ninhos abandonados de aves, refúgios de mamíferos, folhas e matéria orgânica foram coletados manualmente, fixados em etanol 70% e separados em ordem, família, morfoespécie e guildas alimentares. As características das palmeiras medidas foram altura da estipe, diâmetro da copa, número de folhas e de ninhos de aves nas palmeiras. Foram coletados 3.862 indivíduos, pertencentes a 15 ordens, 45 famílias e 135 morfoespécies. As ordens mais abundantes foram Coleoptera (28,6%), Blattodea (21,8%), Collembola (11,4%) e Hemiptera (10,2%). As famílias Blaberidae, Tenebrionidae, Entomobryidae, Reduviidae, Oniscidae, Staphylinidae, Carabidae e Formicidae representaram 82,1% de todos os indivíduos coletados. A maioria das morfoespécies foi pouco abundante, 71 (52,6%) apresentaram uma abundância média igual ou menor que 1 indivíduo/palmeira. Coleoptera compreendeu o maior número de morfoespécies (43,7%) seguida de Araneae (20,0%). A análise das guildas alimentares mostrou prevalência de predadores e hematófagos (36,0%). A riqueza e a abundância de artrópodos foram menores no ambiente periurbano. O número de ninhos de aves apresentou correlação positiva com abundância e riqueza, o que não ocorreu com as medidas das palmeiras. A importância de M. flexuosa para a manutenção da artropodofauna nas "veredas" no bioma Cerrado é discutida.
Palavras-chave: Arthropoda, estrutura da comunidade, buriti, cerrado, riqueza de espécies
Studies on arthropods associated with the tropical canopies are concentrated in the Amazonian region (Erwin 1983, Adis et al. 1984, Hurtado Guerreiro et al. 2003) and Pantanal (Marques et al. 2001, Santos et al. 2003, Battirola et al. 2005) using the spraying method with biodegradable insecticides (canopy fogging). Specifically in relation to the arthropod fauna associated with palm trees, most of the studies are linked to species-plague, pollinators (reviewed by Santos et al. 2003) and vectors of tropical diseases, as the Chagas disease (Barretto et al. 1969, Carcavallo et al. 1998, Gurgel-Gonçalves et al. 2004). However, studies on the arthropod community structure in the crown of palm trees are scarce.
The Brazilian Cerrado is one of the largest biomes of the Americas and exhibits a great variety of ecosystems that allows the maintenance of a high faunal diversity (Oliveira & Marquis 2002). However, this biome has suffered significant human impacts during the past decades due to the rapid expansion of agricultural frontiers in Brazil (Klink & Moreira 2002, Jepson 2005). For this reason, Cerrado is considered as a "hot spot" with extreme priority for conservation (Klink & Machado 2005). Invertebrate fauna, especially arthropods, has been little studied when compared with vertebrates in Cerrado. However, some studies of insect diversity and ecology are known in Cerrado for Lepidoptera (Camargo & Becker 1999, Diniz et al. 2001, Motta 2002), Coleoptera (Pinheiro et al. 1998, Silva 2001, Milhomem 2003) and Hymenoptera (Diniz & Raw 1982, Barros et al. 1992, Silvestre & Brandão 2001).
There is a wide distribution of palm trees of the species Mauritia flexuosa L. (Arecaceae), known as "buriti", in the Cerrado biome, characterizing a typical ecosystem of central Brazil, the "vereda". In that phytophysiognomy, M. flexuosa and other groups of arboreal-herbaceous species develop in clean fields with a very humid soil (Ribeiro & Walter 1998, Oliveira & Marquis 2002). M. flexuosa can be found in forests borders, being a palm tree with a wide distribution in South America (Brazil, Colombia, Venezuela, Guyana, Ecuador, Peru and Bolivia). Palm height may achieve 25 m. Palmate leaves may be as long as 3-4 m and inflorescences may weigh up to 40 kg (Henderson et al. 1995). In the Federal District of Brazil (hereafter DF), M. flexuosa is thoroughly distributed in humid environments of rural areas, periurban environments and ecological reserves. Several birds and mammals species make their nests in M. flexuosa and use it as a temporary shelter and as a food resource, mainly in the dry season, when most of the fruits and seeds are scarce in the Cerrado (Prada 1994). There is a lack of knowledge regarding other organisms associated to that palm tree species, except for some works about diversity of fungi (Chaves 1998) and Triatominae (Hemiptera: Reduviidae) (Barretto et al. 1969, Carcavallo et al. 1998, Gurgel-Gonçalves et al. 2003, 2004). The present study aims to describe the arthropod community structure associated with M. flexuosa, to analyze the diversity patterns and to relate abundance and morphospecies richness with characteristics of the palm trees in different environments (ecological reserves, rural areas and periruban environments) in the rainy season.
Material and Methods
The DF (15°30'-16°03' S and 47°25'-48°12' W) occupies the center of Brazil in one of the highest areas of the region, the Central Plateau. Altitude is approximately 1,000 m and mean annual rainfall of 1,545 mm. A marked dry season (precipitation < 100 mm) extends from from May to September and mean annual temperatures range from 20°C to 21°C.
Six "veredas" were sampled in wet season in environments with different degrees of disturbance, including ecological reserves (wild environments) such as the Estação Ecológica de Águas Emendadas (15º34'27'' S, 47º36'28'' W), and the Reserva Ecológica do IBGE (15º55' 54"S, 47º54'02" W), rural areas as the Colônia Agrícola Rajadinha (15º46'14'' S, 47º38'58'' W) and Alphaville (16º04'05'' S, 47º32'39'' W), besides habitational areas (periurban environments) as the Setor de Mansões Park Way (15º54'12'' S, 47º56'51'' W) and Samambaia (15º54'12'' S, 48º09'02'' W). The classification of the environments occurred in accordance to the adjacent area to each sampling location; native vegetation (gallery forests and Cerrado strictu sensu) for wild environment; plantations and grasslands for rural environment and habitational areas for periurban environment. Other criteria such as the access conditions, proprietor permission (private areas) or the government permission (public areas) were also observed in the selection of the areas.
The total of 25 palm trees were sampled in each "vereda" using the method described by Gurgel-Gonçalves et al. (2004). Stem height and diameter of the crown were measured and number of leaves and bird nests were counted. The palm trees were climbed and the material present in the crown (organic matter, leaves and abandoned bird nests) was collected in plastic bags (Gurgel-Gonçalves et al. 2003, 2004). The bags were taken to the laboratory, where the material was selected, sorting the arthropods by order and development stage (adult and immature) and later by family and by morphospecies, according to Oliver & Beattie (1996). The families were separated in categories of feeding guilds in agreement with Moran & Southwood (1982), and those categories were sorted in three wider guilds: predatory/hematophagous, herbivorous/fungivorous and detritivorous/saprophagous.
To describe the arthropod community structure we used morphospecies composition and richness, relative abundance, diversity index (Shannon-Weaver), similarity (Morisita-Horn) and estimates of richness of arthropod morphospecies, through three models: Abundance-base Coverage Estimator (ACE), Chao model (Chao), and Model of Jacknife of Second Order (Jack). Those models count the rare species that could be captured in the collection, being obtained in the program of diversity analysis Estimates (Colwell 1997). The correlation among morphospecies richness, abundance of arthropods, tree height, diameter of the cup, the number of leaves and the number of bird nests in M. flexuosa palm trees were tested for significance using Spearman correlation coefficient (Zar 1996).
Results and Discussion
In 150 sampled palm trees we collected 3,862 individuals of Arthropoda (Insecta, Arachnida and Crustacea) distributed in 15 orders, 45 families and 135 morphospecies (Table 1). The most abundant orders were Coleoptera (28.6%), Blattodea (21.8%), Collembola (11.4%), Hemiptera (10.2%), Isopoda (7.3%), Araneae (6.6%) and Hymenoptera (6.2%). Among the identified families, the most abundant were Blaberidae (Blattodea) (21.6%), Tenebrionidae (Coleoptera) (13,0%), Entomobryidae (Collembola) (11.4%), Reduviidae (Hemiptera) (9.1%), Oniscidae (Isopoda) (7.3%), Staphylinidae (Coleoptera) (6,9%), Carabidae (Coleoptera) (6,7%) and Formicidae (Hymenoptera) (6.1%), representing 82.1% of all individuals. Blaberidae was also the most frequent family, occurring in 35.0% of the sampled palm trees. The majority of the families (77.0%) presented a frequency inferior to 10.0%, in other words, they occurred in less than 15 palm trees (Table 1).
Studies on pollination and insects which are visitors of other palm tree species like Acrocomia aculeata (Jacq.) (Scariot et al. 1991) and Attalea phalerata (Mart.) (Anderson et al. 1988) showed presence of Coleoptera, mainly of the families Curculionidae, Nitidulidae and Scarabaeidae. The Coleoptera were also more abundant in Butia yatay (Mart.) in Corrientes, Argentina (Bar 2001) and A. phalerata in the Pantanal, Brazil, (Santos et al. 2003) showing the predominance of this group amongst different palm tree species. The high number of Blattodea and Collembola can be explained by the accumulation of organic matter found in the base of the leaves of the palm trees, which could offer a lot of food for those detritivorous groups. The abundance of Hemiptera can be explained by the high frequency of individuals of hematophagous species Rhodnius neglectus (Lent) (Hemiptera: Reduviidae) and Psammolestes tertius (Lent & Jurberg) (Hemiptera: Reduviidae) (Gurgel-Gonçalves et al. 2004). Several studies on Triatominae populations were already made in palm trees, mainly of the genera Attalea, Acrocomia, Butia and Mauritia (Barretto et al. 1969, Carcavallo et al. 1998, Gurgel-Gonçalves et al. 2004) showing that the palm trees are favorable ecotope for the development of Triatominae colonies, vectors of the Chagas' disease.
The analysis of the orders of arthropods in the crown of M. flexuosa (Table 1) indicates two orders with the largest richness (Coleoptera and Araneae) which make up 86 morphospecies (63.7%). Three orders showed intermediate richness (Hymenoptera, Hemiptera and Blattodea) comprising 33 morphospecies (24.4%) and 10 orders low richness comprising 16 morphospecies (11.8%). Few morphospecies were abundant: 71 (52.6%) were represented by less than one individual per tree (Fig. 1). Coleoptera was the order with the largest morphospecies richness (43.7%) mainly of the families Staphylinidae and Curculionidae, followed by the order Araneae (20.0%) and the family Theridiidae as the most representative (Table 1). Coleoptera has been the order with the largest morphospecies richness in tropical canopies, including palm trees.
The largest richness of Staphylinidae and Curculionidae and the largest abundance of Tenebrionidae are verified in other studies (Basset 2001, Santos et al. 2003). The relative abundance of Araneae in relation to the total arthropod fauna was of 6.6%, a very close value to that obtained by Santos et al. (2003) from palm trees in the Pantanal region. However, in the present study, a larger richness of the family Theridiidae was observed, while in the Pantanal study the most diverse family was Salticidae. Theridiidae was also abundant in Goupia glabra (Aublet) (Asteraceae) in the Central Amazonian with 12 species (Hofer et al. 1994). Hymenoptera, Hemiptera and Blattodea presented larger abundance than richness of morphospecies. This low richness may be due to characteristics of the species of these groups that form enormous organized populations in colonies, as already suggested for Formicidae (Basset 2001). The other orders presented low relative abundance. According to Allison et al. (1997) and Novotny & Basset (2000), more than a half of the insect species in humid tropical forests are considered rare or unknown and that can be due to sampling problems resulting from the inadequacy of collecting methods. The number of rare species can also be increased by the visitor species that have come from adjacent plants (Shmida & Wilson 1985).
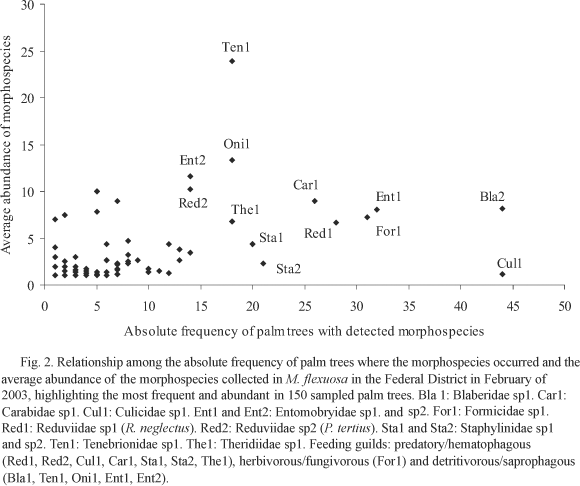
The relation between average morphospecies abundance and absolute frequency of palm trees where the morphospecies occurred (Fig. 2) indicates that there are both few frequent and abundant morphospecies in the community, which belonged to families with different feeding habits and different ecological characteristics. Similar analysis considering the palm trees as a group of a same environment shows that those species were more frequent in the rural environment (Fig. 3).
The analysis of the distribution of the arthropods in feeding guilds showed prevalence of predatory/ hematophagous (36.0%) followed by herbivorous/ fungivorous (34.0%) and detritivorous/ saprophagous (30.0%). The predatory/ hematophagous prevalence can be due to the high number of Coleoptera (mainly Carabidae and Staphylinidae), spiders (mainly Theridiidae) and Triatominae (R. neglectus and P. tertius). Among the main representatives of the guild of the herbivorous/ fungivorous were the arthropods of the families Formicidae (Hymenoptera), Curculionidae (Coleoptera) and Entomobryidae (Collembola) and the guild of the detritivorous/ saprophagous was represented mainly by Blaberidae (Blattodea), Tenebrionidae (Coleoptera) and Oniscidae (Isopoda). Santos et al. (2003) and Marques et al. (2001) showed prevalence of herbivores in A. phalerata and Vochysia divergens (Pohl) (Vochysiaceae) in Pantanal, however, in the palm tree A. phalerata there was a small difference in relation to percentage of predators. The percentage of detritivorous/ saprophagous arthropods found in M. flexuosa was almost three times larger than that observed in A. phalerata in Pantanal, and the abundance of Tenebrionidae among the decompositors was also shown by Erwin (1983).
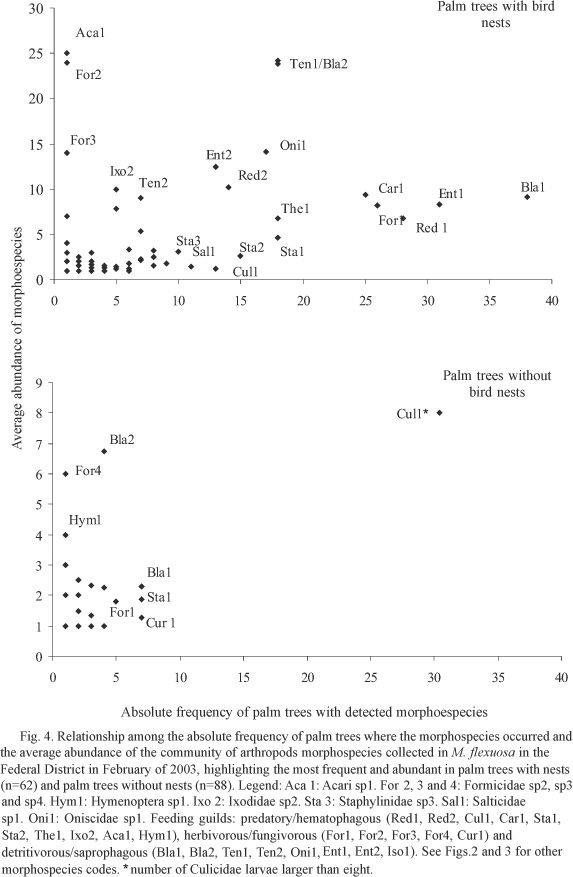
The palm trees of the "vereda" of Samambaia (periurban environment) showed the lowest abundance and morphospecies richness. Rajadinha presented the largest relative abundance of arthropods and the highest richness of morphospecies occurred in Alphaville (Table 2). In periurban environment we observed a largest frequency and abundance of larvae of Culicidae (Fig. 3). This is attributable to the presence of water in the base of the leaves. Morphospecies richness and average abundance of arthropods in palm trees with nests (n = 62) was about to 9.4 ± 7.1 morphospecies and 55.2 ± 73.9 individuals, superior values to the observed ones in palm trees without nests (n = 88): 1.3 ± 1.7 morphospecies and 2.2 ± 4.5 individuals. The number of bird nests showed positive correlation with richness and abundance of arthropods (Table 3). The areas that had larger richness and abundance (rural environment) were also the ones that presented more bird nests in the palm trees. Gurgel-Gonçalves et al. (2004) have also shown that the Triatominae distribution in M. flexuosa is not aleatory and it has been influenced by the presence of nests in the palm trees, mainly from the bird Phacellodomus ruber (Vieillot) (Passeriformes: Furnariidae). Other species are probably related to nests of P. ruber and the arthropod communities that live in the crown of M. flexuosa are clearly influenced by the presence of the nest of that bird species. The relation between average abundance of the morphospecies and absolute frequency of palm trees with nests and without nests shows that the composition of the arthropod fauna is quite different in those two situations, being richer and abundant in the palm trees with nests (Fig. 4). However, in other palm species the nests do not seem to be essential for the development of populations of insects, as Butia yatay (Mart.) (Arecaceae) (Bar 2001) and A. butyracea (Mutis ex L.f.) (Romaña et al. 1999).
The average richness considering the 150 sampled palm trees was about 4.7 ± 6.2 morphospecies per tree, which was noticeably inferior compared with the total morphospecies richness (135 morphospecies). The total richness estimated through the three methods was of 158 (ACE), 177 (Chao) and 204 (Jack) morphospecies. The average similarity among all palm trees was low (0.08). Those patterns of diversity indicate that each palm tree shelters a small number of the arthropods of the community present in M. flexuosa. Recently, Santos et al. (2003) described the arthropod diversity in the crown of A. phalerata in Pantanal, presenting 22 orders and 326 morphospecies in different feeding guilds, demonstrating that palm trees can be used as sources of food, reproduction places and shelters for the arthropod fauna. The differences in the observed patterns of diversity can be due to factors such as different sampling methods (canopy fogging x manual collect of the present material in the base of leaves of the palm tree), different savanna types (Pantanal x Cerrado) and microhabitats (A. phalerata x M. flexuosa).
Seasonality is a phenomenon observed among arthropods, mainly for insect species (Wolda & Wong 1988). Insect abundance can change over a time for a variety of reasons like macroclimatic changes and variation in the availability of food resources (Wolda 1988). The fieldwork was done during the wet season, a period known of high insect abundance in the tropics (Wolda 1978, Shelly 1988). Specifically in Brazilian Cerrado the increase of insect abundance in the wet season was observed for Coleoptera, Hemiptera and Isoptera (Pinheiro et al. 2002). Hence, this study presents the limitations shared by most one-sample community studies: underestimating species richness and giving a biased species abundance distribution pattern with few dominant species and many rare ones (Magurran 1988). A year-round sampling could evaluate more precisely the arthropod community in M. flexuosa crown but this sampling would be labor-expensive and have two trade-offs. Firstly, manual capture is a destructive sampling method, because it removes the substrate of the arthropod community - the organic matter accumulated in an individual palm tree crown. Therefore, the reposition of this organic matter is a slow process, delaying post-sampling recovery of arthropod assemblage in an individual palm crown, and precluding resampling of individual palm trees. Secondly, alternatively neighboring palm trees could be sampled in further sampling periods, but the number of M. flexuosa palm trees is limited. In the present one-sample study, 25 palm trees were sampled in each area (effective sampled area: 0.99-1.98/ ha), while M. flexuosa density ranged from 55 to 246 palm trees /ha in the study areas (Gurgel-Gonçalves et al. 2004). Hence, one sampling period leads to a considerable removal of favorable microhabitat. This strong ecological impact on the community would probably impoverish the natural arthropod assemblage in the further sampling periods.
The community of the arthropods in the crown of M. flexuosa presented two orders with the largest richness (Coleoptera and Araneae), three with intermediate (Hymenoptera, Hemiptera and Blattodea) and ten with lower richness level comprising only 16 morphospecies. The total richness observed was 135 morphospecies during the wet season. The community also showed both few frequent and abundant morphospecies, which belonged to families with different feeding habits and different ecological characteristics. The feeding guild more prevalent was predatory/ hematophagous ones. Bird nests seem to be an important factor for the establishment of the community differently of the measurements of the palm trees.
Besides arthropods, at least fifteen species of vertebrates were already observed associated with M. flexuosa such as Scinax sp. (Amphibia), Micrurus sp. (Reptilia), P. ruber, Gnorimopsar chopi (Vieillot), Polyborus plancus (Miller), Amazona aestiva (Linné) (Aves), Didelphis albiventris (Lund) and Rhipidomys aff. macrurus (Mammalia). They use the palm tree as nesting places, shelter or feeding sites (Prada 1994, Sick 1997). Prada (1994) studied M. flexuosa relating the production of fruits and the associated frugivore species concluding that M. flexuosa represents an important food source to the fauna and indicating the importance of this palm tree in terms of conservation of vertebrate fauna. However, the vertebrates represent less than 10% of the species richness that was already observed in that palm tree species while arthropods are more than 80%. So, M. flexuosa can be considered an important species for the maintenance of animal diversity, and this is one more argument in favor of the conservation of humid areas like the "veredas" of Cerrado biome.
Acknowledgments
Special thanks are due to Samuel, Marcos and Fabio (Universidade de Brasília) for their assistance in fieldwork. We thank Dra. Lourdes A. El-moor Loureiro (Universidade Católica de Brasília), Dra. Elena C. Visciarelli (Universidad Nacional del Sur Argentina) and the anonymous reviewers for critically reading the manuscript. We are grateful to the administrative staff of Reserva Ecológica do IBGE, Estação Ecológica Aguas Emendadas and farmers of the areas for entrance permission to conduct the study. This work was supported by CAPES, FINATEC and CNPq.
Received 05/VII/05. Accepted 21/XI/05.
- Adis, J., Y.D. Lubin & G.G. Montgomery. 1984. Arthropods from the canopy inundated and terra firme forests near Manaus, Brazil, with critical considerations on the pyrethrum-fogging technique. Stud. Neotrop. Fauna Environ. 19: 223-236.
- Allison, A., A. Samuelson & S.E. Miller. 1997. Patterns of beetle species diversity in Catanopsis acuminatissima (Fagaceae) trees studied with canopy fogging in mid-montane New Guinea rainforests, p.224-236. In N.E. Stork, J. Adis & R.K. Didham (eds.), Canopy arthropods. London, Chapman & Hall, 567p.
- Anderson, A.B., W.L. Overal & A. Henderson. 1988. Pollination ecology of a forest-dominant palm (Orbignya phalerata Mart.) in Northern Brazil. Biotropica 20: 192-205.
- Bar, M.E. 2001. Triatominos de la comunidad de palmeras en la provincia de Corrientes. Ecología e importancia epidemiológica. Tesis doctoral, Facultad de Ciencias Exactas y Naturales y Agrimensura, Universidad Nacional del Nordeste, Corrientes, Argentina, 255p.
- Barretto, M.P., D.R.R. Albuquerque & G.K. Funayama. 1969. Estudos sobre reservatórios e vectores silvestres do Trypanosoma cruzi XXXVI: Investigação sobre triatomíneos de palmeiras no município de Uberaba, MG, Brasil. Rev. Bras. Biol. 29: 577-588.
- Barros, H.R.P., I.R. Diniz & K. Kitayama. 1992. Nest density of some social wasp species in Cerrado vegetation of Brazil (Hymenoptera, Vespidae). Entomol. Generalis 17: 265-266.
- Basset, Y. 2001. Invertebrates in the canopy of tropical forests: How much do we really know ? p.7-107. In K.E. Linsemair, A.J. Davies, B. Fiala & R.R. Speight (eds.), Tropical forest canopies: Ecology and management. London, Kluwer Academic Publisher, 370p.
- Battirola, L.D., M.I. Marques, J. Adis & J.H.C. Delabie. 2005. Composição da comunidade de Formicidae (Insecta, Hymenoptera) em copas de Attalea phalerata Mart. (Arecaceae) no Pantanal de Poconé, Mato Grosso, Brasil. Rev. Bras. Entomol. 49: 107-117.
- Camargo, A.J.A. & Becker, V.O. 1999. Saturniidae (Lepidoptera) from the Brazilian cerrado: Composition and biogeographic relationships. Biotropica 31: 696-705.
- Carcavallo, R.U., M.E. Franca Rodríguez, R. Salvatella, S.I. Curto de Casas, I.A. Sherlock, C. Galvão, D.S. Rocha, I. Galíndez Giron, M.A. Otero Arocha, A. Martínez, J.A. da Rosa, D.M. Canale, T.H. Farr & J.M.S. Barata. 1998. Habitats and related fauna, p.561-600. In R.U. Carcavallo, I. Galíndez Girón, J. Jurberg & H. Lent (eds.), Atlas of Chagas disease vectors in Americas v. II. Rio de Janeiro, Editora Fiocruz, 324p.
- Chaves, Z.M. 1998. Fungos associados a Mauritia flexuosa (Buriti) e Mauritiella armata (Buritiana). Dissertação de mestrado. Universidade de Brasília, Brasília, 104p.
- Colwell, R.K. 1997. Estimates: Estatistical Estimation of Species Richness and Shared Species from Samples. Version 5. Guia do Usuário e programas disponíveis in http://viceroy.eeb.uconn.edu/estimates04/04/2005.
- Diniz, I.R. & A. Raw. 1982. Dinâmica de populações da vespa solitária Zeta argillaceum (Linnaeus 1758) (Hymenoptera, Eumenidae). An. Soc. Entomol. Bras. 11: 57-68.
- Diniz, I.R., H.C. Morais & A.J.A. Camargo. 2001. Host plants of lepidopteran caterpillars in the Cerrado of the Distrito Federal, Brazil. Rev. Bras. Entomol. 45: 107-122.
- Erwin, T.L. 1983. Tropical forest canopies: The last biotic frontier. Bull. Entomol. Soc. 30: 14-19.
- Gurgel-Gonçalves, R., A.R.T. Palma, M.N.A. Menezes, R.N. Leite, C.A.C. Cuba. 2003. Sampling Rhodnius neglectus (Triatominae) in Mauritia flexuosa palm trees (Arecaceae): A field study in the Brazilian Savanna. Med. Vet. Entomol. 17: 347-349.
- Gurgel-Gonçalves, R., M.A. Duarte, E.D. Ramalho, A.R.T. Palma & C.A.C. Cuba. 2004. Distribuição espacial de populações de Triatominae (Hemiptera: Reduviidae) em palmeiras da espécie Mauritia flexuosa no Distrito Federal. Rev. Soc. Bras. Med. Trop. 37: 241-247.
- Henderson, A., G. Galeano & R. Bernal. 1995. Field guide to the palms of the Americas. Princeton Univ. Press, Princeton, 351p. pls 1-64.
- Hofer, H., A.D. Brescovit, J. Adis & W. Paarmann. 1994. The spider fauna of Neotropical tree canopies in Central Amazonia: First results. Stud. Neotrop. Fauna Environ. 29: 23-32.
- Hurtado Guerreiro, J.C., C.R.V. da Fonseca, P.M. Hammond & N.E. Stork. 2003. Seasonal variation of canopy arthropods in Central Amazon, p.170-175. In Y Basset , V.R. Novotny, S.E. Miller & R.L. Kitching (eds), Arthropods of tropical forests. Spatio-temporal dynamics and resource use in the canopy. Cambrige, Cambrige University Press, 474 p.
- Jepson, W. 2005. A disappearing biome? Reconsidering land-cover change in the Brazilian savanna. Geograph. J. 171: 99-111.
- Klink, C.A. & A.G. Moreira 2002. Past and current human occupation and land use. p.69-88. In P.S. Oliveira & R.J. Marquis (eds.), The Cerrados of Brazil: Ecology and natural history of a Neotropical savanna. New York, Columbia University Press, 373p.
- Klink, C.A. & R.B. Machado. 2005. Conservation of the Brazilian Cerrado. Conserv. Biol. 19: 707-713.
- Magurran, A.E. 1988. Ecological diversity and its measurement. New Jersey, Princeton University press, 179p.
- Marques, M.I., J. Adis, C. Nunes da Cunha & G.B. Santos. 2001. Arthropod biodiversity in the canopy of Vochysia divergens Pohl (Vochysiaceae), a forest dominant in the Brazilian Pantanal. Stud. Neotrop. Fauna Environ. 36: 205-210.
- Milhomem, M.S. 2003. A fauna de Scarabaeidae sensu stricto (Coleoptera, Scarabaeidae) do Cerrado de Brasilia, DF: Variação anual, efeito do fogo e da cobertura vegetal. Tese de doutorado, Universidade de Brasília, Brasília, 92p.
- Moran, V.C. & T.R.E. Southwood. 1982. The guild composition of arthropod communities in trees. J. Animal Ecol. 51: 289-306.
- Motta, P.C. 2002. Butterflies from the Uberlândia region, Central Brazil: Species list and biological comments. Braz. J. Biol. 62: 151-163.
- Novotny, V. & Y. Basset. 2000. Rare species in communities of tropical insect herbivores: Pondering the mystery of singletons. Oikos 89: 564-572.
- Oliver, I & A J. Beattie 1996. Invertebrate morphospecies as surrogates for species: A case study. Conserv. Biol. 10: 99-109.
- Oliveira, P.S. & R.J. Marquis. 2002. The Cerrados of Brazil: Ecology and natural history of a Neotropical savanna. New York, Columbia University Press, 373p.
- Pinheiro, F., I.R. Diniz & K. Kitayama. 1998. Comunidade local de Coleoptera no Cerrado: Diversidade de espécies e tamanho de corpo. An. Soc. Entomol. Bras. 27: 543-550.
- Pinheiro, F., I.R. Diniz, D. Coelho, M.P.S. Bandeira. 2002. Seasonal pattern of insect abundance in the Brazilian cerrado. Austral Ecol. 27: 132-136.
- Prada, M.V. 1994. Guilda de frugívoros associada com o buriti (Mauritia flexuosa: Palmae) numa "vereda" no Brasil Central. Dissertação de mestrado, Universidade de Brasília, Brasília, 99p.
- Ribeiro, J.F. & B.M.T. Walter. 1998. Fitofisionomias do bioma Cerrado. Cerrado Ambiente e Flora, p.89-166. In S.M. Sano & S.P. Almeida (eds), Cerrado Ambiente e Flora, Embrapa, Planaltina, 556p.
- Romaña, C.A., J.C.N. Pizarro, E. Rodas & E. Guilbert .1999. Palm trees as ecological indicator of risk areas for Chagas disease. Trans. R. Soc.Trop. Med. Hyg. 93: 594-595.
- Santos, G.B., M.I. Marques, J. Adis & C.R. De Musis. 2003. Artrópodos associados à copa de Attalea phalerata Mart. (Arecaceae), na região do Pantanal de Poconé, Mato Grosso, Brasil. Rev. Bras. Entomol. 47: 211-224.
- Scariot, A., J. Lleras & J. Hay. 1991. Reproductive biology of the palm Acrocomia aculeata in Central Brazil. Biotropica. 23: 12-22.
- Shelly, T. 1988. Relative abundance of day-flying insects in treefall gaps vc shaded understory in a neotropical forest. Biotropica 20: 114-119.
- Shmida, A. & M. R. Wilson. 1985. Biological determinants of species diversity. J. Biogeogr. 12: 1-20.
- Sick, H. 1997. Ornitologia brasileira. Rio de Janeiro, Edição revista e ampliada por J.F. Pacheco. Editora Nova Fronteira, Rio de Janeiro, 862p.
- Silva F.J. 2001. Fauna de curculionídeos no Cerrado de Brasília. Dissertação de mestrado, Universidade de Brasília, Brasília, 60 p.
- Silvestre, R. & C.R.F. Brandão. 2001. Formigas (Hymenoptera, Formicidae) atraídas a iscas em uma ilha de cerrado no município de Cajuru, estado de São Paulo, Brasil. Rev. Bras. Zool. 44: 71-77
- Wolda, H. 1978. Seasonal fluctuations in rainfall, food and abundance of tropical insects. J. Anim. Ecol. 47: 369-381.
- Wolda, H. 1988. Insect seasonality: Why? Annu. Rev. Ecol. Syst. 19: 1-18.
- Wolda, H & M. Wong. 1988. Tropical insect diversity and seasonality, sweep-samples vs light traps. Proc. Konin Nederl. Akad. Wetens. 91: 203-16.
- Zar, J.H. 1996. Biostatistical analysis. Third edition. Prentice Hall International, INC, Upper Saddle River, New Jersey, 662p.
Publication Dates
-
Publication in this collection
31 July 2006 -
Date of issue
June 2006
History
-
Accepted
21 Nov 2005 -
Received
05 July 2005