Abstracts
Tuta absoluta (Meyrick) is an important tomato pest that also feeds on other host-plants from the Solanceae family. We studied the effect of two cultivated plants, tomato (Lycopersicum esculentum Mill.) and potato Solanum tuberosum L. on the development and populational parameters of T. absoluta related with host-plant suitability. Larval developmental time, pupal weight, mean fecundity and an index of host-plant quality (IPQ = pupal weight / frass weight) were estimated. Age-specific survivorship and fecundity life tables were constructed in the laboratory to evaluate the following populational parameters: net reproductive rate (Ro), intrinsic rate of increase (r) and generation time (T). Larval developmental time was shorter and pupal weight was higher (P < 0.0001) for larvae reared on tomato (P < 0.0001). Mean fecundity was not significantly different on both plants (P = 0.07) and food quality of host-plant was higher for tomato (P = 0.02). Mean population parameters on tomato were: Ro = 48.92; T = 27.98, r = 0.14; and on potato: Ro = 14.43; T = 32.35, r = 0.08. Although results showed that tomato was a more suitable host-plant and had a better nutritional quality than potato, when T. absoluta fed on potato the potential population increase requires attention. Under appropriate climatic conditions, spatial and temporal coincidence between crop and pest, T. absoluta could become a pest for the potato crop.
Insecta; host range; Lycopersicum esculentum; Solanum tuberosum; Integrated Pest Management
Tuta absoluta (Meyrick) es una importante plaga del tomate que puede utilizar también otras solanáceas como plantas huéspedes. Este trabajo evaluó el efecto del tomate, Lycopersicum esculentum Mill. y la papa, Solanum tuberosum L., sobre algunos aspectos del desarrollo y parámetros poblacionales relacionados con la calidad del alimento. Se midieron: duración del estado larval, peso pupal, fecundidad y un índice de la calidad alimentaria de la planta huésped (peso pupal / peso de heces). Por medio de tablas de vida en el laboratorio se obtuvieron datos de supervivencia y fecundidad específicas por edad con los cuales se calcularon: tasa reproductiva neta (Ro), tasa intrínseca de crecimiento (r) y tiempo generacional (T). Los individuos alimentados con tomate tuvieron una menor duración del estado larval (P < 0,0001), y un mayor peso pupal (P < 0,0001) que con papa. La fecundidad promedio fue marginalmente más alta en tomate (P = 0,07) mientras que la calidad nutricional fue mayor para el tomate (P = 0,025). Los parámetros poblacionales promedio fueron: Ro = 48,92; T = 27,98, r = 0,14, en tomate; y Ro = 14,43; T = 32,35, r = 0,08 en papa. Si bien los resultados indicaron que el tomate exhibió una mayor calidad nutricional que la papa, las cohortes de T. absoluta alimentadas con este huésped secundario, mostraron un crecimiento relativamente importante. Bajo condiciones climáticas favorables y una coincidencia espacial y temporal entre la polilla y el cultivo de papa, sus poblaciones podrían convertirse en una plaga para el mismo.
Insecta; amplitud de dieta; Lycopersicum esculentum; Solanum tuberosum; Manejo Integrado de Plagas
CROP PROTECTION
Effect of two solanaceous plants on developmental and population parameters of the tomato leaf miner, Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae)
Efecto de dos solanáceas cultivadas sobre el desarrollo y los parámetros poblacionales de la polilla del tomate, Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae)
Patricia C. Pereyra; Norma E. Sánchez
CEPAVE (Centro de Estudios Parasitológicos y de Vectores), UNLP-CONICET 2 no. 584 (1900) La Plata, Argentina, ppereyra@netverk.com.ar
ABSTRACT
Tuta absoluta (Meyrick) is an important tomato pest that also feeds on other host-plants from the Solanceae family. We studied the effect of two cultivated plants, tomato (Lycopersicum esculentum Mill.) and potato Solanum tuberosum L. on the development and populational parameters of T. absoluta related with host-plant suitability. Larval developmental time, pupal weight, mean fecundity and an index of host-plant quality (IPQ = pupal weight / frass weight) were estimated. Age-specific survivorship and fecundity life tables were constructed in the laboratory to evaluate the following populational parameters: net reproductive rate (Ro), intrinsic rate of increase (r) and generation time (T). Larval developmental time was shorter and pupal weight was higher (P < 0.0001) for larvae reared on tomato (P < 0.0001). Mean fecundity was not significantly different on both plants (P = 0.07) and food quality of host-plant was higher for tomato (P = 0.02). Mean population parameters on tomato were: Ro = 48.92; T = 27.98, r = 0.14; and on potato: Ro = 14.43; T = 32.35, r = 0.08. Although results showed that tomato was a more suitable host-plant and had a better nutritional quality than potato, when T. absoluta fed on potato the potential population increase requires attention. Under appropriate climatic conditions, spatial and temporal coincidence between crop and pest, T. absoluta could become a pest for the potato crop.
Key words: Insecta, host range, Lycopersicum esculentum, Solanum tuberosum, Integrated Pest Management
RESUMEN
Tuta absoluta (Meyrick) es una importante plaga del tomate que puede utilizar también otras solanáceas como plantas huéspedes. Este trabajo evaluó el efecto del tomate, Lycopersicum esculentum Mill. y la papa, Solanum tuberosum L., sobre algunos aspectos del desarrollo y parámetros poblacionales relacionados con la calidad del alimento. Se midieron: duración del estado larval, peso pupal, fecundidad y un índice de la calidad alimentaria de la planta huésped (peso pupal / peso de heces). Por medio de tablas de vida en el laboratorio se obtuvieron datos de supervivencia y fecundidad específicas por edad con los cuales se calcularon: tasa reproductiva neta (Ro), tasa intrínseca de crecimiento (r) y tiempo generacional (T). Los individuos alimentados con tomate tuvieron una menor duración del estado larval (P < 0,0001), y un mayor peso pupal (P < 0,0001) que con papa. La fecundidad promedio fue marginalmente más alta en tomate (P = 0,07) mientras que la calidad nutricional fue mayor para el tomate (P = 0,025). Los parámetros poblacionales promedio fueron: Ro = 48,92; T = 27,98, r = 0,14, en tomate; y Ro = 14,43; T = 32,35, r = 0,08 en papa. Si bien los resultados indicaron que el tomate exhibió una mayor calidad nutricional que la papa, las cohortes de T. absoluta alimentadas con este huésped secundario, mostraron un crecimiento relativamente importante. Bajo condiciones climáticas favorables y una coincidencia espacial y temporal entre la polilla y el cultivo de papa, sus poblaciones podrían convertirse en una plaga para el mismo.
Palabras clave: Insecta, amplitud de dieta, Lycopersicum esculentum, Solanum tuberosum, Manejo Integrado de Plagas
The tomato leafminer, Tuta absoluta (Meyrick), is a major pest of tomato, Lycopersicum esculentum Mill. in South America (Suinaga et al. 1999, Torres et al. 2001). This microlepidopterous of neotropical distribution (Razuri & Vargas 1975, Moore 1983, Souza & Reis 1986) is a multivoltine species that mines leaves, fruits, flowers, buds and stems. The damage is produced when the larvae feed on the leaf mesophyll expanding mines, thus affecting the photosynthetic capacity of the crop with subsequent reduction of yield. Moreover, injury made directly to the fruits causes severe losses (Colomo & Berta 1995).
T. absoluta is a relative recently tomato pest in Argentina. The first register of its occurrence in Mendoza was during the ´60. Few studies regarding to the knowledge of T. absoluta biology and ecology (Bahamondes & Mallea 1969, Pereyra 2002), natural enemies (Botto 1999, Colomo et al. 2002) and chemical control (Lietti et al. 2005) have been developed in this country.
T. absoluta is an oligophagous insect (Siqueira et al. 2000) that feeds on solanaceous species. Other cultivated host-plants, besides tomato, are mentioned in the literature (Vargas 1970, Mallea et al. 1972, Campos 1976) as potato, Solanum tuberosum L., egg-plant S. melongena L. and tobacco, Nicotiana tabacum L. Among wild host-plants, S. nigrum L., S. eleagnifolium L., S. bonariense L., S. sisymbriifolium Lam., S. saponaceum, Lycopersicum puberulum Ph., Datura ferox L., D. stramonium L. and Nicotiana glauca Graham are cited (García & Espul 1982). However, little is known about the trophic relationship between T. absoluta and its potential host-plants, especially other crops; in Perú, this pest already becomes a serious pest of potato crop (Campos 1976, Cisneros & Mujica 1998).
Different patterns of host-plant used by herbivorous insects are related to differences in physiology, morphology, chemical and physical defenses of the plants (Singer et al. 2004), with subsequent differences in the herbivorous life history traits. Quality and quantity of food consumed by herbivores affect the growth, survival and fecundity of individuals, and hence their populational improvement (Scriber & Slansky 1981, Slansky 1993). Particularly, those population parameters that express the potential capacity of growth: intrinsic growth rate, net reproductive rate and generation time, are related to the suitability of the cultivated host-plants consumed (Sauvion et al. 2005). The understanding of the host-plant range and the patterns of crop utilization, could allow predicting the possible expansion to other crops, resulting of great importance in the Integrated Pest Management (Hill et al. 2004).
The objective of this research was to study the effect of tomato, L. esculentum, and potato, S. tuberosum, both cultivated host-plants, on some development attributes and population parameters of T. absoluta, related with the host-plant suitability.
Materials and Methods
Two colonies of T. absoluta have been reared for five generations with tomato (var. Platense) and potato (var. Spunta) leaves under laboratory conditions (25 ± 1ºC; 60 ± 5 % R.H.; photoperiod: 16 L: 8 D). Tomato and potato plants were greenhouse grown in pots (20 cm diam.) with fertilized soil, being watered three times a week.
T. absoluta suitability and nutritional value of the host-plants were measured as suggested by Scriber & Slansky (1981) and Pereyra (1995). Mean developmental time (from hatching to pupation), pupal weight 48h after pupation (mg) and fecundity (total number of eggs / female) for individuals reared with both host-plants were assessed. Fecundity was estimated on one female maintained in plastic bottles (1 L.) with two males containing leaves of each host-plant placed in plastic containers for oviposition. Adults were fed daily with a sugar solution (15%). An index of plant quality (IPQ) was calculated for each host-plant following Tuomi et al. (1981) and Koricheva & Haukioja (1992) formulae where: IPQ = pupal weight (mg) / frass dry weight (mg). To estimate the nutritional value of the leaf, frass of leafminer entire lifetime was collected inside of mines. The effect of different host-plants for each variable was compared by ANOVA (Zar 1999).
For the assessment of populational parameters, life tables of T. absoluta for both host-plants were constructed under laboratory conditions. Newly emerged adult females were maintained as mentioned above. Eggs collected on tomato and potato leaves were divided in three cohorts of 49, 35 and 28, and 46, 39 and 30 eggs, respectively. Each egg was placed individually in a Petri dish (10 cm diam) containing moistened filter paper. Larvae were reared individually from hatching to pupation with the corresponding host-plant and fed ad-libitum. The age-specific survivorship (lx: proportion of individuals alive at age x) and the age-specific fecundity of adult females (mx = number of eggs / female / day) of each cohort were daily registered. Mean survival (lx) and fecundity (mx) curves of cohorts reared with tomato and potato leaves were compared through a Chi square test (Zar 1999). The population parameters as net reproductive rate: Ro = å lx.mx (mean number of female offspring / female/ lifetime), mean generation time (days): T = å x . lx . mx / R0 (mean time between two successive generations) and intrinsic rate of increase (r): å e -rx . lx . mx = 1 (instant rate of population change per individual, calculated with the Lotka equation) were calculated for each cohort of each host-plant. The mean values of the population parameters for the three cohorts of each host-plant were compared with ANOVA test (Zar 1999).
Results
Larvae fed on with tomato had a significantly lower mean developmental time than those fed on potato (ANOVA: F = 72.16; d.f. = 1,40; P < 0.0001). Pupal weight was significantly higher for individuals reared tomato (ANOVA: F = 21.83; d.f. = 1,40; P < 0.0001) (Table 1). Total number of eggs of females reared with both host-plants (Table 1) were not significantly different by ANOVA (F = 3.55; d.f. = 1,23; P = 0.07). The relationship pupal weight / frass weight (IPQ) was higher in tomato (F = 5.59; d.f. = 1,30; P < 0.025), indicating the higher nutritional quality of this host-plant for T. absoluta (Table 1).
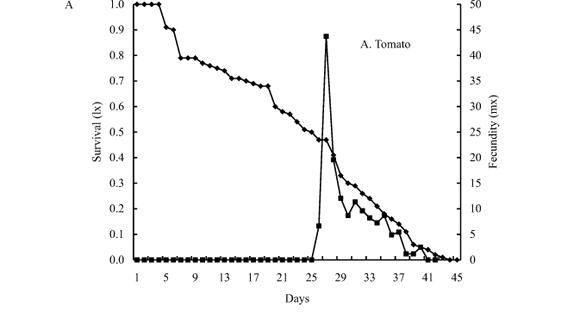
Mean survival curves of cohorts reared with tomato and potato were similar (X2 = 12.52; d.f. = 45; P > 0.05) (Fig. 1A,B ). The survival curve (lx) of T. absoluta individuals fed on tomato extended until day 43, and remained high most of the lifetime, decreasing to 50% at day 25 (Fig. 1A). On the other hand, the mean survival curve (lx) of the cohorts fed potato reached 47 days but decreased abruptly to 50% during the first stages of development at day 17 (Fig. 1B ).
Females fed on tomato during the larval stage showed a very different oviposition pattern than females fed on potato (X2 = 126.16; d.f. = 21; P < 0.001). Although total fecundity was not statistically different between host-plants, the age-specific fecundity curve (mx) was significantly higher on tomato, reaching a peak of 44 eggs / female at day 27 (Fig. 1A). The curve of females reared on tomato reached one peak of 15 eggs / female at day 30, and a smaller one of 10 eggs / female between days 40 and 42 (Fig. 1B ).
The estimated values of life table parameters for T. absoluta cohorts reared on tomato and potato are presented in Table 2. Potential population growth of T. absoluta fed on tomato as intrinsic rate of increase (r: F = 110.45, d.f. = 1,4) and net reproductive rate (Ro: F = 36.34, d.f. = 1,4) were significantly higher on tomato, while mean generation time was lower (T: F = 303.87, d.f. = 1,4).
Discussion
T. absoluta is found almost exclusively on tomato in Argentina. Although this study shows that tomato is a more suitable host-plant than potato, it also demonstrated that T. absoluta populations could develop and grow on potato. Hence, this species could result a potential pest for this crop, as happens in other countries, as Peru (Campos 1976, Cisneros & Mujica 1998).
Many solanaceous have been cited as host-plants (Colomo et al. 2002), however studies regarding its suitability to insects are lacking and thus, the host range of T. absoluta is not well defined. Eggplant (S. melongena) and the wild S. bonariense, mentioned as host-plants (García & Espul 1982) could not be included in the present study as in a preliminary test, because adult females of T. absoluta did not oviposit on eggplant leaves and the larvae forced to feed on them could not reach the complete development in laboratory.
The high nutritional quality of tomato had positive effects on the development (less developmental time and higher pupal weight) of T. absoluta, and on the capacity of population increase (higher Ro and r, and lower T). A higher pupal weight of individuals fed on tomato could explain a trend to higher fecundity in a conventional and generally acceptable F-value (0.05), although marginally statistical differences between the two host-plants of 35.05 were observed. Previous studies developed in our laboratory demonstrated that pupal weight and fecundity of T. absoluta were related, since 71% of variation in fecundity was explained by the pupal weight. This means that those females that reached higher pupal masses were able to lay more eggs for the future generation (Pereyra 2002). This positive relationship between pupal weight and fecundity has also been observed for other lepidopteran species (Bessin & Reagan 1990, Awmack & Leather 2002). Pupal weight of individuals reared on tomato var. Platense were similar to those found on var. Santa Cruz Kada AG-373 (Giustolin & Vendramin 1994) and the mean value found in this study was higher than the one mentioned by Giustolin et al. (1995) under conditions of shorter photophase.
Demographic parameters obtained from cohorts reared on different host-plants under laboratory conditions are useful for the assessment of the biotic potential of an insect facing a new food and for the evaluation of the host-plant quality (Sánchez et al. 1997 Sánchez & Pereyra 1995). Intrinsic rate of increase and mean generation time reflect the suitability of the host-plant (Tsai 1998) as, among other factors, survivorship and fecundity are affected by the host-plant nutritional value. Survival curves of cohorts reared on tomato maintained high values until the reproductive age, suggesting an elevated capacity of population increase. A shorter generation time (T) on tomato would also reduce the time of exposure of T. absoluta to natural enemies under field conditions (Price et al. 1980). This, in turn means a better fitness in terms of survival and fecundity and an increased probability of leaving offspring.
Standard indices of plant digestibility and efficiency of food conversion to insect biomass used for nutritional ecology (Pereyra 1995), based their determinations on consumed foliage, are difficult to measure in leaf-miners. When they present sedentary life-style and locally limited feeding, the exact feeding efficiency is not required; however it is important to compare them under different conditions (i.e. host-plants) (Koricheva & Haukioja 1992). The IPQ is an appropriate nutritional food quality measurement for two host-plants comparisons. The higher IPQ ratio obtained by tomato-fed indicates better food quality than potato-fed T. absoluta.
Many herbivorous insects that expand their diets including cultivated host-plants, tightly related with their wild host-plants, are able to reach pest status. The shift of the herbivore diet to a cultivated plant in general represents a rich and concentrate food resource, positively affecting herbivore growth. One example is the Colorado potato beetle, Leptinotarsa decemlineata (Say) (Coleoptera: Chrysomelidae) in the Northern Hemisphere (Hsiao 1978, Ferro 1987), that usually native solanaceous feeder, shifting to the introduced cultivated potato from the peruvian altiplane, and rapidly becoming a key pest. Population of Colias philodice eriphyle Edwards (Lepidoptera: Pieridae), in North America, that only fed on wild leguminosae expanded its diet and included cultivated alfalfa (Tabashnik 1983). Cultivated or wild alternative host-plants play the role of natural reservoirs and assure the persistence of the populations in certain areas, even when the main crop is not present.
T. absoluta is reported as feeder of potato leaves, stems and tubers in Colombia, Peru and Chile (García & Espul 1982, Campos 1976). In Argentina, T. absoluta has not been considered a potato pest even after it was observed feeding on aerial parts of the plant in the Northern region of the country (Colomo et al. 2002). In this study, the cohorts of T. absoluta fed on potato developed and grew satisfactorily under laboratory conditions, suggesting that under favorable field conditions, such as, climate, geographical and phenological coincidence, resource concentration, elimination of natural enemies through the abusive use of pesticides, etc., the populations of T. absoluta could become a real problem to the crop.
Acknowledgements
We wish to thank Jimena Rosso for technical assistance and Ing. Daniel Caldiz for comments.
Received 20/VI/05.
Accepted 02/I/06.
- Awmack, C.S. & S.R. Leather. 2002. Host plant quality and fecundity in herbivorous insects. Ann. Rev. Entomol. 47: 817-844.
- Bahamondes, L.A. & A.R. Mallea. 1969. Biología en Mendoza de Scrobipalpula absoluta (Meyrick) Povolny (Lepidoptera: Gelechiidae), especie nueva para la República Argentina. Rev. Fac. Cs. Agrarias, UNC (Argentina) 15: 96-104.
- Botto, E.N. 1999. Control biológico de plagas hortícolas en ambientes protegidos. Rev. Soc. Entomol. Argent. 58: 58-64.
- Bessin, R.T. & T.E. Reagan. 1990. Fecundity of sugarcane borer (Lepidoptera: Pyralidae), as affected by larval development on gramineous host plants. Environ. Entomol. 19: 635 639.
- Campos, R.G. 1976. Control químico del "minador de hojas y tallos de la papa" (Scrobipalpula absoluta Meyrick) en el valle del Cañete. Rev. Per. Entomol. 19: 102-106.
- Cisneros, F. & N. Mujica. 1998. The leafminer fly in potato plant reactions and natural enemies as natural mortality factors. CIP (Peru) Program report 1997-98: 129-140.
- Colomo, M.V. & D.C. Berta. 1995. Fluctuación de la población de Scrobipalpula absoluta (Meyrick) (Lepidoptera, Gelechiidae) en plantaciones de tomate en el Departamento de Lules, Tucumán. Acta Zool. Lilloana 43: 165-177.
- Colomo, M.V., D.C. Berta & M.J. Chocobar. 2002. El complejo de himenópteros parasitoides que atacan a la "polilla del tomate" Tuta absoluta (Lepidoptera: Gelechiidae) en la Argentina. Acta Zool. Lilloana 46: 81-92.
- Ferro, D.N. 1987. Insect pest outbreaks in agroecosystems. In P. Barbosa & J.C. Schultz. (eds.), Insect outbreaks. San Diego, Academic Press, 578p.
- Garcia, M.F. & J.C. Espul. 1982. Bioecología de la polilla del tomate (Scrobipalpula absoluta) en Mendoza, República Argentina. Rev. Invest. Agropecuarias INTA (Argentina) XVII: 135-146.
- Giustolin, T. A. & J.D. Vendramin. 1994. Efeito de duas espécies de tomateiro na bilologia de Scrobipalpuloides absoluta (Meyrick). An. Soc. Entomol. Brasil 23: 511-517.
- Giustolin, T.A., J.D. Vendramin & J.R. Parra. 1995. Desenvolvimento de uma dieta artificial para estudios do efeito de aleloquímicos sobre Scrobipalpuloides absoluta (Meyrick). An. Soc. Entomol. Brasil 24: 265-272.
- Hill, C.B., Y. Li & G.L. Hartman. 2004. Resistance of Glycine species and various cultivated legumes to the soybean aphid (Homoptera: Aphididae). J. Econ. Entomol. 97: 1071-1077.
- Hsiao, T.H. 1978. Host plant adaptations among geographic populations of the Colorado potato beetle. Entomol. Exp. Appl. 24: 237-247.
- Koricheva, J. & E. Haukioja. 1992. Effects of air pollution on host plant quality, individual performance and population density of Eriocrania miners (Kepidoptera: Eriocraniidae). Environ. Entomol. 21: 1386 1392.
- Leite, G. L., M. Picanco, J.C. Zanuncio, G.N. Jham & M.F. Moura. 1999. Efecto de los niveles de fertilización en la intensidad de ataque de Tuta absoluta en Lycopersicum hirsutum y L. esculentum Man. Integr. Plagas 53: 72-76.
- Lietti, M.M., E. Botto & R.A. Alzogaray. 2005. Insecticide resistance in argentine populations of Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae). Neotrop. Entomol. 34: 113-119.
- Mallea, A.R, G.S. Mácola, J.C. García, L.A. Bahamondes & J.H Suárez. 1972. Nicotiana tabacum L. var. Virginica, nuevo hospedero de Scrobipalpula absoluta (Meyrick) Povolny (GelechiidaeLepidoptera). Rev. Fac. Cs. Agrarias (Argentina) 18: 13-15.
- Moore, J.E. 1983. Control of tomato leafminer (Scrobipalpula absoluta) in Bolivia. Trop. Pest Manage. 29: 231-238.
- Pereyra, P.C. 1995. Ecología nutricional de la "oruga medidora" Rachiplusia nu (Lepidoptera: Noctuidae). Rev. Soc. Entomol. Argent. 54: 31-40.
- Pereyra, P.C. 2002. Evidencia de la competencia intraespecífica en estadios larvales tempranos de la polilla del tomate, Tuta absoluta (Lepidoptera: Gelechiidae). Ecología Austral 12: 143-148.
- Price, P.W., C.E. Bouton, P. Gross, B.A. Mcpheron, J.N. Thompson & A.E. Weis. 1980. Interactions among three trophic levels: Influence of plants on interactions between insect herbivores and natural enemies. Ann. Rev. Ecol. Syst. 11: 41 65.
- Razuri, V. & E. Vargas. 1975. Biologia e comportamiento de Scrobipalpula absoluta (Meyrick) (Lepidoptera: Gelechiidae) en tomatera. Rev. Peruana Entomol. 18: 84-89.
- Sánchez, N.E. & P.C. Pereyra. 1995. Life table of the soybean looper Rachiplusia nu (Lepidoptera: Noctuidae) in the laboratory. Rev. Soc. Entomol. Argent. 54: 89-96.
- Sánchez, N.E., P.C. Pereyra & M.V. Gentile. 1997. Population parameters of Epinotia aporema (Lepidoptera: Tortricidae) on soybean. Rev. Soc. Entomol. Argent. 56: 151-153.
- Sauvion, S., V. Mauriello, B. Renard & N. Boissot. 2005. Impact of melon accessions resistant to aphids on the demographic potential of silverleaf whitefly. J. Econ. Entomol. 98: 557-567.
- Scriber, J.M. & F. Slansky. 1981. The nutritional ecology of immature insects. Ann. Rev. Entomol. 26: 183-211.
- Singer, M.S., D. Rodrigues, J.O. Stireman III & Y. Carriere. 2004. Roles of food quality and enemy-free space in host use by a generalist insect herbivore. Ecology 85: 2747-2753.
- Siqueira, H.A., R.N. Guedes & M.C. Picanço. 2000. Insecticide resistance in populations of Tuta absoluta (Lepidoptera: Gelechiidae). Agric. and Forest Entomol. 2: 147-153.
- Slansky, F. 1993. Nutritional ecology: The fundamental quest for nutrients, p.29-91. In N.E. Stamp & T.M. Casey (eds.), Caterpillars. Ecological and evolutionary constrains on foraging. New York, Chapman & Hall, 587p.
- Souza, J.C. & P.R. Reis. 1986. Controle da traça do tomateiro em Minas Gerais. Pesq. Agropec. Bras. 21: 343-354.
- Suinaga, F.A., M. Picanço, G.N. Jham & S.H. Brommonschenkel. 1999. Causas químicas de resistencia de Lycopersicum peruvianum (L.) a Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae). An. Soc. Entomol. Brasil 28: 313-321.
- Tabashnik, B. 1983. Host range evolution: Shift from native legume host to alfalfa by the butterfly, Colias philodice eryphile Evolution 37: 150-162.
- Torres, J.B., C.A. Faria, W.S. Evangelista & D. Pratissoli. 2001. Within-plant distribution of the leaf miner Tuta absoluta (Meyrick) immatures in processing tomatoes, with notes on plant phenology. Int. J. Pest Manag. 47: 173-178.
- Tsai, J.H. 1998. Development, survivorship and reproduction of Toxoptera citricida (Kirkaldy) (Homoptera: Aphididae) on eight host plants. Environ. Entomol. 25: 1190-1195.
- Tuomi, J., P. Niemela & R. Mannila. 1981. Leaves as islands: Interactions of Scolineura betuleti (Hymenoptera) miners in birch leaves. Oikos 37: 146-152.
- Vargas, H.C. 1970. Observaciones sobre la biología y enemigos naturales de la polilla del tomate, Gnorimoschema absoluta (Meyrick) (Lepidoptera: Gelechiidae). Idesia 1: 75-110.
- Zar, J.H. 1999. Biostatistical analysis. New Jersery, Prentice Hall (4th ed.), 663p.

Publication Dates
-
Publication in this collection
11 Dec 2006 -
Date of issue
Oct 2006
History
-
Accepted
02 Jan 2006 -
Received
20 June 2005