Resumos
O trabalho teve por objetivo determinar as exigências térmicas para o desenvolvimento e estimar o número de gerações anuais de Aedes aegypti (L.) em campo. O ciclo biológico das populações de A. aegypti foi estudado nas temperaturas constantes de 18, 22, 26, 30 e 34ºC, e fotofase de 12h. Foram avaliados, diariamente, o período de desenvolvimento e a viabilidade da fase de ovo, sobrevivência de larva e pupa, longevidade e a fecundidade dos adultos, determinando-se, ainda, os limites térmicos inferiores de desenvolvimento (Tb) e as constantes térmicas (K). Em função dessas variáveis foram estimados os números de gerações anuais do inseto em laboratório e campo. A temperatura favorável ao desenvolvimento de A. aegypti encontra-se entre 21ºC e 29ºC, e para a longevidade e fecundidade os adultos entre 22ºC e 30ºC. As Tb s, Ks de ovo a adulto, e o número de gerações anuais em campo foram de 9,5, 8,5, 3,4, 7,1, 13,5ºC; 244,5, 273,9, 298,5, 280,9 e 161,8 graus-dias; e 21,9, 23,8, 24,2, 21,1 e 22,1 gerações para as populações de A. aegypti das regiões de Boqueirão, Brejo dos Santos, Campina Grande, Itaporanga e Remígio, respectivamente.
Insecta; vetor; ciclo biológico; temperatura
The present work aimed at estimating the thermal requirements for the development and the number of generations per year of Aedes aegypti (L.) under natural conditions. The life cycle of A. aegypti populations was studied at constant temperatures of 18, 22, 26, 30 and 34ºC, and 12h photophase. The development period, egg viability and larval and pupal survival were evaluated daily as well as adult longevity and fecundity. The low threshold temperature of development (Tb) and the thermal constant (K) were determined. The number of generations per year in laboratory and field were also estimated. The favorable temperature to A. aegypti development is between 21ºC and 29ºC, and to longevity and fecundity is between 22ºC and 30ºC. The egg to adult basal temperature, thermal constant and the number of generations in field were, in order, 9.5, 8.5, 3.4, 7.1, 13.5ºC; 244.5, 273.9, 298.5, 280.9 and 161.8 degree-days; and 21.9, 23.8, 24.2, 21.1 and 22.1 generations in populations from Boqueirão, Brejo dos Santos, Campina Grande, Itaporanga and Remígio.
Insecta; vector; biological cycle; temperature
PUBLIC HEALTH
Biologia e exigências térmicas de Aedes aegypti (L.) (Diptera: Culicidae) provenientes de quatro regiões bioclimáticas da Paraíba
Biology and thermal exigency of Aedes aegypti (L.) (Diptera: Culicidae) from four bioclimatic localities of Paraíba
Eduardo B. BeserraI; Francisco P. de Castro Jr.II; José W. dos SantosIII; Tatiana da S. SantosIV; Carlos R.M. FernandesIV
IIGraduando em Ciências Biológicas, Bolsista PIBIC/CNPq. Depto. Biologia. Univ. Estadual da Paraíba-UEPB, Rua Juvêncio Arruda s/n, Bodocongó, 58109-753, Campina Grande, PB
IVEstatística, Bolsista do CNPq/EMBRAPA tatysilvasantos@gmail.com. Centro Nacional de Pesquisa de Algodão, Embrapa Algodão, Rua Oswaldo Cruz, 1143, Centenário, 58107-720, Campina Grande, PB. Mestrando do PRODEMA/UFPB/UEPB, Av. das Baraúnas, 35, Campus Universitário, Bodocongó, 58109-753 Campina Grande, PB
RESUMO
O trabalho teve por objetivo determinar as exigências térmicas para o desenvolvimento e estimar o número de gerações anuais de Aedes aegypti (L.) em campo. O ciclo biológico das populações de A. aegypti foi estudado nas temperaturas constantes de 18, 22, 26, 30 e 34ºC, e fotofase de 12h. Foram avaliados, diariamente, o período de desenvolvimento e a viabilidade da fase de ovo, sobrevivência de larva e pupa, longevidade e a fecundidade dos adultos, determinando-se, ainda, os limites térmicos inferiores de desenvolvimento (Tb) e as constantes térmicas (K). Em função dessas variáveis foram estimados os números de gerações anuais do inseto em laboratório e campo. A temperatura favorável ao desenvolvimento de A. aegypti encontra-se entre 21ºC e 29ºC, e para a longevidade e fecundidade os adultos entre 22ºC e 30ºC. As Tbs, Ks de ovo a adulto, e o número de gerações anuais em campo foram de 9,5, 8,5, 3,4, 7,1, 13,5ºC; 244,5, 273,9, 298,5, 280,9 e 161,8 graus-dias; e 21,9, 23,8, 24,2, 21,1 e 22,1 gerações para as populações de A. aegypti das regiões de Boqueirão, Brejo dos Santos, Campina Grande, Itaporanga e Remígio, respectivamente.
Palavras-chave: Insecta, vetor, ciclo biológico, temperatura
ABSTRACT
The present work aimed at estimating the thermal requirements for the development and the number of generations per year of Aedes aegypti (L.) under natural conditions. The life cycle of A. aegypti populations was studied at constant temperatures of 18, 22, 26, 30 and 34ºC, and 12h photophase. The development period, egg viability and larval and pupal survival were evaluated daily as well as adult longevity and fecundity. The low threshold temperature of development (Tb) and the thermal constant (K) were determined. The number of generations per year in laboratory and field were also estimated. The favorable temperature to A. aegypti development is between 21ºC and 29ºC, and to longevity and fecundity is between 22ºC and 30ºC. The egg to adult basal temperature, thermal constant and the number of generations in field were, in order, 9.5, 8.5, 3.4, 7.1, 13.5ºC; 244.5, 273.9, 298.5, 280.9 and 161.8 degree-days; and 21.9, 23.8, 24.2, 21.1 and 22.1 generations in populations from Boqueirão, Brejo dos Santos, Campina Grande, Itaporanga and Remígio.
Key words: Insecta, vector, biological cycle, temperature
Aedes (Stegomyia) aegypti (L.) é um importante vetor em saúde pública devido a seu papel como transmissor de dengue e de febre amarela. É um mosquito com ampla distribuição geográfica, predominando nas áreas tropicais e subtropicais situadas entre os paralelos de latitudes 45° Norte e 40° Sul (Forattini 2002) e nas zonas isotermais intermediadas a 20ºC (Gadelha 1985). Devido ao seu hábito antropofílico, A. aegypti apresenta grande capacidade de adaptação a criadouros artificiais o que possibilita o aumento de sua população e, por conseguinte, o aparecimento de epidemias de dengue (Lozovei 2001).
A temperatura é um dos principais fatores ecológicos que influi tanto direta como indiretamente sobre os insetos, seja no seu desenvolvimento, seja na sua alimentação (Silveira-Neto et al. 1976). Vários autores demonstraram os efeitos da temperatura sobre o ciclo de vida dos insetos e de como este fator ecológico pode ser utilizado para se entender a dinâmica populacional de vetores e com isto desenvolver estratégias adequadas para o seu controle. Costa et al. (1994) estudaram a influência de nove temperaturas que variaram entre 5°C e 45°C, sobre o ciclo aquático de Culex quinquefasciatus Say (Diptera: Culicidae) e verificaram que a faixa ótima para o desenvolvimento encontra-se entre 20°C e 30°C. Ribeiro et al. (2001) constataram que a 30°C, sob condições de laboratório, Ophyra aenescens Wiedemann (Diptera: Culicidae), apresenta 19,9 gerações por ano, com viabilidade total de 75,9%. Calado & Navarro-Silva (2002a) relataram que a atividade hematofágica e a oviposição de Aedes albopictus Skuse, sofrem influência da temperatura, e que as baixas temperaturas de 15°C e 20°C atuam como fator limitante ao crescimento populacional do inseto. Para esses autores, a eliminação dos criadouros artificiais nos meses mais frios, pode ser uma forma de diminuição da população de fêmeas aptas a realizar o repasto sangüíneo. Segundo Calado & Navarro-Silva (2002b) a manutenção de criadouros artificiais durante o período de temperaturas acima do limiar de desenvolvimento de A. albopictus favorecerá o aumento de sua população, o que deve ser considerado na definição das estratégias de controle.
Assim, deve-se considerar que o conhecimento sobre os efeitos da temperatura sobre o ciclo de vida, e o estudo das exigências térmicas das diferentes fases de desenvolvimento dos insetos é um importante elemento para a definição das estratégias de controle. Tais conhecimentos, quando aplicados a A. aegypti permitirão determinar o número de gerações anuais e a época favorável à ocorrência de altas densidades populacionais em campo, e assim auxiliar na adoção de medidas de monitoramento e controle. Este trabalho teve como objetivo verificar a influência da temperatura sobre o ciclo de vida, determinar as exigências térmicas para o desenvolvimento e o número de gerações anuais de A. aegypti.
Material e Métodos
A pesquisa foi conduzida com populações de A. aegypti coletadas em municípios pertencentes a quatro regiões bioclimáticas da Paraíba, a saber: 1 - Nordestino quente de seca atenuada com temperatura média anual em torno de 25ºC, médias pluviométricas entre 700 mm e 900 mm e umidade relativa do ar de 80% a 85%, com estação seca de quatro a cinco meses, compreendendo os municípios de Campina Grande (07º13'32"S 35º54'15"W) e Remígio (Latitude de 06º58'01"S 35º47'29"W); 2 - Nordestino quente de seca acentuada com temperatura média em torno de 23ºC, precipitação pluviométrica média anual entre 300 mm e 400 mm e umidade relativa do ar de 75%, com estação seca de sete a oito meses, compreendendo o município de Boqueirão (07º29'27"S 36º08'09"W); 3 - Tropical quente de seca média com temperatura média anual em torno de 25ºC, precipitação pluviométrica de 600 mm a 700 mm, umidade relativa do ar de 65%, com estação seca de cinco a sete meses compreendendo o município de Brejo dos Santos (06º22'48"S 37º49'29"W); 4 - Tropical quente de seca acentuada com temperatura média anual em torno de 25ºC, precipitação pluviométrica de 600 mm a 800 mm, umidade relativa do ar de 65%, com estação seca de sete a oito meses, compreendendo o município de Itaporanga (07º18'00"S 37º09'00"W) (Lima & Heckendorff 1985).
As colônias de A. aegypti e os bioensaios de laboratório foram conduzidos no Laboratório de Controle Biológico pertencente ao Núcleo de Manejo Integrado de Pragas da Universidade Estadual da Paraíba UEPB, em sala de criação climatizada a temperatura de 26ºC ± 2ºC e fotofase de 12h e câmaras climatizadas (B.O.D.). As larvas de A. aegypti foram criadas em bandejas plásticas (40 x 27 x 7,5 cm) cobertas por uma tela de malha fina, ofertando-se a elas, 1,0 mg/larva de ração para peixe ornamental (Alcon/Goldfish crescimento). Os adultos foram mantidos em gaiolas, construídas de armação de madeira e tecido tipo organza (40 x 40 x 20 cm), ofertando-se diariamente uma solução de mel a 20%. Às fêmeas, foi permitido o repasto sanguíneo, em codornas, durante 1h, três vezes por semana.
O ciclo biológico de cada população foi estudado em câmaras climatizadas e em salas de criação reguladas a 18, 22, 26, 30 e 34ºC, e fotofase de 12h, utilizando-se a primeira geração de laboratório. Cada câmara recebeu 12 copos de polietileno de 250 ml, contendo água desclorada e 10 larvas recém-eclodidas (L1) de A. aegypti. Diariamente aferiu-se a temperatura da água dos copos de desenvolvimento larval através de um termômetro químico com aferição até 60ºC. Os adultos foram mantidos em gaiolas de madeira telada (20 cm3), em no total de cinco gaiolas por tratamento, cada uma contendo vinte casais. A forma de alimentação das larvas e adultos seguiu aquela da metodologia de criação. Para a fase de ovo foram avaliadas para cada temperatura 20 posturas contendo em média 80 ovos, acondicionados em placas-de-petri de 9 cm de diâmetro x 1,5 cm de profundidade, contendo água desclorada suficiente para encobrir os ovos. Os ovos que não apresentaram eclosão de larvas após dez dias da oviposição foram considerados inviáveis. As avaliações foram diárias, registrando-se para cada temperatura, o período de desenvolvimento e a mortalidade das fases de larva, pupa e ovo. E para a fase adulta a emergência, a longevidade e a fecundidade.
O delineamento experimental foi inteiramente casualizado em esquema fatorial 5 x 4, cinco populações e quatro temperaturas, para as fases de ovo e de ovo a emergência do adulto, e de 5 x 5, cinco populações e cinco temperaturas, para as fases de larva e pupa e para a longevidade e fecundidade dos adultos, sendo as variáveis biológicas estudadas submetidas à análise de variância e as médias comparadas pelo teste de Tukey (P < 0,05),
Com base nas durações das fases imaturas de desenvolvimento e ciclo de ovo a adulto, foram determinados os limites térmicos inferiores de desenvolvimento (Tb) e as constantes térmicas (K), através do método da hipérbole (Haddad & Parra 1984) dado pela equação K= y (t a) onde: K a constante térmica expressa em graus-dia; y o tempo de desenvolvimento (dias); t a temperatura ambiente (ºC); a, a temperatura do limiar de desenvolvimento.
A partir dos dados biológicos obtidos na temperatura na qual o ciclo foi completado em menor tempo e em função da constante térmica do ciclo biológico e do total de graus-dia disponíveis para as populações no ano, foi estimado o número de gerações anuais da espécie em laboratório. O número provável de gerações anuais de A. aegypti em campo foi estimado com base em suas exigências térmicas, e nas normais térmicas das áreas de procedência das populações do vetor, obtidas do Laboratório de Meteorologia, Recursos Hídricos e Sensoriamento Remoto da Paraíba/ Universidade Federal de Campina Grande-UFCG, através da equação NG= [T (tc tb)/K], onde T é o tempo considerado em ano, e tc as temperaturas médias mensais das regiões de estudo, sendo K e tb a constante térmica e as temperaturas base definidas anteriormente.
Resultados e Discussão
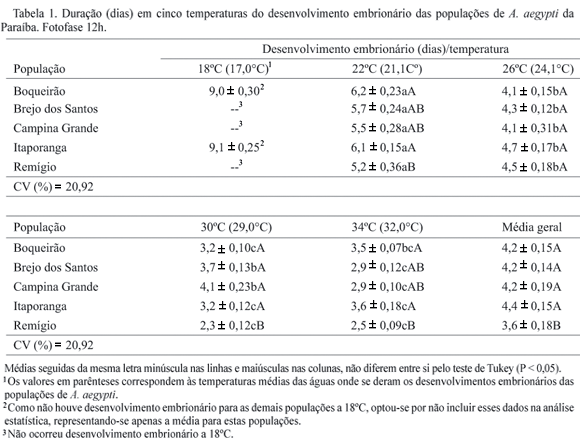
Ciclo de vida de A. aegypti em cinco temperaturas constantes. As temperaturas médias das águas em que se estudou o desenvolvimento do ovo à emergência dos adultos das populações de A. aegypti foram de aproximadamente 17ºC, 21ºC, 24ºC, 29ºC, e 32ºC para as temperaturas ambientes de 18ºC, 22ºC, 26ºC, 30ºC e 34ºC, respectivamente. Os resultados referentes ao período de desenvolvimento foram discutidos em relação à temperatura da água.
Ocorreram interações significativas entre população e temperatura com relação ao ciclo biológico do mosquito, indicando que as populações de A. aegypti se comportaram de forma diferente em relação à temperatura (Tabelas 1 e 2). Não ocorreu eclosão na temperatura da água de 17ºC, para as populações originárias de Brejo dos Santos, Campina Grande e Remígio, sendo que entre 21ºC e 32ºC o tempo de desenvolvimento reduziu à medida que a temperatura aumentou, com as menores médias registradas a 29°C e 32ºC. Em geral o menor período de desenvolvimento embrionário foi registrado para a população de Remígio com média de 3,62 dias (Tabela 1).
Não se avaliou se a não eclosão de larvas das populações de Brejo dos Santos, Campina Grande e Remígio a 17°C foi conseqüência da morte do embrião ou função da interrupção do desenvolvimento devido à baixa temperatura. É possível que o mosquito adaptado às temperaturas médias mensais das suas regiões de origem, que estão acima dos 20°C, não se desenvolva quando submetido a temperaturas mais baixas, como a 17°C da presente pesquisa. Assim Ribeiro et al. (2004) constataram desenvolvimento embrionário a 15°C em uma população de C. quinquefasciatus coletada em Pelotas, RS, onde as temperaturas mínimas estão abaixo dos 15°C durante o inverno, atribuindo tais aspectos à adaptação climática às condições ambientais daquela região.
Comparando-se as populações de A. aegypti entre as temperaturas de 21ºC e 32ºC, detectou-se que a menor viabilidade ocorreu em ovos da população de Campina Grande quando submetidos à temperatura da água de 32ºC, e foi de 33,3%, porém os ovos da população oriunda de Remígio foram os mais sensíveis aos efeitos da temperatura, apresentando viabilidade inferior a 70% em todas as temperaturas, com média geral de 61,7% (Tabela 2). Deve-se ressaltar que a viabilidade da fase de ovo observada na presente pesquisa para todas as populações pode ser considerada baixa se comparada às viabilidades dos ovos de A. aegypti relatada em outros estudos, como o de Silva & Silva (1999), que encontraram a porcentagem de eclosão de 85,4%.
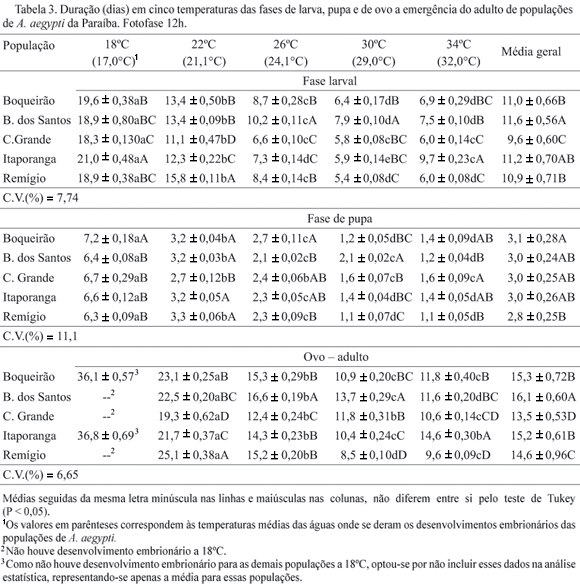
O período larval diminuiu com a elevação térmica, com a menor duração média registrada para a população de Campina Grande a 32°C que foi de 6,7 dias (Tabela 3). A maior sobrevivência da fase de larva foi observada para população de Brejo dos Santos a 21°C, em torno de 100% (Tabela 2). Durante a fase de pupa, o menor período de desenvolvimento foi constatado na população de A. aegypti de Remígio a 32°C, sendo o maior período observado para a população de Campina Grande a essa mesma temperatura (Tabela 3). Para a população de Campina Grande, também foi observada a menor sobrevivência pupal, de 54% a 17°C. Para as demais populações, a sobrevivência em todos os tratamentos foi superior a 90% nas fases de larva e pupa (Tabela 2), sendo que a população coletada em Remígio, apresentou 100% de sobrevivência pupal em todas as temperaturas.
Houve interação significativa entre temperatura e população de A. aegypti com relação à duração do período de desenvolvimento de ovo à emergência do adulto. A população de Campina Grande foi a menos afetada pela temperatura com desenvolvimento mais rápido na faixa de 21°C a 32ºC, com duração média variando de 19,4 a 10,6 dias. As populações de A. aegypti de Boqueirão e Remígio sofreram maior influência das temperaturas, principalmente a 21°C, onde os tempos de desenvolvimento de ovo a emergência do adulto, foram de 23,2 e 25,1 dias respectivamente (Tabela 3).
A longevidade dos adultos de A. aegypti dependeu da interação entre população e temperatura, sendo estas significativamente maiores a 18ºC, diminuindo nas temperaturas de 30ºC a 34ºC. Considerando-se a média geral das longevidades em todas as temperaturas, os adultos da população de A. aegypti de Remígio foram mais longevos que os adultos das demais populações estudadas (Tabela 4).
Os extremos de temperatura, 18ºC e 34ºC, foram altamente deletérios à fecundidade das fêmeas, diminuindo consideravelmente o número de ovos destas. O maior número médio de ovos/fêmeas foi obtido a 26ºC, com médias de 271,9 e 260,40 ovos para as populações de Campina Grande e Remígio respectivamente, porém sem diferir significativamente entre o número de ovos por fêmea das populações de Boqueirão e Itaporanga.
Considerando-se a média geral de ovos de todas as temperaturas, não ocorreram diferenças significativas entre as populações avaliadas (Tabela 4). A 18ºC, embora tenha aumentado a longevidade das fêmeas, o número de ovos, principalmente para a população de Campina Grande, foi consideravelmente baixo (Tabela 4), podendo-se inferir que para essa temperatura, parte da energia que seria empregada na reprodução, foi alocada para a sobrevivência do adulto. Isto significa que, como ocorre para outros insetos, em condições climáticas desfavoráveis, as fêmeas de A. aegypti diminuem a fecundidade para aumentar sua longevidade e com isto superar as condições adversas. Dados de Chadee (1997) mostram que, sob condições desfavoráveis, fêmeas de A. aegypti retêm os ovos e essa retenção pode resultar na reabsorção de folículos maduros e aumentar a sobrevivência (Judson 1968). Pode acontecer também a alocação de menos proteína do sangue ingerido para produção de ovos e utilização de carboidratos do sangue para manutenção de outras funções (Frankino & Juliano 1999), como, por exemplo, a sobrevivência.
Exigências térmicas e número de gerações anual de A. aegypti. Como não ocorreu desenvolvimento embrionário à temperatura da água de 17ºC (18ºC temperatura ambiente), para as populações de Brejo dos Santos, Campina Grande e Remígio, essa temperatura foi excluída dos cálculos da temperatura base (Tb) e constante térmica (K). Para esses cálculos foram utilizadas as temperaturas das águas para o ciclo de ovo a emergência do adulto a partir de 21°C. A velocidade de desenvolvimento de A. aegypti em função da temperatura ajustou-se ao modelo linear determinado através da recíproca da equação da hipérbole (Haddad & Parra 1984). As exigências térmicas variaram dependendo da população e do estágio de desenvolvimento do inseto (Tabela 5).
Os limites térmicos (Tb) de desenvolvimentos e as respectivas constantes térmicas (K), expressas em graus-dias (GD), para as fases de ovo, larva, pupa e de ovo a emergência do adulto variaram de acordo com as populações estudadas (Tabela 5). As menores temperaturas bases para as fases de ovo, larva, pupa e de ovo a emergência do adulto foram observadas para as populações de Boqueirão, Brejo dos Santos, Campina Grande e Itaporanga, sendo as maiores para A. aegypti de Remígio. Essas diferenças entre as populações podem ser conseqüências das adaptações ecológicas as suas regiões de origem, porém, não era de se esperar que as populações de Boqueirão, Brejo dos Santos e Itaporanga apresentassem menores temperaturas bases do que a população de Remígio, já que este município está localizado no Brejo paraibano, onde as temperaturas são mais amenas e, portanto, sua população teria maiores condições de resistirem às baixas temperaturas. Por outro lado os valores das temperaturas bases da fase de ovo, principalmente para as populações de Brejo dos Santos (Tb = 9,8°C), Campina Grande (Tb = 2,9°C) e Remígio (Tb = 13,9°C), foram muito baixos, considerando-se que para essas populações não ocorreu eclosão à temperatura da água de 17°C (Tabela 1).
As temperaturas bases e constantes térmicas diferiram de acordo com o estágio de desenvolvimento de cada população, sendo as menores temperaturas bases para as fases de ovo e larva (Tabela 5). Segundo Calado & Navarro-Silva (2002b), tais variações podem surgir como conseqüência de diferenças estruturais e fisiológicas inerentes a cada estágio, que favorecem a adaptação da espécie às variações do ambiente. Essas variações foram constatadas em várias outras espécies como O. aenescens (Ribeiro et al. 2001), A. albopictus (Calado & Navarro-Silva 2002b) e C. quinquefasciatus (Ribeiro et al. 2004). Os resultados referentes às temperaturas bases e constantes térmicas para as fases de ovo e larva das populações de A. aegypti estudadas diferiram dos observados para A. albopictus por Calado & Navarro-Silva (2002b), que foram de 9,07°C e 214,4 GD e de 11,95°C e 110,11GD (fêmea) e de 12,03°C e 99, 48 GD (macho), respectivamente, contudo para a fase de pupa esses autores constaram valores de 11,60°C e 35,30 GD para fêmea e 11,87°C e 32,40GD para macho, que foram muito próximos aos encontrados na presente pesquisa.
O limite térmico inferior de 2,9ºC observado para a fase de ovo da população de Campina Grande foi muito abaixo dos observados para A. aegypti provenientes de Boqueirão, Brejo dos Santos, Itaporanga e Remígio. Esse resultado não era esperado, mesmo considerando-se que essas populações ocorrem em condições climáticas diferentes, e principalmente porque, para a região de Campina Grande a menor temperatura média mensal está em torno dos 20ºC, com a mínima variando de 17,8ºC no mês de agosto a 20,4 ºC no mês de março, segundo dados do Laboratório de Meteorologia, Recursos Hídricos e Sensoriamento Remoto da Paraíba-UFCG. Para as populações estudadas, os tempos de desenvolvimentos embrionários, nas diferentes temperaturas, foram muito próximos e em geral não diferiram significativamente entre si (Tabela 1). Para as fases de larva e pupa os valores da temperatura base e da constante térmica não sofreram grandes variações entre elas (Tabela 5), levando a crer que a temperatura base para fase de ovo de Campina Grande deveria estar mais próxima daquelas encontradas para Boqueirão, Brejo dos Santos, Itaporanga e Remígio. A menor temperatura base da fase de ovo refletiu no limiar térmico de desenvolvimento de ovo a adulto da população de A. aegypti de Campina Grande, que foi de 3,43ºC, portanto considerado baixo se comparado àqueles calculados para as demais populações.
As constantes térmicas para o ciclo de ovo a adulto foram de 244,5, 273,9, 298,5, 280,9 e 161,8 graus-dias (GD), respectivamente para as populações de Boqueirão, Brejo dos Santos, Campina Grande, Itaporanga e Remígio. Tomando-se por base a temperatura de criação do inseto que é de 26ºC, o número de gerações anuais em condições de laboratório foi de 23,26, 24,63, 27,59, 24,56 e 28,22 para as populações de Boqueirão, Brejo dos Santos, Campina Grande, Itaporanga e Remígio respectivamente. Em condições de campo, o número de gerações anuais variou de 21,1 a 24,2 (Tabela 5), ou aproximadamente duas gerações do inseto por mês, o que também foi verificado para C. quiquefasciatus por Ribeiro et al. (2004).
As variações no ciclo de vida em função das temperaturas entre as populações de A. aegypti de Boqueirão, Brejo dos Santos, Campina Grande, Itaporanga e Remígio, provavelmente estão associadas às adaptações as condições climáticas das suas respectivas regiões de origem. Porém, para todas elas a faixa favorável ao desenvolvimento do vetor encontra-se entre 22°C e 30°C, portanto, dentro da faixa de temperatura das suas regiões de ocorrência. As temperaturas bases para o desenvolvimento das diferentes fases do ciclo de vida do inseto foram inferiores às temperaturas mínimas de cada região, demonstrando, que para essas populações, com relação à temperatura, todas as regiões apresentam condições favoráveis ao desenvolvimento e multiplicação do vetor ao longo do ano, e que esforços devem ser concentrados na eliminação dos criadouros artificiais, principalmente nas estações mais quentes e secas, quando a população procura acumular água em depósitos artificiais para enfrentar a estiagem.
A amplitude de temperatura favorável ao ciclo de vida das populações de A. aegypti em condições de laboratório encontra-se entre 22°C e 30°C, e que os extremos de temperatura de 18°C e 34°C apresentaram efeitos negativos sobre o desenvolvimento e a fecundidade do inseto, embora o limiar térmico de desenvolvimento de ovo a adulto, de 3,43°C e 13,5°C, esteja abaixo da temperatura mínima de 18°C. Baseado nos dados de constante térmica e nas normais climatológicas de cada região verificou-se que todos municípios apresentam condições favoráveis ao estabelecimento da população do vetor durante todo o ano, permitindo a ocorrência de mais de 20 gerações anuais de A. aegypti.
Agradecimentos
Agradecemos às Secretarias Municipais de Saúde de Boqueirão, Brejo dos Santos, Campina Grande, Itaporanga e Remígio, e ao senhor Eurivaldo dos Anjos, coordenador da vigilância epidemiológica do município de Campina Grande, pelas informações e apoio ao projeto. À Fundação de Amparo a Pesquisa do Estado da Paraíba, FAPESq, à Fundação Nacional de Saúde, FUNASA, e ao Ministério da Saúde, pelo apoio financeiro através do convênio FAPESq/FNS/MS, 1256/2001. Ao CNPq, pela concessão de bolsa de iniciação científica.
Received 04/III/06.
Accepted 07/VI/06.
- Calado, D.C. & M.A. Navarro-Silva. 2002a. Influência da temperatura sobre a longevidade, fecundidade e atividade hematofágica de Aedes (Stegomyia) albopictus Skuse, 1894 (Diptera, Culicidae) sob condições de laboratório. Rev. Bras. Entomol. 46: 93-98.
- Calado, D.C. & M.A. Navarro-Silva. 2002b. Exigências térmicas de Aedes (Stegomyia) albopictus Skuse, 1894 (Diptera, Culicidae) em condições de laboratório. Rev. Bras. Entomol. 46: 547-551.
- Chadee, D.D. 1997. Effects of forced egg-retention on the oviposition patterns of female Aedes aegypti (Diptera: Culicidae). Bull. Entomol. Res. 87: 649-651.
- Costa, P.R.P., E.E.S. Vianna; P. Silveira Jr & P.B. Ribeiro. 1994. Influência da temperatura na longevidade e viabilidade do ciclo aquático do Culex quinquefasciatus Say,1823 (Diptera: Culicidae) em condições de laboratório. Rev. Bras. Parasit. Vet. 3: 87-92.
- Forattini, O.P. 2002. Culicidologia médica. São Paulo, Edusp. 860p.
- Frankino, W.A. & S.A. Juliano. 1999. Costs of reproduction and geographic variation in the reproductive tactics of the mosquito Aedes triseriatus Oecologia 120: 59-68.
- Gadelha, D.T.A. 1985. Biologia e comportamento do Aedes aegypti Rev. Bras. Mal. Trop. 37: 29-36.
- Haddad, M.L. & J.R.P. Parra. 1984. Métodos para estimar os limites térmicos e a faixa ótima de desenvolvimento das diferentes fases do ciclo evolutivo dos insetos. Piracicaba, FEALQ, 12p.
- Judson, C.L. 1968. Physiology of feeding and oviposition behavior in Aedes aegypti (L.) experimental dissociation of feeding and oogenesis. J. Med. Entomol. 5: 21-23.
- Lima, P.J. & W.D. Heckendorff. 1985. Climatologia. In Atlas geográfico do estado da Paraíba. João Pessoa, Secretária de Educação do Estado. Universidade Federal da Paraíba UFPB, 100p.
- Lozovei, A.L. 2001. Culicideos (mosquitos), p.59-104. In C.B. Marcondes (org.), Entomologia médica e veterinária. 1Ş ed. São Paulo, Atheneu, 432p.
- Ribeiro, P.B., C.J.B. Carvalho, M. Regis & P.R.P. Costa. 2001. Exigências térmicas e estimativas do número de gerações de Ophyra aenescens Wiedemann,1830 (Diptera: Muscidae, Azeliinae), em Pelotas, RS. Arq. Inst. Biol. 68: 75-82.
- Ribeiro, P.B, P.R.P. Costa, A.E. Loeck, E.E.S. Viannna & P. Silveira Jr. 2004. Exigências térmicas de Culex quinquefasciatus (Diptera, Culicidae) em Pelotas, Rio Grande do Sul, Brasil. Iheringia. Ser. Zool. 94: 177-180.
- Silva, H.H.G. & I.G. Silva. 1999. Influência do período de quiescência dos ovos sobre o ciclo de vida de Aedes aegypti (Linnaeus, 1762) (Diptera, Culicidae) em condições de laboratório. Rev. Soc. Bras. Med. Trop. 32: 349-355.
- Silveira Neto, S., O. Nakano, D. Barbin & N. Villa Nova. 1976. Manual de ecologia dos insetos. São Paulo, Agronômica Ceres, 419p.
Datas de Publicação
-
Publicação nesta coleção
31 Jan 2007 -
Data do Fascículo
Dez 2006
Histórico
-
Aceito
07 Jun 2006 -
Recebido
04 Mar 2006