Abstracts
The multivariate allometric growth patterns from longitudinal data of leg and rostral segments of all instars of Belostoma elegans (Mayr) and B. cummingsi De Carlo are presented for the first time, and the allometric coefficients are compared. The segments of the middle and hind legs present allometric coefficients with opposite polarity to those of forelegs. This discrepancy in the ontogenetic trajectories may be due to the different functions of the legs. The foreleg main function is to capture of the prey, while the middle and hind legs are adapted to swimming. The relative size of rostral segments is an important taxonomic character in Belostoma. In B. cummingsi, the proximal segment is longer than the middle one, while in B. elegans they are subequal. Our purpose is to explain these differences through the analysis of their ontogenetic trajectories in an attempt to illuminate the morphological differences among species from a developmental perspective.
Nymph; postembryonic development; water bug; morphometry
Los patrones de crecimiento alométrico de los segmentos del rostro y de las patas de Belostoma elegans (Mayr) y B. cummingsi De Carlo son presentados y comparados por primera vez. Se emplearon datos longitudinales de todos los estadios ninfales en un contexto multivariado. Los segmentos de la pata media y posterior presentan coeficientes alométricos con polaridad opuesta a los de la pata anterior. Estas diferencias observadas pueden deberse al diferente rol que presentan las patas. La función principal de la pata anterior es la captura de las presas, mientras que las patas media y posterior están adaptadas para la natación. El tamaño relativo de los segmentos del rostro es una característica taxonómica importante en Belostoma. En B.cummingsi el segmento proximal es más largo que el medio, mientras que en B. elegans son subiguales. Nuestro propósito es explicar estas diferencias a través del análisis de las trayectorias ontogéneticas en un intento de aclarar diferencias morfológicas entre especies desde una perspectiva del desarrrollo.
Ninfa; desarrollo postembrionario; insecto acuático; morfometría
SYSTEMATICS, MORPHOLOGY AND PHYSIOLOGY
A longitudinal study of two species of Belostoma Latreille (Heteroptera: Belostomatidae): allometry and ontogeny
Estudio longitudinal en dos especies de Belostoma Latreille (Heteroptera: Belostomatidae): alometría y ontogenia
Mónica S. Iglesias, María S. Gaspe; Alejandra C. Valverde
Depto. de Biodiversidad y Biología Experimental, Facultad de Ciencias Exactas y Naturales, Univ. de Buenos Aires, Argentina; iglesias@bg.fcen.uba.ar, solgaspe@bg.fcen.uba.ar, valverde@bg.fcen.uba.ar
ABSTRACT
The multivariate allometric growth patterns from longitudinal data of leg and rostral segments of all instars of Belostoma elegans (Mayr) and B. cummingsi De Carlo are presented for the first time, and the allometric coefficients are compared. The segments of the middle and hind legs present allometric coefficients with opposite polarity to those of forelegs. This discrepancy in the ontogenetic trajectories may be due to the different functions of the legs. The foreleg main function is to capture of the prey, while the middle and hind legs are adapted to swimming. The relative size of rostral segments is an important taxonomic character in Belostoma. In B. cummingsi, the proximal segment is longer than the middle one, while in B. elegans they are subequal. Our purpose is to explain these differences through the analysis of their ontogenetic trajectories in an attempt to illuminate the morphological differences among species from a developmental perspective.
Key words: Nymph, postembryonic development, water bug, morphometry
RESUMEN
Los patrones de crecimiento alométrico de los segmentos del rostro y de las patas de Belostoma elegans (Mayr) y B. cummingsi De Carlo son presentados y comparados por primera vez. Se emplearon datos longitudinales de todos los estadios ninfales en un contexto multivariado. Los segmentos de la pata media y posterior presentan coeficientes alométricos con polaridad opuesta a los de la pata anterior. Estas diferencias observadas pueden deberse al diferente rol que presentan las patas. La función principal de la pata anterior es la captura de las presas, mientras que las patas media y posterior están adaptadas para la natación. El tamaño relativo de los segmentos del rostro es una característica taxonómica importante en Belostoma. En B.cummingsi el segmento proximal es más largo que el medio, mientras que en B. elegans son subiguales. Nuestro propósito es explicar estas diferencias a través del análisis de las trayectorias ontogéneticas en un intento de aclarar diferencias morfológicas entre especies desde una perspectiva del desarrrollo.
Palabras-clave: Ninfa, desarrollo postembrionario, insecto acuático, morfometría
There is a variety of ways to define allometry, but all agree that it deals with variation either between two traits within an organism or between one trait and the whole organism (Klingenberg 1998). Different aspects of allometry have been investigated, such as the mechanisms that generate the patterns of organ size relative to total body size in holometabolous insects (Stern & Emlen 1999) and the genetic analysis of size change during ageing (Cheverud et al. 1983).
Recently, Wu & Hou (2006) designed a hyperspace model to identify individual quantitative trait loci for phenotypic variation by integrating biologically meaningful mathematical functions of growth curves and developmental allometry. The covariation of traits may be analysed in three different ways. Static allometry reflects trait covariation among individuals in a particular instar; ontogenetic allometry examines growth relationships during development, and evolutionary or phylogenetic allometry inspects the size relationships of organisms among various species. In addition, these three levels are interrelated. Any evolutionary change in morphology is accompanied by a corresponding change in ontogeny (and vice versa), and evolutionary change depends on the heritable static variation of morphological traits in various life-history stages produced by ontogenetic variation (Klingenberg 1998).
Most studies on ontogenetic allometry employ cross-sectional data where each individual specimen is measured in a single instar, and only an average allometric trajectory can be obtained as a composite from many individuals (Wiig 1985, Iglesias & Crespo 2003, Masurani et al. 2005). Only in a few cases longitudinal data is applied, where each individual is measured multiple times during growth, thus allowing to assess the individual variability of allometric trajectories (Klingenberg 1996). Alberch (1985) recognized that a major challenge in systematics would be to incorporate ontogenetic trajectories into a methodological framework, but a lot remains to be done.
The allometric framework was applied in a variety of animals (Chabot & Stenson 2000, Palestrini et al. 2000, Weston 2003, Masunari et al. 2005), among which hemimetabolous insects turned to be an excellent model as involved structures remain inextensible throughout each instar (Klingenberg & Spence 1993, Tseng & Rowe 1999).
The Belostomatidae family includes nearly 150 species of aquatic insects distributed almost all over the world. Interest in these heteropterous grew when the sanitary importance of this group as biological control agents of mosquito larvae and gastropods (Bionphalaria spp.), some of them vectors of human diseases, became evident (Cônsoli et al. 1989, Pereira e Melo 1998, Aditya et al. 2005). In the Belostoma Latreille genus, certain topics such as geographic distribution (Morrone et al. 2004), reproductive ecology and life history (Schnack et al. 1989, Valverde & Shang 1994, Armúa de Reyes & Estévez 1997), interspecific and intraespecific competition (Schnack et al. 1980, Schnack et al. 1981), food preference and behaviour (Armúa de Reyes & Estévez 2006, Amarilla & Armúa de Reyes 2004) have been widely investigated, but there are no previous reports on the ontogenetic allometry.
Belostoma elegans (Mayr) and B. cummingsi De Carlo are usually found both in temporary and permanent lentic environments and in low volume streams, with profuse aquatic vegetation. There is at present no consensus on the behaviour of B. elegans as an " r" or " k" strategist. It shows great aptitude for flight, extensive geographical distribution and high fertility while in stable environments (Domizi et al. 1978, Schnack et al. 1987, Armúa de Reyes & Kehr 1999). More recently, Perez Goodwyn (2001) stated that it does not require that type of habitat to survive, inferring that B. elegans is more probably an " r" strategist. In this context, no reports are available on B. cummingsi, but we have captured adults using light traps, which may indicate dispersion by flight and therefore a similar ecological strategy. It is relevant to study the ecology and behavior in the framework of the allometric growth patterns to test adaptive hypothesis.
Belostoma species are arranged in subgroups on the basis of morphological traits (Lauck 1962), and investigations on the ontogenetic allometry are likely to provide new data for the discussion on the monophylia of the subgroups proposed by this author.
The ontogenetic trajectories of B. elegans and B. cummingsi based on a multivariate longitudinal data set are presented and compared. Further work on the Belostoma genus and other belostomatids might provide a clearly understanding of the general trends of growth in this family in order to devise hypothesis on evolutive models of growth
Material and Methods
This study is based on longitudinal data from all instars (N1-N5) of B. elegans and B. cummingsi. Exuviae collected from individually reared bugs allowed to obtain measurements from single individuals in all growth instars.
Study organism. The 70 species approximately described in the genus Belostoma (Nieser 1975, Estévez & Polhemus 2001) are found mainly in the American neotropics. B. elegans is widely spread in South America, where its southernmost limit is the Province of Río Negro, Argentina. B. cummingsi is distributed in Brazil, Paraguay, Argentina and Uruguay (Morrone et al. 2004).
Belostoma spp. are excellent models for allometry research due to their particular reproductive behaviour, which involves female oviposition on male backs. Consequently, following the capture of males carrying tenths of eggs, it is possible to obtain a complete life cycle in laboratory and to accurately identify juvenile instars, which can be individually reared. Measurements of the cuticle cast after moulting provide an accurate record of growth, being each moult a distinct developmental event.
Laboratory rearing and measurements. Specimens used in this study were the offspring of a sample of adults captured with an insect water net of fine mesh. B. elegans was collected in the Luján River and B. cummingsi in the city area of Tigre, both in the Province of Buenos Aires, Argentina.
Laboratory rearing was performed under environmental temperature and natural light regime. The younger nymphs were fed with Tubifex sp. (Oligochaeta: Tubificidae), and the older nymphs and adults with larvae of Ulomoides dermestoides (Fairmaire) (Coleoptera: Tenebrionidae). All specimens were periodically fed ad libitum. Male adults with their eggs were kept in containers with abundant vegetal support until larvae hatched. Hatchings were transferred into individual containers with water and vegetal support. After each moult, exuviae were collected and stored in 70% ethanol, and measurements were made using a Leica stereomicroscope with a graduate ocular. The data set includes only those individuals for which all variables could be measured in all five nymphal instars. Morphometric data of total length without head (TL), maximum width (MW), and the length of the femur (Fe I, Fe II, Fe III), tibiae (Ti I, Ti II, Ti III), and rostrum (R1, R2, R3) of B. elegans (n = 50) were taken. TL, MW, R1 and R2 of B. cummingsi (n = 50) were measured (Fig.1). The head was excluded from total length as it increased the measurement error by a positional effect.
Multivariate statistics. The principal components analysis (PCA) was applied to data corresponding to all five instars (N1-N5). This method finds hypothetical variables (components) that account for as much of the variance as possible, according to the available data set. These new variables are linear combinations of the original ones. The analysis was performed with data of leg and rostrum segments of B. elegans. The procedure was repeated using the following measurements: TL, MW, R1, and R2 of B. elegans and B. cummingsi. The PCA loading for each variable and the percentage of the variability explained by the most important components were obtained. PC1 vs. PC2 was plotted and results for both species were compared.
A multivariate allometry method applied to a data set based on Jolicoeur (1963) with extensions by Kowalewski et al. (1997) was used. Ninety five percent confidence intervals for the allometric coefficients were estimated by bootstrapping, and 2000 bootstrap replicates were made. The morphometric data of rostral and leg segments of B. elegans, and TL, MW, R1 and R2 of both species were analysed and compared.
Results and Discussion
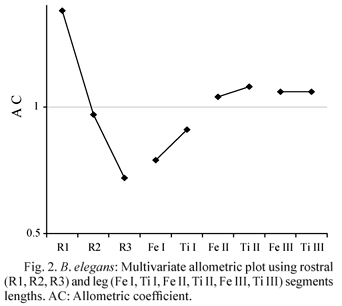
The multivariate allometric analysis of rostral and leg segments of B. elegans (Table 1) shows the existence of a decreasing proximo-distal gradient of allometric coefficients for the beak. R1 presents a clearly positive allometry; the hypothesis of isometry cannot be rejected for R2, and the apical segment shows hypoallometry. A similar growth trend was found in the antennae of other groups of aquatic heteropterous as a common pattern (Matsuda 1960, Iglesias & Crespo 2003). Further research might reveal if this decreasing proximo-distal gradient is also a generalised pattern for the rostrum of Belostoma.
The femur and tibiae of middle and hind legs present a moderate positive allometry, while forelegs evidenced negative allometric growth (Fig. 2). The main function of forelegs is to capture and restrain the prey, while middle and hind legs participate in swimming. Legs are likely to be subjected to different selection pressures during ontogeny. Allometric patterns are informative in themselves, allowing to recognize developmental variation and to predict differences in the functionality of structures.
The PCA plot (Figs. 3-4) clearly exhibits five areas in the morphospace that correspond to the five instars of both species. In the case of field-captured specimens, multivariate studies might allow a prompt identification of the instar with only a few measurements. The correct instar determination in Belostoma is crucial to understand the population structure in a specific region and to quantify its alimentary impact at a certain moment.

The first principal component accounts for 98% of the variance, revealing an excellent fit of the simple ontogenetic allometry model to the analyzed data. These results have also been obtained in different multivariate ontogenetic studies in other insects (Davies & Brown 1972, Klingenberg 1996) and animals (Wiig 1985).
The relative contribution of each segment to the total length of the rostrum in B. elegans differs in each instar (Fig. 5). R2 is relatively stable along the postembryonic development, whereas R1 contributes 26% to the total length of the rostrum in N1, and 38% in N5. The apical segment grows more slowly than the rostrum as a whole, and its final contribution is proportionally lower than the original one. It represents 33% of the rostrum total length in N1, but only 23% in N5.
When comparing B. elegans and B. cummingsi using TL, MW, R1 and R2, more than 98.9% of the total variance in each species is accounted for by the PC1s. The PCA loadings for both species were very similar (Table 2).
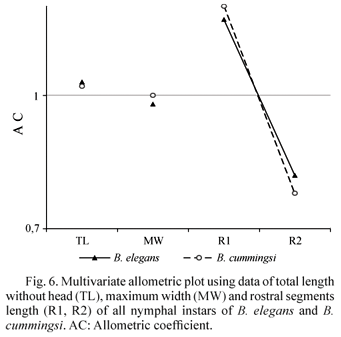
The relative length of rostral segments varies within Belostoma and it is a trait of considerable systematic importance. In B. elegans, R1 and R2 are subequal in adults as a diagnostic character, while in B. cummingsi, the second segment is shorter than the proximal one.
Comparing the multivariate allometric analysis for both species (Table 3; Fig. 6) it may be inferred that the allometric growth coefficients of B. elegans are slightly lower for R1 and higher for R2. The proximal segment of B. elegans responds to hyperallometry and the second segment to hypoallometry. Nonetheless, in the case of B. elegans, segments reach length balance only in the adult stage, while in B. cummingsi allometric growth rates are more divergent. Therefore, R1 in the latter outgrows R2 in size during N5, emphasizing this difference in adults. Our results reveal that minor allometric ontogenetic changes in different structures can cause significant divergences in the final stage.
Acknowledgments
This work was funded by the Secretaría de Ciencia y Técnica, Universidad de Buenos Aires.
Received 19/II/07. Accepted 13/X/08.
- Aditya, G., S. Bhattacharyya, N. Kundu & G.K. Saha. 2005. Frequency - dependent prey - selection of predacious water bugs on Armigeres subalbatus immatures. J. Vect. Borne Dis. 42: 9-14.
- Alberch, P. 1985. Problems with the interpretation of developmental sequences. Syst. Zool. 34: 46-58.
- Amarilla, L.R. & A.C. Armúa de Reyes. 2004. Estudio de la preferencia alimentaria de Belostoma elegans (Heteroptera: Belostomatidae) y su importancia sanitaria. Comunicaciones Científicas y Tecnológicas 2004. UNNE, 2p.
- Armúa de Reyes, A.C. & A.I. Kehr. 1999. Ciclo de vida y aspectos reproductivos de una población de Belostoma elegans (Heteroptera: Belostomatidae) en un ambiente permanente de Corrientes, Argentina. Reunión de Comunicaciones Científicas y Tecnológicas (UNNE, Corrientes) 6.21-6.24.
- Armúa de Reyes, A.C. & A.L. Estévez. 1997. Observaciones biológicas de los estados preimaginales de Belostoma elegans Mayr en condiciones controladas. FACENA 13: 141-143.
- Armúa de Reyes, A.C. & A.L. Estévez. 2006. Predation on Biomphalaria sp. (Mollusca: Planorbidae) by three species of the genus Belostoma (Heteroptera: Belostomatidae). Braz. J. Biol. 66: 1033-1035.
- Chabot, D. & G.B. Stenson. 2000. Implantation date, growth rate, and allometric relationships in foetal Northwest Atlantic harp seals (Phoca groenlandica). Can. J. Zool/Rev. Can. Zool. 78: 501-505.
- Cheverud, J.M., J. Rutledge & W. Atchley. 1983. Quantitative genetics of development: Genetic correlations among age-specific trait values and the evolution of ontogeny. Evolution 37: 895-905.
- Cônsoli, R.A.G.B., M.H. Pereira, A.L. Melo & L.H. Pereira. 1989. Belostoma micantulum Stal (Hemiptera: Belostomatidae) as a predator of larvae and pupae of Aedes fluviatilis (Diptera: Culicidae) in laboratory conditions. Mem. Inst. Oswaldo Cruz 84: 577-578.
- Davies, R.G. & V. Brown. 1972. A multivariate analysis of postembryonic growth in two species of Ectobius (Dyctioptera: Battidae). J. Zool. 168: 51-79.
- Domizi, E.A., A.L. Estévez, J.A. Schnack & G.R. Spinelli. 1978. Ecología y estrategia de una población de Belostoma oxyurum (Dufour) (Hemiptera: Belostomatidae). ECOSUR 10: 157-168.
- Estévez, A.L. & J.T. Polhemus. 2001. The small species of Belostoma (Hemiptera: Belostomatidae). I. Introduction, Key to species groups and a revision of the denticolle group. Inheringia 91: 151-158.
- Iglesias, M.S. & F.A. Crespo. 2003. Crecimiento relativo en Brachymetra albinerva albinerva (Amyot & Serville, 1843) (Heteroptera: Gerridae). Physis (Buenos Aires). Secc. B. 60: 5-10.
- Jolicoeur, P. 1963. The multivariate generalization of the allometry equation. Biometrics 19: 497-499.
- Klingenberg, C.P. 1996. Individual variation of ontogenies: A longitudinal study of growth and timing. Evolution 50: 2412-2428.
- Klingenberg, C.P. 1998. Heterochrony and allometry: The analysis of evolutionary change in ontogeny. Biol.Rev. 73: 79-123.
- Klingenberg, C.P. & J.R. Spence. 1993. Heterochrony and allometry: Lessons from the water strider genus Limnoporus Evolution 47: 1834-1853.
- Kowalewski, M., E. Dyreson, J.D. Marcot, J.A. Vargas, K.W. Flessa & D.P. Hallmann. 1997. Phenetic discrimination of biometric simpletons: Paleobiological implications of morphospecies in the lingulide brachiopod Glottidia Paleobiology 23: 444-469.
- Lauck, D.R. 1962. A monograph of the genus Belostoma (Hemiptera). Part I. Introduction and Belostoma dentatum and subspinosum groups. Bull. Chicago Acad. Sci. 11: 34-81.
- Masurani, S., N. Disshena & R.C. Falcao. 2005. Crescimento relativo e destreza dos quelipodos de Uca aracoani (Latreille) (Crustacea: Decapoda: Ocypodidae) no Baixio Mirim, Baia de Guaratuba, Parana, Brasil. Rev. Bras. Zool. 22: 974-983.
- Matsuda, R. 1960. Morphology, evolution and classification of the Gerridae (Hemiptera: Heteroptera). Univ. Kansas. Sci. Bull. 41: 25-632.
- Morrone, J.J., S.A. Mazzucconi & A.O. Bachmann. 2004. Distributional patterns of chacoan water bugs (Heteroptera: Belostomatidae, Corixidae, Micronectidae and Gerridae). Hydrobiología 523: 159-173.
- Nieser, N. 1975. The water bugs (Heteroptera: Nepomorpha) of the Guyana region. Studies on the fauna of Suriname and other Guyanas 16: 1-308.
- Palestrini, C., A. Rolando & P. Laiolo. 2000. Allometric relationships and character evolution in Onthophagus taurus (Coleoptera: Scarabeidae). Can. J. Zool/Rev. Can. Zool. 78: 1199-1206.
- Pereira, M.H. & A.L. Melo. 1998. Influência do tipo de presa no desenvolvimento e na preferência alimentar de Belostoma anurum Herrich-Schäffer 1948 e Belostoma plebejum (Stal, 1858) (Heteroptera: Belostomatidae). Ecología de insectos aquaticos. Series Oecol. Brasiliensis 5: 41-49.
- Perez Goodwyn, P.J. 2001. Muscle regression on Belostomatid bugs. Aquat. Insects 23: 23-32.
- Schnack, J.A., E.A. Domizi, A.L. Estévez & G.R. Spinelli. 1980. Determinantes ecológicos de la competencia sexual en Belostomatinae. Consideraciones sobre una población de Belostoma oxyurum (Dufour) (Hemiptera: Belostomatidae). ECOSUR 7: 1-13.
- Schnack, J.A., E.A. Domizi, A.L. Estévez & G.R. Spinelli. 1989. Ecología del comportamiento reproductivo de Belostomatinae. I. Sistemas de apareamiento en poblaciones del área platense de Belostoma oxyurum,B. elegans y B. micantulum (Hemiptera: Belostomatidae). Rvta. Soc. Ent. Argent. 45: 85-94.
- Schnack, J.A., E.A. Domizi, G.R. Spinelli & A.L. Estévez. 1981. Influencia de la densidad sobre la fecundidad y competencia interespecífica con referencia especial a una población de Belostomatidae. (Insecta: Hemiptera). Limnobios 2: 239-246.
- Schnack, J.A., G.R. Spinelli, A.L. Estévez & E.A. Domizi. 1987. Ciclo anual y estructura de edades de Belostoma micantulum (Stal) y B. elegans (Mayr), con referencia a dos poblaciones de la provincia de Buenos Aires. (Hemiptera: Belostomatidae). Rev. del Museo de La Plata (N.S.), Sección Zool. 14: 83-91.
- Stern, D.L. & D.J. Emlen. 1999. The developmental basis for allometry in insects. Development 126:1091-1101.
- Tseng, M. & L. Rowe. 1999. Sexual dimorphism and allometry in the giant water strider Gigantometra gigas Can. J. Zool. 77: 923-929.
- Valverde, A.C. & M.M. Schang. 1994. Caracterización de los estadios larvales de Belostoma cummingsi De Carlo (Heteroptera: Belostomatidae). Rev. Soc. Entomol. Argent. 53: 109-119.
- Weston, E.M. 2003. Evolution of ontogeny in the hippopotamus skull: Using allometry to dissect developmental change. The Linnean Society of London. Biol. J. Linn. Soc. 80: 625-638.
- Wiig, O. 1985. Morphometric variation in the hooded seal (Cystophora cristata). J. Zool. 206: 497-508.
- Wu, R. & W. Hou. 2006. A hyperspace model to decipher the genetic architecture of developmental processes: Allometry meets ontogeny. Genetics 172: 627-637.
Publication Dates
-
Publication in this collection
16 Jan 2009 -
Date of issue
Dec 2008
History
-
Received
19 Feb 2007 -
Accepted
13 Oct 2008