Resumo
Although Merostachys fischeriana is very abundant in the Brazilian Atlantic Rainforest, little attention has been paid to the biological interactions with other animals. The present study describes some of the interactions between ants and this bamboo species. The experiment was carried out in a fragment of a montane tropical forest in the Parque Estadual do Itacolomi, near Ouro Preto, MG, Brazil. Thirty culms of bamboo were randomly collected. The ants were obtained by direct collection from nodes and internodes. Morphometric variables of the bamboo were recorded for characterization of potential ant habitat. Merostachys fischeriana grows in rosettes as a thin bamboo (average = 1,0 cm; se = 0,27; n = 20) and is tall enough to reach the upper canopy of this low forest (average = 9,1 m; se = 2,72; n = 20). Fifteen ant species were sampled. Brachymyrmex heeri Forel was the most abundant in the nodes, while Camponotus crassus Mayr (Hymenoptera: Formicidae) was the most abundant in the internodes. The composition of the species that inhabit the internodes was different from the composition in the node (Q-test: Q = 3,76; P = 0,05). The level of occupation was defined by the number of holes (F = 10,33; P < 0,01), the number of internodes in the canopy (F = 6,84; P = 0,01) and the length of the culm (F = 7,52; P = 0,01). The plant's morphology allowed the occurrence of additional species of ants in the canopy and influenced the composition of the entire ant assemblage.
Brachymyrmex; Camponotus; canopy habitat enrichment; ant nesting behavior; ant-plant interaction
Brachymyrmex; Camponotus; canopy habitat enrichment; ant nesting behavior; ant-plant interaction
ECOLOGY, BEHAVIOR AND BIONOMICS
O bambu Merostachys fischeriana (Bambusoideae: Bambuseae) como habitat para formigas de Floresta Tropical Montana
The bamboo Merostachys fischeriana (Bambusoideae: Bambuseae) as a canopy habitat for ants of Neotropical Montane Forest
Roberth FagundesI; Gilberto TerraII; Sérvio P. RibeiroI; Jonathan D. MajerIII
ILab de Ecologia Evolutiva de Insetos de Dossel e Sucessão Natural, DEBIO/ICEB/Univ Federal de Ouro Preto, Ouro Preto, MG, Brasil; roberthbio@yahoo.com.br; spibeiro@iceb.ufop.br
IIUniv Federal de São Carlos, São Carlos, SP, Brasil; terra@bol.com.br
IIICentre for Ecosystem Diversity and Dynamics in the Dept of Environmental and Aquatic Sciences, Curtin Univ of Technology, Perth, Australia; j.majer@curtin.edu.au
ABSTRACT
Although Merostachys fischeriana is very abundant in the Brazilian Atlantic Rainforest, little attention has been paid to the biological interactions with other animals. The present study describes some of the interactions between ants and this bamboo species. The experiment was carried out in a fragment of a montane tropical forest in the Parque Estadual do Itacolomi, near Ouro Preto, MG, Brazil. Thirty culms of bamboo were randomly collected. The ants were obtained by direct collection from nodes and internodes. Morphometric variables of the bamboo were recorded for characterization of potential ant habitat. Merostachys fischeriana grows in rosettes as a thin bamboo (average = 1,0 cm; se = 0,27; n = 20) and is tall enough to reach the upper canopy of this low forest (average = 9,1 m; se = 2,72; n = 20). Fifteen ant species were sampled. Brachymyrmex heeri Forel was the most abundant in the nodes, while Camponotus crassus Mayr (Hymenoptera: Formicidae) was the most abundant in the internodes. The composition of the species that inhabit the internodes was different from the composition in the node (Q-test: Q = 3,76; P = 0,05). The level of occupation was defined by the number of holes (F = 10,33; P < 0,01), the number of internodes in the canopy (F = 6,84; P = 0,01) and the length of the culm (F = 7,52; P = 0,01). The plant's morphology allowed the occurrence of additional species of ants in the canopy and influenced the composition of the entire ant assemblage.
Key words:Brachymyrmex, Camponotus, canopy habitat enrichment, ant nesting behavior, ant-plant interaction
A relação entre formigas e plantas superiores é diversa e muitas vezes de dependência (Huxley et al 1991). Dispersoras de sementes, polinizadoras, visitantes de nectários florais e extraflorais, habitantes, forrageadoras, seja qual for a relação existente, as formigas são os insetos não estritamente herbívoros mais comumente encontrados em plantas (Davidson et al 2003).
Outra característica peculiar das formigas é a grande diversidade de locais de nidificação. Seus ninhos variam de galerias simples no solo e cavidades de plantas até ninhos construídos com seda ou papel e suspensos em árvores (Hölldobler & Wilson 1990). A plasticidade em lidar com microhabitats, devido a sua heterogeneidade morfológica, permite que as espécies de formigas ocupem cavidades preexistentes nos troncos das árvores, galhos ocos e mortos, cavidades no interior do súber e ainda no interior de colmos de bambus (Hölldobler & Wilson 1990). Em contrapartida, as plantas evoluíram diversas estruturas para usufruírem a presença das formigas (Dorow & Maschwitz 1990).
Os bambus, ou taquaras, compreendem as espécies pertencentes à subfamília Bambusoideae (Poaceae). São conhecidas 1.200 espécies que se distribuem desde os 51º N até os 47º S, e desde o nível do mar até os 4.300 m de altitude. O Brasil possui a maior riqueza de bambus do Novo Mundo (Judziewicz et al 1999). Entre os 18 gêneros brasileiros, seis são endêmicos, e das 160 espécies, 129 (83%) são endêmicas. Os gêneros mais ricos são Merostachys (53 espécies), seguido por Chusquea (40 espécies) (Filgueiras & Santos-Gonçalves 2004). Esses dois gêneros constituem os bambus mais comuns da Mata Atlântica, sendo particularmente abundantes em matas secundárias onde desempenham papel de plantas pioneiras no processo de sucessão ecológica (Oliveira-Filho et al 1994).
Os bambus podem ser encarados como um ramificado conjunto de câmaras vazias e potencialmente colonizáveis. Os bambus da Australasia tropical são habitados por vários taxa de formigas, muitas delas especialistas, que habitam o interior dos entrenós dos colmos (Kohout 1988, Dorow & Maschwitz 1990). Entretanto as relações interespecíficas envolvendo bambus tratam normalmente de pássaros (Kratter 1998) e mamíferos (Silveira 2001). Os registros de interações entre invertebrados e bambus são escassos, com exceção dealguns estudos efetuados na Ásia (Maschwitz & Moog 2000) e outros na Região Neotropical (Louton 1996, Davidson et al 2003, Kondo & Gullan 2004).
As formigas são insetos de grande relevância ecológica para a Região Neotropical (Davidson et al 2003, Davidson 2005). Particularmente, há pouca informação sobre a ocupação e manipulação de microhabitats por formicídeos (Kaspari & Weiser 2000). Por outro lado, bambus arbóreos podem contribuir com novos habitats de dossel, o que é um aspecto negligenciado pela visão de que o bambu tem papel negativo na dinâmica sucessional da floresta (Touyama et al 1998).
O presente estudo pretende evidenciar a existência da interação formiga-bambu para uma floresta tropical montana e caracterizá-la no que diz respeito à estrutura do bambuzal e à assembléia de formigas associadas. De posse disso, propõe-se testar as hipóteses de que os colmos ocos do bambu Merostachys fischeriana fornecem um sítio de nidificação para formigas no interior das matas e que a distribuição das formigas ao longo dos colmos é determinada por caracteres morfológicos do próprio bambu e sua disposição no habitat.
Material e Métodos
Área de estudo. O Parque Estadual do Itacolomi (PEIT), abrangendo 7.000 ha, localiza-se no extremo sul da Cadeia do Espinhaço (20º22'30" e 20º30'00" S, 43º32'30" e 43º22'30" W), mais precisamente nos municípios de Ouro Preto e Mariana, MG. A precipitação varia de 1.000 mm a 1.500 mm anuais e a temperatura oscila entre 19ºC e 22ºC (Fujaco 2007).
No PEIT os Campos Rupestres dividem espaço com a Floresta Estacional Semidecidual Montana e Floresta Tropical Pluvial Montana (Rizzini 1979). A segunda é uma formação da Mata Atlântica localizada entre 800 m e 1.700 m, cobrindo morros e serras. As árvores atingem 20 m a 30 m de altura, porém são muitas vezes contorcidas e esguias.
No local de estudo, além da destacada presença de M. fischeriana, ocorre uma segunda espécie de Merostachys, duas espécies do gênero Chusquea e diversas espécies arbustivas e arbóreas das famílias Myrtaceae, Euphorbiaceae, Melastomataceae, Lauraceae, Rutaceae e Fabaceae, entre outras, que compõem o sub-bosque e o dossel da mata.
O bambu M. fischeriana. O estudo foi realizado com a espécie M. fischeriana. Esse bambu ocorre em manchas ascendendo até as copas das árvores. Seus colmos são finos e numerosos se abrindo em rosetas ao mesmo tempo em que se apóiam nas árvores. Essa espécie, além de ser muito comum no PEIT, é endêmica do Brasil e pertence ao gênero mais abundante na Mata Atlântica, sendo muito usado em programas de reflorestamento.
Merostachys fischeriana é encontrada tanto nas bordas quanto no interior de matas (Filgueiras & Santos-Gonçalves 2004). Seus colmos são grandes e com numerosos nós com gemas desenvolvidas em densas ramificações subiguais, típicas do gênero (Sendulski 2001), que envolvem os nós e permitem enovelamento e fixação dos colmos entre si e com o dossel sobrejacente. Apesar de verticais na base, os colmos assumem morfologia arqueada ao ascenderem às copas das árvores. O colmo é oco, apesar de fino, e apresenta inúmeros acessos ao seu interior, tanto por buracos como por rachaduras.
Amostragem de formigas e medidas arquitetônicas. O estudo foi realizado em junho e julho de 2006. Trinta colmos adultos do bambu M. fischeriana foram amostrados aleatoriamente dentro de um trecho de mata. Os colmos amostrados encontravam-se distantes entre si em no mínimo cinco metros. Foram considerados passíveis de coleta apenas os bambus adultos, que já apresentavam ramificações, vivos e íntegros, ou seja, sem quebra na parte mediana.
Cada colmo foi aberto ao meio sobre um guarda-chuva entomológico. As formigas que caíram no dispositivo foram recolhidas em saco plástico. Os sacos foram etiquetados e lacrados. Os nós foram tratados com poda dos ramos foliares e abertos ao meio para coleta dos indivíduos que se encontravam no seu interior.
As formigas foram congeladas e posteriormente acondicionadas em frascos contendo etanol 70%. Após montados, os indivíduos foram separados em morfoespécies e identificados até gênero com o auxílio da chave de identificação de Bolton (1994) e espécie/morfoespécie por comparação com a coleção entomológica do Laboratório de Ecologia Evolutiva de Insetos de Dossel e Sucessão Natural da Universidade Federal de Ouro Preto (UFOP), em Ouro Preto. Essa coleção compreende um conjunto de exemplares de Formicidae cujas espécies foram identificadas no Laboratório de Mirmecologia (CEPEC/CEPLAC), em Ilhéus, BA.
Do montante inicial de 30 colmos, foram registrados dados estruturais e morfológicos de 20 deles. Foram tomadas as seguintes medidas: número de entrenós, comprimento e diâmetro dos entrenós, número de furos, comprimento total do colmo e a altura em que ele atinge o dossel. A região do colmo onde ocorre a primeira ramificação também foi verificada, assim como o número de furos e a ocorrência ou não de formigas para cada entrenó. Para medir o comprimento acima do dossel, foi considerado que o mesmo se iniciaria acima da primeira bifurcação da árvore na qual o colmo se apoiava. O efeito das variáveis morfométricas sobre a distribuição da assembléia de formigas foi analisado com uso de modelos de regressão linear simples.
As espécies foram discriminadas quanto ao seu local de ocorrência no colmo: nó ou entrenó. Esses dados foram usados para comparar os dois locais de modo a testar a singularidade de cada local em relação à assembléia de formigas. O nível de diferença das duas assembléias foi submetido ao teste χ2, quanto a número de colmos ocupados, e ao teste Q, quanto a composição de espécies, para confirmação do observado. Todos os testes estatísticos foram realizados com o auxílio do programa BioEstat 5.0.
Resultados
Os colmos de M. fischeriana apresentaram em média 24,0 ± 7,39 entrenós por colmo (n = 20) distribuídos em seus 9,1 ± 2,72 m de comprimento (n = 20). Do comprimento total, aproximadamente 5,1 ± 1,57 m (n = 20) foram encontrados dentro do volume ocupado pelas copas das árvores. Os nós eram ramificados por folhas compridas e finas que ocorriam em toda extensão do colmo. Os entrenós possuíam em média 39,4 ± 5,56 cm de comprimento (n = 20) e 1,0 ± 0,28 cm de diâmetro (n = 20). Os entrenós foram praticamente iguais em seus diâmetros exceto pelos três ou quatro entrenós apicais que eram muito finos, medindo 0,3 ± 0,15 cm (n = 20), mas esse caráter não se mostrou significativamente determinante para a nidificação e formigas (veja abaixo).
Dentre os colmos analisados, 46% se encontravam lascados ou furados. A maior parte dessas aberturas foi registrada no terço médio dos colmos, em diversas posições e localizações nos entrenós e, em menor número, nos nós. Havia diversos tipos de perfurações, de diversas formas e tamanhos. A abertura externa é maior do que a abertura interna (para o interior do entrenó), de formato arredondado a irregular, com diâmetro de 3 mm, indicando que foram feitas de fora para dentro. As perfurações dispunham-se longitudinalmente no entrenó e a sua origem não foi determinada.
Para as perfurações encontradas nas regiões nodais, ou muito próximo a elas, um coleóptero provavelmente é o organismo perfurador, pois para alguns nós, principalmente aqueles nos quais havia formigas, foram encontradas larvas dessa ordem e suas respectivas galerias. As perfurações nos nós eram arredondadas e de menores dimensões que aquelas dos entrenós. Em alguns dos nós as galerias faziam uma conexão entre a região nodal e o interior do entrenó. Estas galerias também ocorreram em todos os casos nos quais foram encontradas formigas no interior do entrenó, sugerindo ser esta uma das formas de colonização da região interna.
Foram encontradas formigas tanto nos nós como nos entrenós do bambu. Encontramos 15 espécies de formigas incluídas em nove gêneros e quatro subfamílias. Os 30 nós ocupados abrigavam cinco espécies, enquanto que nos 33 entrenós ocupados foram coletadas 12 espécies, sendo apenas duas delas comuns aos dois microhabitats (Tabela 1). O teste χ2 revelou que o número de nós ocupados foi igual ao número de entrenós ocupados. Porém, a composição das espécies que habitava o nó foi diferente daquela que habitava os entrenós (teste-Q: Q = 3.77; P = 0.05). Os entrenós apresentaram um número maior de espécies (teste χ2: χ2 = 46,12; P < 0,0001).
Além da diferença na composição de espécies, cada guilda revelou peculiaridades na estrutura do ninho e colônia. Os ninhos de Brachymyrmex heeri Forel foram mais comuns nos nós do bambu. Essa espécie nidificou na base das ramificações foliares dos nós. Seus ninhos eram estruturados em sedimentos arenosos e detritos orgânicos acumulados no emaranhado de caulículos das folhas (Fig 1). Em todas as 30 colônias de B. heeri foi encontrada uma única rainha, sugerindo que essas sejam monogínicas, ao contrário dos ninhos das espécies de entrenó, nos quais quase não foram encontradas rainhas, exceto por Solenopsis sp1 e Linepithema humile Mayr. Os ninhos de entrenós eram caracterizados apenas pela ocupação de operárias, muitas vezes com larvas e pupas. Não foi encontrada qualquer estrutura interna elaborada, exceto para Camponotus rufipes Fabricius que construiu paredes laminares formadas pelo mesmo material cartonado que elas tipicamente usam para construção de seu ninho de solo. Aparentemente, o número de operárias foi menor para as formigas maiores, como Pachycondyla villosa autor. Enquanto Solenopsis sp1 apresentou uma colônia completa, para P. villosa foram encontradas apenas algumas operárias e algumas pupas, provavelmente um possível ninho satélite.
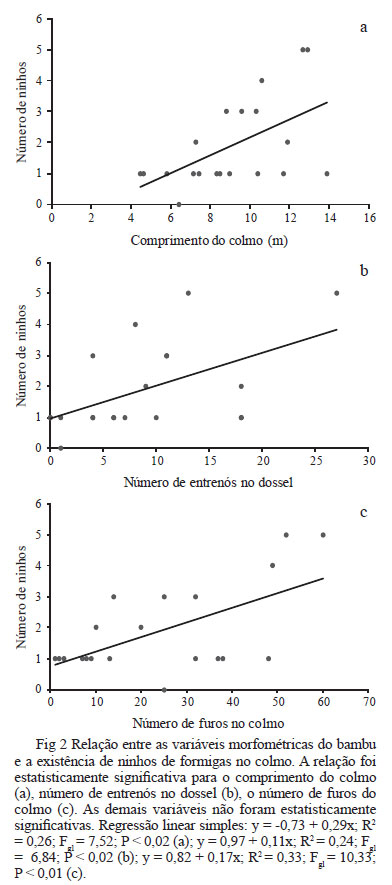
A partir dos dados morfométricos do bambu foi possível determinar que apenas o número de furos no colmo (Regressão Linear Simples: y = 0,82 + 0,17x; R2 = 0,33; Fgl = 10,33; P < 0,01), o número de entrenós no dossel (regressão linear simples: y = 0,97 + 0,11x; R2 = 0,24; Fgl= 6,84; P < 0,02) e o comprimento do colmo (regressão linear simples: y = -0,73 + 0,29x; R2 = 0,26; Fgl = 7,52; P < 0,02) estiveram relacionados com o número de colônias no bambu (Fig 2). Assim, quanto maior, mais perfurado e mais intrincado no dossel da mata estava o colmo de M. fischeriana, mais formigas foram encontradas em seus nós e entrenós. As demais variáveis não foram estatisticamente significativas.
Discussão
A interação entre bambu e formigas é um assunto ainda mal explorado (Kohout 1988, Dorow & Maschwitz 1990, Buschinger et al 1994, Maschwitz & Moog 2000). Emborahaja alguns registros dessa interação na Ásia (Maschwitz & Moog 2000), para as florestas neotropicais há registro de formigas associadas apenas a bambus do gênero Guadua (Davidson et al 2006). Essas formigas são especialistas em nidificar dentro de entrenós dos bambus, estejam eles vivos ou mortos (Davidson et al 2006). Porém, nenhum registro, até este estudo, havia sido feito para florestas montanas e bambus finos como M. fischeriana.
Davidson et al (2006) listam diversas espécies de formigas de bambu em seu estudo na Amazônia. Os autores registraram 62,7% das ocupações dos entrenós por Camponotus, demonstrando seu grande sucesso nesse habitat. Cobb et al (2006) introduziram entrenós de bambus para inventariar formigas nos cacaueiros da Bahia e encontraram muitas das espécies aqui relatadas nidificando em bambus naturais. A despeito dessa composição diversificada da mirmecofauna de bambu, Davidson et al (2006) consideraram muitas espécies ocupantes como oportunistas, quando observadas em bambus mortos, mas isso não é o caso dos colmos vivos. Entre essas espécies incluem aquelas dos grupos Ectatomma, Wasmannia e Solenopsis. Segundo Espírito-Santo (2008), L. humile e C. rufipes são numericamente dominantes no dossel da mata estudada.
Apesar das tentativas de descrever a interação em si, pouco se investigou sobre o grau de especialização existente dentro da assembléia. Davidson et al (2006) abordaram o fato de os bambus, apesar de mirmecófilos, não possuírem qualquer atrativo especial para formigas, o que indicaria uma associação unidirecional para as mesmas. Isso é evidente em algumas especializações morfológicas por parte das formigas, tais como a cabeça mais alongada, delgada e retangular de algumas espécies asiáticas, como Camponotus longipilis Emery, C. mirabilis Emery e C. depressus Mayr (Dorow 1996), que facilitaria a entrada no bambu por buracos pré-existentes. Tais buracos, em nosso estudo, foram determinantes na ocupação das formigas. Entretanto, Davidson et al (2006) discutiram que espécies especialistas em bambus são capazes de perfurar as paredes dos entrenós, exceto quando há aumento na lignificação da planta hospedeira. Esse comportamento de perfuração é semelhante ao encontrado por Gonsales (2002) para formigas que habitam a mirmecófita Cecropia purpurascens (Cecropiacea). As formigas ocupam o interior do caule da planta e alimentam-se de corpúsculos müllerianos ricos em glicogênio, produzidos nas axilas das folhas, denominados triquílias. Porém, nada disso se aplicaria às formigas de nós, pois seus ninhos são internos ao bambu e são dependentes de acúmulo de sedimentos na base dos ramos de folhas.
Comportamentos característicos de nidificação, como os observados para B. heeri, também foram registrados para C. mirabilis (Davidson et al 2006), Polyrhachis arachne Emery e Polyrhachis hodgsoni Forel (Dorow & Maschwitz 1990). Estas últimas são trofobiontes em uma associação com hemípteros (coccídeos e membracídeos) que vivem no interior dos ninhos de entrenó alimentando-se da seiva do bambu (Dorow & Maschwitz 1990, Dorow 1996). Para as formigas não-trofobiontes, Davidson et al (2006) defendem que os insetos de serrapilheira seriam a fonte nutricional. A ocorrência de trofobioses internas a ninhos de entrenós não foi verificada nesse trabalho.
Touyama et al (1998) registraram redução na riqueza em florestas invadidas por bambus. Entretanto, o bambu M. fischeriana mostra-se um importante componente para as florestas pluviais montanas, trazendo maior heterogeneidade de habitats e, consequentemente, mais espécies, interações e funções. Em comparação com Espírito-Santo (2008), a maioria das espécies encontradas em bambu não ocorre na floresta circunvizinha. Assim, o bambu cria condições e recursos compatíveis com o nicho de diversas espécies que dificilmente ocupariam a floresta ou o fariam em menor densidade.
A existência de uma assembléia de formigas estruturada em uma guilda funcional de habitantes de bambus indica o quão singular podem ser as Matas Montanas. A acumulação de 15 espécies utilizando especificamente 30 colmos de bambus arbóreos nesta comunidade aponta para a importância do estudo de micro-habitats. A riqueza encontrada para bambus corresponde a 30,5% da riqueza encontrada por Espírito-Santo (2008) para o dossel da mata estudada. Além disso, exceto por C. rufipes, nenhuma outra espécie coletada no bambu foi também coletada pela autora.
A relação entre a alta taxa de ocupação, o pequeno diâmetro dos entrenós e a proximidade com as copas já foi observado por Cobb et al (2006). O pequeno diâmetro dos canudos poderia favorecer a instalação de formigas por facilitar tanto a defesa do ninho quanto a manutenção das condições adequadas ao desenvolvimento sadio da colônia. O fato de os bambus serem mais ocupados quando próximos da copa indicaria a baixa disponibilidade de sítios de nidificação para as formigas nesse tipo de habitat. Nas copas, as formigas abrem mão de buracos pré-existentes ou lascas do próprio tronco para a nidificação. Cavidades como os colmos do bambu parecem ser uma alternativa. Além disso, no caso de formigas que utilizam exudatos de hemípteros, a colonização dos bambus no dossel, ainda que por ninhos satélites, permitiria maior proximidade com as agregações desses sugadores, reduzindo o gasto energético com o forrageio. Assim como demonstrado por Cobb et al (2006), este trabalho encontrou que dentre todas as características mensuradas do bambu, a altura e diâmetro do entrenó e a presença de furos são determinantes para a sua ocupação por formigas. Logo, necessita-se de estudos mais detalhados sobre como cada uma dessas características realmente afeta a ocupação, distribuição e composição da mirmecofauna do bambu.
Cabe salientar que o fato de os colmos atingirem o dossel sugere que há uma conexão direta entre solo e copa das árvores. Isso pode favorecer as espécies de artrópodes que habitam os colmos, pois para elas seria uma via de acesso a vários pontos no interior do dossel, já que os colmos permeiam a camada formada pelas copas das árvores. Esses fatos caracterizam M. fischeriana como provável via de acesso ao teto da floresta e afirma sua contribuição para a complexidade do dossel per se. Porém, estudos específicos são necessários, pois essa via também pode servir para predadores, influenciando negativamente a mirmecofauna.
O bambu M. fischeriana, por atingir o dossel superior a partir de um crescimento muito mais rápido que o das árvores, e ao mesmo tempo também mais efêmero, é um novo componente de habitat, renovável e dinâmico. Apesar de sua complexidade (com colmos, entrenós e nós de dimensões variadas), esse bambu cria condições para o estabelecimento de colônias de formigas com clara dominância e, ao mesmo tempo, partição do recurso. Desta forma, uma organização em mosaico vertical em escala pequena pode estar se formando dentro desse componente particular da floresta, contrapondose às pressões competitivas e às dominâncias principais existentes nas copas das árvores.
Espírito-Santo (2008) mostrou que as formigas dessas florestas são tipicamente dominadas por Myrmelachista catharinae Mayr e C. rufipes, que aparentemente não exercem pressão sobre as formigas dentro dos bambus, ao contrário do que ocorre fora deles. Consequentemente, o bambu torna-se espécie-chave na estruturação de toda uma comunidade de insetos que se relaciona com ele direta ou indiretamente, aumentando a diversidade alfa a partir de uma clara partição de recursos que resulta em compactação de espécies no espaço. Para os bambus, não há nenhum registro de interações com insetos para o gênero Merostachys, cuja distribuição geográfica abrange grande parte do Sul e Sudeste do Brasil (Sendulski 1995, Judziewicz et al 1999). Considerando que esse gênero é o de maior riqueza dentre todos os gêneros de bambus arborescentes no Brasil (Filgueiras & Santos-Gonçalves 2004), essas relações podem se estender a áreas bem maiores e para ambientes bem distintos daqueles do Parque Estadual do Itacolomi.
Agradecimentos
Ao professor Jacques H C Delabie pela literatura gentilmente fornecida. Ao suporte financeiro das patrocinadoras do IV Curso de Ecologia de Dossel: Conselho Nacional de Desenvolvimento Científico e Tecnológico - CNPq, Fundação de Amparo a Pesquisa de Minas Gerais - FAPEMIG, Parque Estadual do Itacolomi/Instituto Estadual de Florestas de Minas Gerais - PEIT/IEF, Global Canopy Programme, Embaixada Britânica no Brasil, Universidade Federal de Ouro Preto - UFOP e Ministério do Meio Ambiente - MMA.
Received 09/IV/09.
Accepted 08/VIII/10.
Edited by Paulo R Pereira - EMBRAPA
- Bolton B (1994) Identification guide to the ant genera of the world. Cambridge, Harvard University Press, 232p.
- Buschinger A, Klein R W, Maschwitz U (1994) Colony structure of a bamboo-dwelling Tetraponera sp. (Hymenoptera: Formicidae: Pseudomyrmecinae) from Malaysia. Insect Soc 41: 29-41.
- Cobb M, Watkins K, Silva E N, Nascimento I C, Delabie J H C (2006) An exploratory study on the use of bamboo pieces for trapping entire colonies of arboreal ants (Hymenoptera: Formicidae). Sociobiology 47: 215-223.
- Davidson D W (2005) Ecological stoichiometry of ants in a New World rain forest. Oecologia 142: 221-231.
- Davidson D W, Arias J A, Mann J (2006) An experimental study of bamboo ants in western Amazonia. Insects Soc 53: 108-114.
- Davidson D W, Cook S C, Snelling R R, Chua T H (2003) Explaining the abundance of ants in lowland tropical rainforest canopies. Science 300: 969-972.
- Dorow W H O (1996) Polyrhachis (Myrmhopla) schellerichae n. sp., a new ant of the hector-group from the Malay Peninsula. Sencken-berg Biol 76: 121-127.
- Dorow W H O, Maschwitz U (1990) The arachne-group of Polyrhachis (Formicidae, Formicinae): weaver ants cultivating Homoptera on bamboo. Insect Soc 37: 73-89.
- Espírito-Santo N B (2008) Assembléia de formigas do Parque Estadual do Itacolomi (MG) e relações intra e interespecíficas entre espécies dominantes. Dissertação de mestrado, Juiz de Fora, ICB/UFJF, 86p.
- Filgueiras T S, Santos-Gonçalves A P (2004) A checklist of the basal grasses and bamboos in Brazil (Poaceae). Sci Cult 18: 7-18.
- Fujaco M A G (2007) Influência dos diferentes tipos de substrato e geomorfologia na distribuição espacial e arquitetônica do gênero Eremanthus sp. no Parque Estadual do Itacolomi. Dissertação de mestrado, Ouro Preto, DEGEO/UFOP, 101p.
- Gonsales E M L (2002) Controle da estrutura de colônias de formigas Azteca alfari (Hymenoptera, Formicidae) pela mirmecófita Cecropia purpurascens (Cecropiacea). In Jansen Z, Venticinque E Curso de campo ecologia da floresta amazônica - 2002. Manaus, INPA, 122p.
- Hölldobler B, Wilson E O (1990) The ants. Cambridge, Belknap Press, 746p.
- Huxley C R, Cutler D F (1991) Ant-plant interactions. Oxford, Oxford University Press, 601p.
- Judziewicz E J, Clark L G, Londoño X, Stern M J (1999) American bamboos. Washington, Smithsonian Institution Press, 392p.
- Kaspari M, Weiser M D (2000) Ant activity along moisture gradients in a Neotropical forest. Biotropica 32: 703-711.
- Kohout R J (1988) A new species of Polyrhachis from Papua New-Guinea with a review of the New Guinean and Australian species (Hymenoptera: Formicidae: Formicinae). Mem Queensl Mus 25: 417-428.
- Kondo T, Gullan P J (2004) A new species of ant-tended soft scale of the genus Cryptostigma Ferris (Hemiptera: Coccidae) associated with bamboo in Peru. Neotrop Entomol 33: 717-723.
- Kratter A W (1998) The nests of two bamboo specialists: Celeus spectabilis and Cercomacra manu J Fie Ornit 69: 37-44.
- Louton J (1996) The aquatic macrofauna of water-filled bamboo (Poaceae: Bambusoideae: Guadua) internodes in a Peruvian lowland tropical forest. Biotropica 28: 228-242.
- Maschwitz U, Moog J (2000) Communal peeing: a new mode of flood control in ants. Naturwissenschaften 87: 563-565.
- Oliveira-Filho A T, Vilela E A, Gavilanes M L L, Carvalho D A (1994) Effect of floding regime and understorey bambos on the physiognomy and trees species composition of a tropical semideciduous forest in Southeastern Brazil. Vegetatio 113: 99-124.
- Rizzini C T (1979) Tratado de fitogeografia do Brasil: aspectos sociológicos e florísticos. São Paulo, EDUSP e Editora HUCITEC, 747p.
- Sendulski T (1995) Merostachys multiramea (Poaceae: Bambusoideae) and similar species from Brazil. Novon 5: 76-96.
- Silveira M (2001) A floresta aberta com bambu no sudoeste da Amazônia: padrões e processos em múltiplas escalas. Tese de doutorado, Brasília, UNB, 123p.
- Touyama Y, Yamamoto T, Nakagoshi N (1998) Myrmecofaunal change with bamboo invasion into broadleaf forests. J For Res 3: 155-159.
Datas de Publicação
-
Publicação nesta coleção
24 Jan 2011 -
Data do Fascículo
Dez 2010
Histórico
-
Recebido
09 Abr 2009 -
Aceito
08 Ago 2010