Abstracts
From September, 92 to August, 93 bee sampling was done in a secondary forest near the Pepital River, in Alcântara, MA, in order to study the local Euglossini fauna. Five aromatic compounds were used: eucaliptol, eugenol, methyl salicylate, vanillin, and benzoate. Four hundred sixty-seven male Euglossini bees were captured, distributed in 4 genus and 19 species. Euglossa was the most abundant and with high diversity (302 specimens and 14 species), followed by Eulaema (121; 3), Eufriesea (41; 1), and Exaerete (3; 1). The species which more frequently visited the bait were Euglossa piliventris (141 specimens; 30.19%), Euglossa cingulata (113; 24.21%), Euglossa ignita (45; 9.64%), Eufriesea pulchra (41; 8.78%), and Euglossa gaianii (33; 7.07%) corresponding to 79.88% of the sampling universe. The bees were active throught the year, however during the rainy season more activity and diversity were observed. The most attractive essence was eucaliptol (44.32% specimens and 84.21% species). In spite of this study having been done in a forest fragment, a secondary vegetation area smaller than other areas studied in Maranhão, it showed a significant diversity rate. This result reinforces the importance of fragments in the conservation of local bee communities.
Euglossini; diversity; abundance; seasonality; chemical baits
De setembro/92 a agosto/93, foram feitas coletas em uma área de vegetação secundária, próxima ao rio Pepital, Alcântara, MA, com o objetivo de conhecer a fauna de Euglossini local. Foram coletados 467 machos, 4 gêneros e 19 espécies. Euglossa foi o gênero mais abundante e com maior número de espécies (302 indivíduos, 14 espécies), seguido de Eulaema (121; 3), Eufriesea (41; 1) e Exaerete (3; 1). As espécies que mais se destacaram em visita às iscas-odores foram: Euglossa piliventris (141 ind.; 30,19% do total de abelhas coletadas), Eulaema cingulata (113; 24,21%), Euglossa ignita (45; 9,64%), Eufriesea pulchra (41; 8,78%) e Euglossa gaianii (33; 7,07%) representando 79,88% do universo amostral. Foram ativas durante todo o ano, entretanto, foram observadas maior atividade e maior diversidade de abelhas em visita às essências no período chuvoso. A essência que atraiu o maior número de espécies e indivíduos foi o Eucaliptol (84,21% e 44,32%, respectivamente). O sítio de estudos, embora apresentasse vegetação secundária, cuja área amostrada foi menor do que outras áreas já estudadas no Maranhão, revelou uma significativa diversidade de Euglossini. Tal resultado reforça a importância dos fragmentos na manutenção das comunidades de abelhas na região.
Euglossini; diversidade; abundância; sazonalidade; iscas-odores
COMMUNITY OF MALE EUGLOSSINI BEES (HYMENOPTERA: APIDAE) IN A SECONDARY FOREST, ALCÂNTARA, MA, BRAZIL
BRITO, C. M. S. de and RÊGO, M. M. C.
Departamento de Biologia, Universidade Federal do Maranhão, Campus do Bacanga, Av. dos Portugueses s/n, CEP 65080-040, São Luís, MA, Brazil
Correspondence to: Marcia Maria Corrêa Rêgo, Departamento de Biologia, Universidade Federal do Maranhão, Campus do Bacanga, Av. dos Portugueses s/n, CEP 65080-040, São Luís, MA,Brazil, e-mail: mmcrego@uol.com.br
Received May 22, 2000 Accepted August 18, 2000 Distributed November 30, 2001
(With 1 figure)
ABSTRACT
From September, 92 to August, 93 bee sampling was done in a secondary forest near the Pepital River, in Alcântara, MA, in order to study the local Euglossini fauna. Five aromatic compounds were used: eucaliptol, eugenol, methyl salicylate, vanillin, and benzoate. Four hundred sixty-seven male Euglossini bees were captured, distributed in 4 genus and 19 species. Euglossa was the most abundant and with high diversity (302 specimens and 14 species), followed by Eulaema (121; 3), Eufriesea (41; 1), and Exaerete (3; 1). The species which more frequently visited the bait were Euglossa piliventris (141 specimens; 30.19%), Euglossa cingulata (113; 24.21%), Euglossa ignita (45; 9.64%), Eufriesea pulchra (41; 8.78%), and Euglossa gaianii (33; 7.07%) corresponding to 79.88% of the sampling universe. The bees were active throught the year, however during the rainy season more activity and diversity were observed. The most attractive essence was eucaliptol (44.32% specimens and 84.21% species). In spite of this study having been done in a forest fragment, a secondary vegetation area smaller than other areas studied in Maranhão, it showed a significant diversity rate. This result reinforces the importance of fragments in the conservation of local bee communities.
Key words: Euglossini, diversity, abundance, seasonality, chemical baits.
RESUMO
Comunidade de machos de Euglossini (Hymenoptera: Apidae) em floresta secundária, Alcântara, MA, Brasil
De setembro/92 a agosto/93, foram feitas coletas em uma área de vegetação secundária, próxima ao rio Pepital, Alcântara, MA, com o objetivo de conhecer a fauna de Euglossini local. Foram coletados 467 machos, 4 gêneros e 19 espécies. Euglossa foi o gênero mais abundante e com maior número de espécies (302 indivíduos, 14 espécies), seguido de Eulaema (121; 3), Eufriesea (41; 1) e Exaerete (3; 1). As espécies que mais se destacaram em visita às iscas-odores foram: Euglossa piliventris (141 ind.; 30,19% do total de abelhas coletadas), Eulaema cingulata (113; 24,21%), Euglossa ignita (45; 9,64%), Eufriesea pulchra (41; 8,78%) e Euglossa gaianii (33; 7,07%) representando 79,88% do universo amostral. Foram ativas durante todo o ano, entretanto, foram observadas maior atividade e maior diversidade de abelhas em visita às essências no período chuvoso. A essência que atraiu o maior número de espécies e indivíduos foi o Eucaliptol (84,21% e 44,32%, respectivamente). O sítio de estudos, embora apresentasse vegetação secundária, cuja área amostrada foi menor do que outras áreas já estudadas no Maranhão, revelou uma significativa diversidade de Euglossini. Tal resultado reforça a importância dos fragmentos na manutenção das comunidades de abelhas na região.
Palavras-chave: Euglossini, diversidade, abundância, sazonalidade, iscas-odores.
INTRODUCTION
Euglossine bees (Apidae: Euglossini) are an important exclusively neotropical pollinator group that are most closely related with certain angiosperm families. Without specifically trying to collect pollen, they are able to secure pollination of species of Caesalpiniaceae, Maranthaceae, Euphorbiaceae, Araceae, Gesneriaceae, Bignoniaceae, Orchidaceae and others (Armbruster & Webster, 1979; Williams & Whitthen, 1983).
Male euglossine bees are the only pollinators of some orchids, e.g. Gongora, Castasctum, and Coryanthes. They are attracted to the essences produced by these orchids' flowers. The collection of these essences is by special pads on the tarsi of the forelegs and storage is in the pockets in the hind tibia (Bennett, 1972).
The use of these essences is not so well understood. However, Dodson (1970), Williams (1982), and Williams & Whitthen (1983) report that they can be used as a sexual attractant.
These chemical compounds are synthesized and can be used in ecological studies about the euglossine population, which has been done by Braga (1976), Janzen et al. (1982), Ackerman (1983, 1989), Pearson & Dressler (1985), Powell & Powell (1987), Roubik & Ackerman (1987), Raw (1989), Armbruster & McCormick (1990), Oliveira & Campos (1995, 1996), Oliveira (1999), Rebelo & Cabral (1997), Rebelo & Garófalo (1991, 1997), and Silva & Rebelo (1999).
Janzen (1971) reports that the euglossine sampling in a single area is representative for that area, habitat, and period. However, Becker et al. (1991) verified that abundance of euglossini males can vary in small intervals (300-700 m) so a sampling in one site cannot be regarded as representative of all the species of the region.
Although Maranhão State presents wide diversity of natural plant formations (Brasil, 1991), concern for the composition of euglossini community only began in the last decade with the work of Gomes (1991) and Fernandes (1991) in São Luís (north of the state), Silva & Rebelo (1999) in the pre-Amazonian region (west of the state), and Rebelo & Cabral (1997) in the wet littoral of Maranhão. Most of the state continues to have unknown euglossine fauna, in spite of accelerated devastation of the natural environment.
MATERIAL AND METHODS
Study site
The study site is located on the west littoral of the Maranhão State, on the west side of São Marcos Bay, between coordinates 2°23'00''S and 44°25'00''W.
The sample site is 25 km away from Alcântara city, along the Pepital River. The weather is warm and humid with an average temperature of around 25°C (annual variation between 0.8 and 3.2°C) and pluviometric index between 1,000 and 1,800 mm (Feitosa, 1983). Medium (8-10 m) and small trees compose the vegetation. The other side of the Pepital River has a plane area saturated during the rainy period and with occurrence of Mauritia flexuosa (buriti), Euterpe oleracea (juçara), Clusia sp. e Toccoca sp.
Sample
The survey was done from September 1992 to August 1993, in intervals of 28 and 30 days. The sampling activities were performed in two consecutive days, the first from 12:00 to 6:00 p.m. and the second from 6:00 a.m. to 12:00 p.m.
Five aromatic compounds were utilized to attract the male euglossine bees: eucaliptol, eugenol, methyl salicylate, vanillin, and benzil benzoate in cotton balls attached to twine.
The chemical baits were put at an average height of 2.0 m, at an average distance of 7 m. Every two hours, the baits were filled with the respective essence. The baits were observed every hour, but to avoid over-sampling the collector remained at each essence bait for only 12 minutes per observation. The bees were captured with an entomological net, after visiting the baits and killed in a jar with ethyl acetate (C4H8)2). They were kept in plastic bags with identification (capture hour and bait). They are now preserved in the entomological collection of Maranhão Federal University.
The temperature, relative humidity, and rain fall data were obtained in the Metereologic Station of Ponta da Madeira Terminal CVRD (São Luís), 20 km from the sample site.
RESULTS AND DISCUSSION
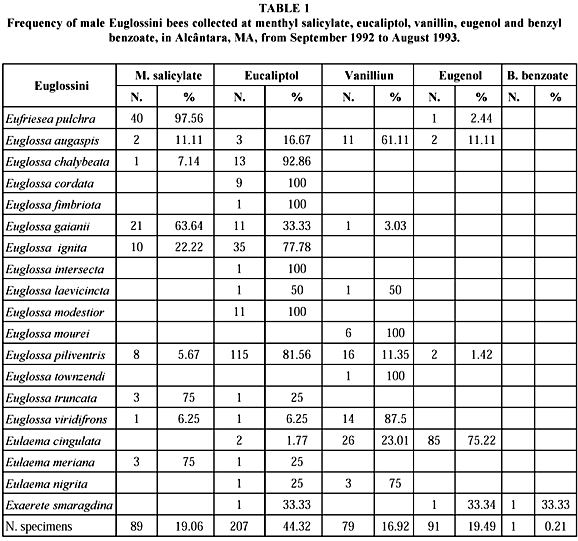
Four hundred sixty-seven male euglossini bees were captured, belonging to 4 genera and 19 species. Euglossa was the most abundant genera and showed the highest diversity (302 specimens and 14 species), followed by Eulaema (121; 3), Eufriesea (41; 1) and Exaerete (3; 1) (Table 1).
Comparing these results to those of Gomes, 1991 (1,728 specimens and 13 species), Fernandes, 1991 (428 and 9), Silva & Rebelo, 1999 (1,740 and 37), and Ferreira, 1994 (69 and 11), all done in Maranhão State, shows that the largest diversity and fauna abundance were obtained by Silva & Rebelo (1999) in a 10,000 ha area in the Forest Reserve of Vale do Rio Doce in Buriticupu, an Amazonian region of the state (Table 1). In spite of the present study having been done in a forest fragment along the Pepital River, a secondary vegetation area smaller than the study areas cited above, its results were second in specific diversity. Although Powell & Powell (1987) noted that there were fewer nisitors of the Euglossini species in accordance with fragment size, it also shows that in the absence of big forested areas, the small ones are utilized as refuges by fauna. This result reinforces the importance of developing conservation studies in fragments of natural ecosystems. This can also be argued because in 1988, 31.02% of Maranhão State's original forest areas were devastated, and 2.11% of the remaining area were located in fragments smaller than 10,000 ha, as cited by Skole & Tuckers (1993).
The species which more frequently visited the baits were Euglossa piliventris (141 specimens, 30.19% of the total), Eulaema cingulata (113, 24.2%), Euglossa ignita (45, 9.64%), Eufriesea pulchra (41, 8.78%) and Euglossa gaianii (33, 7.07%) corresponding to 79.88% of the sampling universe (Table 2).
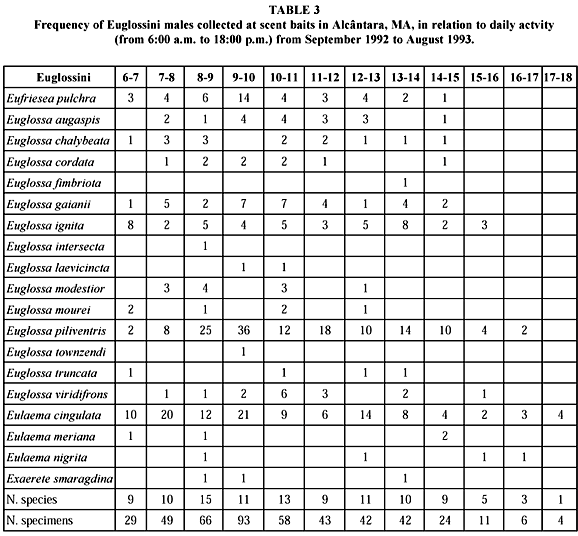
The Euglossini were active throughout the year, however during rainy period more activity and specific diversity were observed. More active specimens representing 17.13% of the total specimens sampled, occurred in March 1993, the rainiest period, and the least activity was observed in September-November 1992 (Fig. 1 and Table 3). Strong rains probably explain the reduced activity registered in April.
Euglossapiliventris was present during the whole year, especially in December (32 specimens, representing 65.31% of the total sampled during that month). The dominance of this species contradicts the results in the studies of Silva & Rebelo (1999), Powell & Powell (1987), and Oliveira & Campos (1995), in which this species was rare.
E. ignita and E. gaianii were active for almost the whole year, except in October and November 1992. Euglossa augaspis was strongly seasonal, being in activity during September and October/1992 and from May to August 1993. Another species with strong preference for some period of the year was E. viridifrons, active in December 1992 and January 1993, absent in February and active again in March and April.
Although E. mourei, E. modestior, E. intersecta, E. townsendi, E. truncata, E. fimbriota, and E. laevicincta were represented by few specimens, they also demonstrated evidences of seasonality. These species were active especially in April and August 1993, corresponding to the end of the rainy and beginning of the dry season.
The most abundant species of Euglossa (E. piliventris, E. ignita, and E. gaianii) weren't necessarily the same in other surveys in forest areas in Maranhão. Gomes (1991) and Fernandes (1991) reported E. chalybeata and E. ignita as the most abundant, a justificable similarity because they sampled the same area, the Sacavém Forest Reserve (760 ha), in São Luís. However, Silva & Rebelo (1999) obtained dominance of E. pleostica, E. truncata and E. avicula. Janzen et al. (1982) argue that shifts of Euglossini populations and communities in different or isolated geographic habitats are acculiarly frequent.
Among the three species sampled of the genera Eulaema, E. cingulata were active during the entire sampling period, with more activity in February and March 1993. E. nigrita and E. meriana were represented by few specimens, but all were sampled in the rainy period. The low frequency of E. nigrita, in this area reinforces data showing this species, preference for open and dry areas (Ducke, 1902; Zucchi et al., 1969; Rebelo & Garófalo, 1991). In Buriticupu, transition of forest and "cerrado", E. nigrita was one of the most frequent bees found (Silva & Rebelo, 1999). Before this study, the only wet forest where it was frequently sampled was São Luís, where the vegetation is more similar to that of the Amazon region as observed by Gomes (1991) and Fernandes (1991).
The low abundance of E. meriana, as compared to that shown in Gomes (1991), Fernandes (1991), Silva & Rebelo (1999), Morato et al. (1992), Powell & Powell (1987) and Oliveira & Campos (1995), can be explained, as Braga (1976) argues its preference for natural forest environments.
The genus Eufriesea normally occurs in few months of the year (Ackerman et al., 1982; Ackerman, 1983). In this study it was represented by just one species, E. pulchra, observed visiting the baits during almost the whole year, except in September and December 1992.
Exaeretesmaragdina, parasites of euglossinis nests (Kinsey, 1979), had three specimens sampled in October 1992 and May 1993.
Janzen (1981) reported that although many euglossini populations can remain active during the entire years; they must reduce their population size and can thus remain in some sites according to the resources available.
According to Dressler (1968) and Williams & Dodson (1972), the seasonal patterns can be explained by the movement pattern of the species.
However, Ackerman (1983), in studying the diversity and seasonality of euglossini males in Central Panama, verified that seasonal patterns of nidification and emerging of the bees could propably cause the seasonal fluctuation of abundance. Zimmerman & Madrinan (1988) verified that young males visit odorific substances, because they need to collect these substances before establishing territories while old males visit nectar sources. Rebelo & Garófalo (1991) observed that 77% of the males collecting odorific substances were young. The seasonal variation in preference for odorific substances can be related to the orchid floration patterns (Oliveira & Campos, 1996).
During the day, the bees were most active between 9:00 and 10:00 o'clock (Table 3). However, the biggest diversity of species was observed between 8:00 and 9:00 o'clock.
The daily activity pattern of the specimens of Eulaema is unlike the general pattern observed: shows three daily peaks of activity, between 7:00 and 8:00 a.m., 9:00 and 10:00 a.m., and 12:00 and 1:00 p.m.
The most attractive essence was eucaliptol (44.32% of all captured specimens), followed by eugenol (19.49%), menthyl salicitate (19.06%), vanillin (16.92%), and benzil benzoate (0.21%). The number of specimens attracted to eucaliptol can be explained by the presence of E. piliventris, with 55.56% of the captured bees (Table 3).
The eucaliptol also attracted most species, 16 of the 19 sampled. It failed to attract E. townsendi, E. mourei and Eufriesea pulchra.
Silva & Rebelo (1999) found benzil benzoate attracted Eufriesea pulchra, Exaerete smaragdina, and Euglossa cordata, but in Alcântara this essence attracted only one specimen of Exaerete smaragdina, even with the presence of the other species mentioned above.
Table 1, shows the preference of Euglossini species for the baits exposed. E. piliventris was observed visiting four of the five compounds utilized, but 81.56% of these specimens were captured collecting eucaliptol. In the studies of Powell & Powell (1987), Ackerman (1989), Armbruster & McCormick (1990), Morato et al. (1992) and Silva & Rebelo (1999), E. ignita, E. chalybeata, E. moestior and E. cordata preferred cineol. However, in the present survey these species were more abundant in eucaliptol.
For Euglossa gaiani, represented by 33 specimens, 21 (63.64%) were attracted to methyl salicitate, 11 (33.33%) to eucaliptol, and only 1 (3.03%) to vanillin; this preference to methyl salicitate was observed by Powell & Powell (1987), Armruster & McCormick (1990), and Morato et al. (1992).
Euglossaaugaspis and E. viridifrons also demonstrated a preference for a specific compound; 61.11% and 87.5% of their specimens were attracted to vanillin.
The other Euglossa species were represented by less than ten specimens, making a detailed analysis about their preferences for chemical compounds more difficult.
Eulaemacingulata was strongly attracted to eugenol, as indicated B' Janzen (1981), Janzen et al. (1982) and Morato et al. (1992). The other species of this genus were E. meriana, attracted to methyl salicitate and eucaliptol, and E. nigrita, attracted to eucaliptol and vanillin. Discussion about their preferences for the baits in the sample site is almost impossible because they were represented by a reduced number of individuals.
Eufrieseapulchra showed 97.56% of captured specimens visiting methyl salicitate; its preference for this essence is widely known as mentioned by Braga (1976), Ackerman (1989), Morato et al. (1992), and Silva & Rebelo (1999).
Acknowledgments We acknowledge the Fundação de Amparo à Pesquisa no Maranhão, Fapema (in memorium), for supporting this research and Pe. Jesus Santiago Moure (Federal University of Paraná), for identifying species.
- ACKERMAN, J. D., 1983, Diversity and seasonality of male euglossine bees (Hymenoptera: Apidae) in central Panama. Ecology, 64(2): 274-283.
- ACKERMAN, J. D., 1989, Geographic and seasonal variation in fragrance choices and preferences of male euglossine bees. Biotropica, 21(4): 340-347.
- ACKERMAN, J. D., MESLER, M. R., LU, K. L. & MONTALVO, A. M., 1982, Food-foraging behavior of male Euglossini (Hymenoptera: Apidae): Vagabonds or Trapliners? Biotropica, 14(4): 241-248.
- ARMBRUSTER, W. S. & McCORMICK, K. D., 1990, Diel foraging patterns of male Euglossine bees: Ecological Causes and Evolutionary Response. Biotropica, 22(2): 160-171.
- ARMBRUSTER,W. S. & WEBSTER, G. L., 1979, Pollination of two species of Dalechampia (Euphorbiaceae) in Mexico by euglossine bees. Biotropica, 11: 278-283.
- BECKER, P., MOURE, J. S. & PERALTA, F. J. A., 1991, More about euglossine bees in Amazonian Forest Fragments. Biotropica, 23: 340-591.
- BENNETT, F. D., 1972, Baited McPhail fruitfly traps to collect euglossine bees. J. New York Ent. Soc., 80: 137-145.
- BRAGA, P. I. S., 1976, Atraçăo de abelhas polinizadoras de orchidaceae com auxílio de iscas-odores na campina, campinarana e floresta tropical úmida da regiăo de Manaus. Cięncia e Cultura, 28: 767-773.
- BRASIL, 1991, Diagnóstico dos principais problemas ambientais do Estado do Maranhão Instituto Brasileiro de Meio Ambiente e dos Recursos Naturais renováveis (Ibama), Secretaria de Estado do Meio Ambiente e Turismo do Maranhão (Sematur). São Luís.
- DODSON, C. H., 1970, The role of chemical attractans in orchid pollination. In: K. L. Chambers (ed.), Biochemical Coevolution Corvallis. OR, Oregon State Univ., pp. 83-1077.
- DRESSER, R. L., 1968, Observations on orchids and euglossine bees in Panama and Costa Rica. Rev. Biol. Trop, 15(1): 143-183.
- DUCKE, A., 1902, As espécies paraenses do gęnero Euglossa Latr. Bio. Mus. Goeldi, 3: 561-575.
- FEITOSA, A. C., 1983, O Maranhăo Primitivo: uma tentativa de reconstituiçăo Editora Augusta, Săo Luís, 142p.
- FERNANDES, A. A., 1991, Avaliaçăo do grau de preferęncia de abelhas Euglossini a ambientes com diferentes níveis de degradaçăo. Monografia apresentada ao curso de Cięncias Biológicas da Universidade Federal do Maranhăo, Săo Luís, 51p.
- FERREIRA, R. das G., 1994, Estrutura de comunidades de abelhas (Hymenoptera: Apoidea) numa regiăo de baixada em Vitória do Mearim, MA, Brasil Monografia apresentada ao curso de Cięncias Biológicas, UFMA.
- GOMES, L. F., 1991, Diversidade e flutuaçăo de abelhas Euglossini em diferentes ecossistemas do Maranhăo. Monografia apresentada ao curso de Cięncias Biológicas da Universidade Federal do Maranhăo, Săo Luís, 60p.
- JANZEN, D. H., 1971, Euglossine bees as long-distance pollinators of tropical plants. Science, 171: 203-205.
- JANZEN, D. H., 1981, Reduction in euglossine bee species richness on Isla del Cano, a Costa Rican Offshore Island. Biotropica, 13(3): 238-240.
- JANZEN, D. H., DEVRIES, P. J., HIGGINS, M. L. & KINSEY, L. S., 1982, Seasonal and site variation in Costa Rican euglossine bees at chemical baits in lowland deciduos and evergreen forests. Ecology, 63: 66-74.
- KINSEY, S. L., 1979, An illustrated key to the genus Exarete with descriptions of male genitalia and biology. J. Kans. Ent. Soc, 52: 735-746.
- MORATO, M. L., CAMPOS, L. A. O. & MOURE, J. S., 1992, Abelhas Euglossini (Hymenoptera: Apidae) coletadas na Amazônia Central. Rev. Bras. Ent, 36(4): 767-771.
- OLIVEIRA, M. L., 1999, Sazonalidade e horário de atividade de abelhas Euglossinae (Hymenoptera, Apidae), em floresta de terra firme na Amazônia Central. Rev. Bras. Zool., 16(1): 83-90.
- OLIVEIRA, M. L. & CAMPOS, L. A. O., 1995, Abundância, riqueza e diversidade de abelhas Euglossinae (Hymenoptera: Apidae) em florestas contínuas de terra firme na Amazônia Central, Brasil. Rev. Bras. Zool, 12(3): 547-556.
- OLIVEIRA, M. L. & CAMPOS, L. A. O., 1996, Preferęncia por estratos florestais e por substâncias odoríferas em abelhas Euglossinae (Hymenoptera, Apidae). Rev. Bras. Zool, 13(4): 1075.
- PEARSON, D. L. & DRESSLER, R. L., 1985, Two-year study of male orchid bee (Hymenoptera, Apidae, Euglossini) attraction to chemical baits in lowland South-easter Peru. Jour. Trop. Ecol, 1: 37-54.
- POWELL, A. H. & POWELL, G. V. N., 1987, Population dynamics of male euglossine bees in Amazonian forest fragments. Biotropica, 19: 176-179.
- RAW, A., 1989, The dispersal of euglossine bees between isolated patches of eastern Brazilian wet forest (Hymenoptera: Apidae). Rev. Bras. Ent, 33(1): 103-107.
- REBELO, J. M. M. & CABRAL, A. J., 1997, Abelhas Euglossinae de Barreirinhas, zona do litoreal da baixada oriental maranhense. Acta Amazonica, 27: 145-152.
- REBELO, J. M. M. & GARÓFALO, C. A., 1991, Diversidade e sazonalidade de machos de Euglossini (Hymenoptera: Apidae) e preferęncias por iscas-odores em um fragmento de floresta no sudeste do Brasil. Rev. Brasil. Biol., 51: 787-799.
- REBELO, J. M. M. & GARÓFALO, C. A., 1997, Comunidades de machos de euglossini (Hymenoptera: Apidae) em matas semidecíduas do nordeste do Estado de Săo Paulo. An. Soc. Entomol. Brasil, 26: 243-255.
- ROUBIK, D. W. & ACKERMAN, J. D., 1987, Long-term ecology of euglossine orchid-bees (Apidae: Euglossini) in Panama. Oecologia, 73: 321-333.
- SILVA, F. S. & REBELO, J. M. M., 1999, Euglossine bees (Hymenoptera: Apidae) of Buriticupu, Amazonia of Maranhăo, Brazil. Acta Amazonica, 29(4): 587-599.
- SKOLE, D. & TUCKER, C., 1993, Tropical deforestation in the Amazon: Satellite data from 1978 to 1988. Science, 260: 1.905-1.910.
- WILLIAMS, N. H., 1982, The biology of orchids and euglossine bees. In: J. Arditti (ed.), Orchids biology: Reviews and Perspectives, 11: 119-171. Cornell University Press, Ithaca, New York.
- WILLIAMS, N. H. & DODSON, C. H., 1972, Selective attraction of male euglossine bees to orchid floral fragrance and its importance in long distance pollen flow. Evolution, 26: 84-95.
- WILLIAMS, N. H. & WITTHEN, W. M., 1983, Orchid floral fragrances and male euglossine bees. Methods and advances in the last sesquidecade. Biol. Bull, 164: 355-395.
- ZIMMERMAN, J. K. & MADRINAN, S. R., 1988, Age structure of male Euglossa imperialis (Hymenoptera, Apidae, Euglossini) at nectar and chemical sources in Panama. Jour Trop. Ecol., 4: 303-306.
- ZUCCHI, R., SAKAGAMI, S. F. & CAMARGO, J. M. F., 1969, Biological observations on a neotropical parasocial bee, Eulaema nigrita, with a review on the biology of Euglossinae (Hymenoptera, Apidae). A comparative study. J. Fac. Sci. Hokkaido, Univ. Ser. Vi. Zool, 17: 271-382.
Publication Dates
-
Publication in this collection
24 May 2002 -
Date of issue
Nov 2001
History
-
Accepted
18 Aug 2000 -
Received
22 May 2000