Abstracts
We assessed spatial and seasonal changes in the diet of Oligosarcus hepsetus in order to describe the strategy developed by this species that allows their very high abundance in Lajes reservoir, Rio de Janeiro, Brazil. Fish samplings were carried out using gill nets, deployed during ca. 12 and 24 hours, between April 2001 and May 2002. A total of 289 individuals were examined, of which 97 showed gut contents. We used the index of relative importance (IRI) to compare probable dietary shifts, and the frequency of occurrence (% OC) to analyze possible ontogenetic influences on feeding. O. hepsetus showed carnivorous habits, feeding preferably on fish and insects, the latter of which occurred in 71.0% of the guts presenting contents. O. hepsetus consumed different items along the three reservoir zones: insects (61.0% IRI) and Cichla monoculus (38.9% IRI) in the lower zone; Lepidoptera (57.0% IRI) in the middle zone; and C. monoculus (77.0% IRI) in the upper zone. Food items changed seasonally with C. monoculus predominating in autumn 2001, and Hymenoptera and Lepidoptera in the winter. In spring almost all food was Lepidoptera (99.8% IRI), while in the summer Hemiptera dominated in the diet. In autumn 2002 Hemiptera (97.0% IRI) was dominant, in significant contrast with the previous autumn. Individuals smaller than 190 mm SL fed heavily on insects, while fishes predominated in the diet of individuals larger than 190 mm SL. Shifts in prey-capture ability among length classes suggest decreasing intraspecific competition. A higher food plasticity seems to be the strategy employed by this opportunist species, which used food resources available in the reservoir.
food habit; diet; Characiforms; reservoirs
Foram analisadas variações na dieta de O. hepsetus conforme comparações espaciais e sazonais, com o objetivo de descrever a estratégia desenvolvida por essa espécie que permite sua elevada abundância no reservatório de Lajes, Rio de Janeiro, Brasil. As amostras foram coletadas utilizando redes de espera com permanência aproximadamente de 12 e 24 horas, no período entre abril de 2001 e maio de 2002. Dos 289 exemplares capturados, 97 peixes continham alimento no estômago. A análise foi baseada no índice de importância relativa (IIR), enquanto eventuais influências ontogenéticas foram determinadas pela freqüência de ocorrência (% FO). O. hepsetus apresentou hábito alimentar carnívoro, alimentando-se preferencialmente de peixes e insetos. O item Insetos apresentou relevante participação na dieta, ocorrendo em 71% dos estômagos que continham alimento. O. hepsetus consumiu diferentes itens entre as três zonas do reservatório: insetos (61% IIR) e Cichla monoculus (38,9% IIR) na zona baixa; Lepidoptera (57% IIR) na zona intermediária; e C. monoculus (77% IIR) na zona alta. No geral, ocorreram diferenciações durante todo o ciclo sazonal, em que C. monoculus predominou no outono de 2001 e Hymenoptera e Lepidoptera, no inverno. Na primavera, a alimentação foi basicamente constituída por Lepidoptera (99,8% IIR), enquanto no verão, Hemiptera predominou na dieta. No outono de 2002 foi registrado 97% IIR para Hemiptera, diferindo do outono anterior. Indivíduos menores que 190 mm de comprimento-padrão (CP) consumiram preferencialmente insetos, enquanto os espécimes de maior tamanho (> 190 mm CP) alimentaram-se principalmente de peixes. Mudanças na captura de presas entre classes de tamanho sugerem diminuição da competição intra-específica. A alta plasticidade na alimentação parece ser uma estratégia usada por essa espécie oportunista, consumindo os recursos alimentares disponíveis no reservatório.
hábito alimentar; dieta; Characiformes; reservatórios
Spatial and seasonal changes in the diet of Oligosarcus hepsetus (Characiformes, Characidae) in a Brazilian Reservoir
Variações espaciais e sazonais na dieta de Oligosarcus hepsetus (Characiformes, Characidae) em um reservatório brasileiro
Araújo, F. G; Andrade, C. C; Santos, R. N; Santos, A. F. G. N; Santos, L. N
Laboratório de Ecologia de Peixes, Universidade Federal Rural do Rio de Janeiro, km 47, Antiga Rodovia Rio-SP, CEP 23851-970, Seropédica, RJ, Brazil
Correspondence to Correspondence to Francisco Gerson Araújo Laboratório de Ecologia de Peixe Universidade Federal Rural do Rio de Janeiro km 47, Antiga Rodovia Rio-SP CEP 23851-970, Seropédica, RJ, Brazil E-mail: gerson@ufrrj.br
ABSTRACT
We assessed spatial and seasonal changes in the diet of Oligosarcus hepsetus in order to describe the strategy developed by this species that allows their very high abundance in Lajes reservoir, Rio de Janeiro, Brazil. Fish samplings were carried out using gill nets, deployed during ca. 12 and 24 hours, between April 2001 and May 2002. A total of 289 individuals were examined, of which 97 showed gut contents. We used the index of relative importance (IRI) to compare probable dietary shifts, and the frequency of occurrence (% OC) to analyze possible ontogenetic influences on feeding. O. hepsetus showed carnivorous habits, feeding preferably on fish and insects, the latter of which occurred in 71.0% of the guts presenting contents. O. hepsetus consumed different items along the three reservoir zones: insects (61.0% IRI) and Cichla monoculus (38.9% IRI) in the lower zone; Lepidoptera (57.0% IRI) in the middle zone; and C. monoculus (77.0% IRI) in the upper zone. Food items changed seasonally with C. monoculus predominating in autumn 2001, and Hymenoptera and Lepidoptera in the winter. In spring almost all food was Lepidoptera (99.8% IRI), while in the summer Hemiptera dominated in the diet. In autumn 2002 Hemiptera (97.0% IRI) was dominant, in significant contrast with the previous autumn. Individuals smaller than 190 mm SL fed heavily on insects, while fishes predominated in the diet of individuals larger than 190 mm SL. Shifts in prey-capture ability among length classes suggest decreasing intraspecific competition. A higher food plasticity seems to be the strategy employed by this opportunist species, which used food resources available in the reservoir.
Key words: food habit, diet, Characiforms, reservoirs.
RESUMO
Foram analisadas variações na dieta de O. hepsetus conforme comparações espaciais e sazonais, com o objetivo de descrever a estratégia desenvolvida por essa espécie que permite sua elevada abundância no reservatório de Lajes, Rio de Janeiro, Brasil. As amostras foram coletadas utilizando redes de espera com permanência aproximadamente de 12 e 24 horas, no período entre abril de 2001 e maio de 2002. Dos 289 exemplares capturados, 97 peixes continham alimento no estômago. A análise foi baseada no índice de importância relativa (IIR), enquanto eventuais influências ontogenéticas foram determinadas pela freqüência de ocorrência (% FO). O. hepsetus apresentou hábito alimentar carnívoro, alimentando-se preferencialmente de peixes e insetos. O item Insetos apresentou relevante participação na dieta, ocorrendo em 71% dos estômagos que continham alimento. O. hepsetus consumiu diferentes itens entre as três zonas do reservatório: insetos (61% IIR) e Cichla monoculus (38,9% IIR) na zona baixa; Lepidoptera (57% IIR) na zona intermediária; e C. monoculus (77% IIR) na zona alta. No geral, ocorreram diferenciações durante todo o ciclo sazonal, em que C. monoculus predominou no outono de 2001 e Hymenoptera e Lepidoptera, no inverno. Na primavera, a alimentação foi basicamente constituída por Lepidoptera (99,8% IIR), enquanto no verão, Hemiptera predominou na dieta. No outono de 2002 foi registrado 97% IIR para Hemiptera, diferindo do outono anterior. Indivíduos menores que 190 mm de comprimento-padrão (CP) consumiram preferencialmente insetos, enquanto os espécimes de maior tamanho (> 190 mm CP) alimentaram-se principalmente de peixes. Mudanças na captura de presas entre classes de tamanho sugerem diminuição da competição intra-específica. A alta plasticidade na alimentação parece ser uma estratégia usada por essa espécie oportunista, consumindo os recursos alimentares disponíveis no reservatório.
Palavras-chave: hábito alimentar, dieta, Characiformes, reservatórios.
INTRODUCTION
Oligosarcus hepsetus (Cuvier 1829) is a very common Characidae species that inhabits almost all freshwater environments in southeastern Brazil and prefers densely-vegetated shallow microhabitats in small tributaries or littoral zones of the main rivers. Most Oligosarcus species showed a carnivorous feeding habit, feeding on insects, crustaceans and small fish (Lowe Mc-Connel, 1975). In Lajes reservoir, the largest impoundment in Rio de Janeiro State, O. hepsetus is one of the most successful species, playing an important ecological role within the trophic chain since the juveniles are eaten by large piscivorous fishes (SANTOS et al., 2001).
Trophic ecology studies of carnivorous species are very useful to fishery monitoring and management programs, since they provide important data about regulation mechanisms in fish populations and communities (Zavala-Camim, 1996). Remarkable seasonal changes can take place in fish diets and according to area. These are primarily related to shifts in food resource composition and availability, which are associated to reproductive pulses and physicochemical changes in aquatic ecosystems. Diet also can be influenced by ontogenetic effects, which for their part are usually related to morphological changes that lead to shifts in prey selection and capture (Nikolsky, 1963; Wooton, 1992). Therefore, fish-diet knowledge provides information essential to autoecological research as it supplies additional data on the trophic structure of an entire ecosystem (Basile-Martins et al., 1983).
Feeding studies on the fish community of Lajes reservoir have focused only on Cichla monoculus, an introduced piscivorous species, while little attention have been given to other native carnivorous species (Santos et al., 2001). No studies on trophic ecology of O. hepsetus have been reported either for Lajes reservoir or for other artificial lakes of southeastern Brazil. Likewise, few researchers have focused on spatial and temporal dietary switches, as influenced by biotic and abiotic variations, in the Oligosarcus species (Hartz et al., 1996; Gealh & Hahn, 1998). Since a previous study of the Lajes reservoir had shown the fish community patterns as changing spatially and seasonally (Araújo & Santos, 2001), we investigated the diet of O. hepsetus, relating the spatial-seasonal changes of its food intake with the peculiar environmental characteristics of this reservoir. Possible ontogenetic effects as well as the relationships between the O. hepsetus diet and the ecology of other reservoir fish species also were considered, in order to evaluate how its feeding strategy could have led to the high success of this species in the impoundment.
MATERIAL AND METHODS
Study area
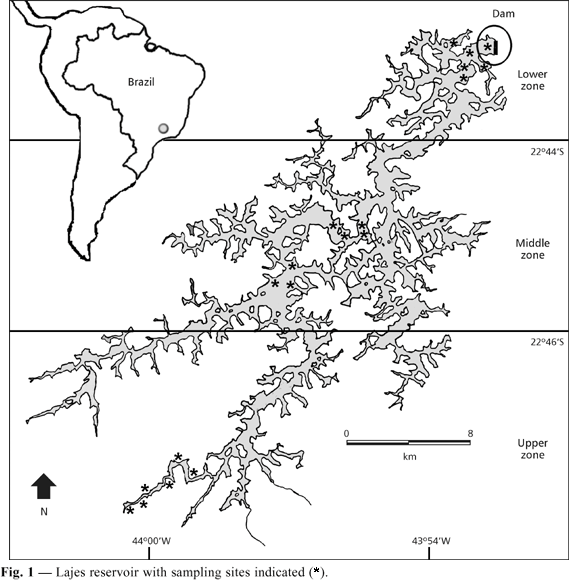
Located on the upper slopes of the hills of the southeastern Brazilian coast, Lajes reservoir (22º42'-22º50'S; 43º53'-44º05'W) is a major impoundment in Rio de Janeiro State. The reservoir, located at ca. 415 m above mean sea level, occupies about 30 km2 and is a well-preserved oligotrophic lake, surrounded by stretches of Atlantic Forest in which no large urban-industrial centers are located (Araújo & Santos, 2001).
According to Santos et al. (2001), the reservoir is divided into three zones (Fig. 1) having different environmental characteristics: (1) the upper zone, situated near the reservoir headwaters at the entrance of the main tributaries. The slope is mainly plain with few steep areas. Depth is shallow compared to other zones, and the marginal area is occasionally colonized by aquatic macrophytes; (2) middle zone, located between the upper and the lower zones, which is the largest area and comprises a great number of bays, several islands, and exitless waterways; and (3) lower zone, situated near the dam, where sharp slopes and greater depths predominate, and in which macrophytes and other submersed shelters are lacking.
Fish sampling and data analysis
Two sampling programs, using gillnets 20 m long x 1.5 in height, with mesh size ranging from 2.5 to 5.5 mm, were designed to collect fishes. From April 2001 to January 2002, nine gillnets were set at three-month intervals in each reservoir zone. From May 2001 to May 2002, twelve to fifteen gillnets were installed only in the lower zone, except in months included in the preceding program. Nets were kept in the water for 12 and 24 hours for the first and last sampling programs, respectively.
All fishes were preserved in ice in the field, and subsequently transferred to the laboratory, where they were counted, weighed (to nearest 0.1 grams), measured (to 1.0 mm), and dissected for digestive tract analysis. All food items were examined using a stereoscopic microscope and identified to the lower taxonomic level possible.
They were counted, weighed (0.001 grams), and measured (0.01 mm). Gut contents in individuals were analyzed by the index of relative importance (IRI) (Pinkas, 1971), which deals with numerical (% NM) and gravimetrical (% GR) percents and frequency of occurrence (% OC), where IRI = % FO x (% FN + % FP) x 100. The percent IRI (% IRI) was calculated dividing the IRI values from each food item by the sum of all IRI values.
The IRI was compared among zones (lower, middle, and upper), and seasons as follows: autumn 2001: April to June; winter: July to September; spring: October to December; summer: January to March; and autumn 2002: April to May. The frequency of occurrence (% OC) was also calculated for each food item along the length classes (mm) so as to detect possible ontogenetic influences on the diet.
RESULTS
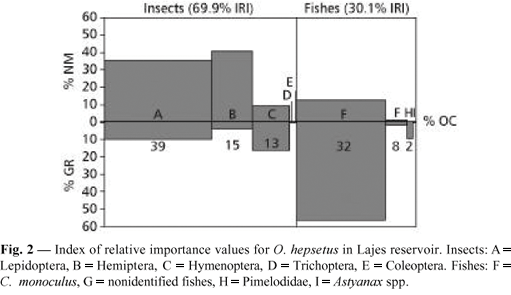
A total of 289 individuals were examined, of which 97 (33.56%) showed some gut content, ranging in standard length (SL) from 130 to 224 mm. The diet was comprised by insects (56.04% IRI) and fishes (43.9% IRI): Cichla monoculus (43.0% IRI); Lepidoptera (36.0% IRI); Hemiptera (14.0% IRI); and Hymenoptera (6.0% IRI). Other minor components of the diet, e.g., Pimelodidae, Astyanax spp., nonidentified fishes, Trichoptera, and Coleoptera contributed less than 1.0% IRI (Fig. 2).
The diet changed along the reservoir zones (Fig. 3): insects (61.0% IRI), namely Lepidoptera (40.0% IRI), Hemiptera (12.0% IRI), and Hymenoptera (9.0% IRI), and C. monoculus (39.0% IRI) were the main items consumed in the lower zone; Pimelodidae, Astyanax spp., nonidentified fishes, Trichoptera, and Coleoptera contributed less than 1.0% IRI. In the middle zone, Lepidoptera (57.0% IRI), C. monoculus (30.0% IRI), and non-identified fishes (13.0% IRI) occurred in the gut content, while C. monoculus (77.0% IRI), Pimelodidae (6.0% IRI), and Hemiptera (12.0% IRI); nonidentified fishes, Lepidoptera, and Hymenoptera amounted to 4.0% IRI.
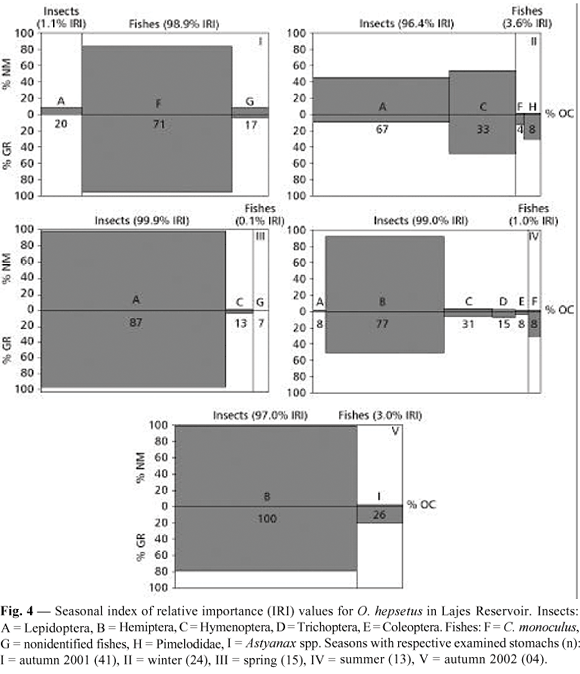
Seasonal dietary shifts were found (Fig. 4). C. monoculus was the major prey in autumn 2001 (97.0% IRI), with nonidentified fishes and Lepidoptera showing little contribution (2.0 and 1.0% IRI, respectively). Lepidoptera (49.0% IRI) and Hymenoptera (46.0% IRI) dominated during winter, with little contribution of Pimelodidae (4.0% IRI) and C. monoculus (1.0% IRI). Lepidoptera was the almost exclusive prey in spring (99.0% IRI), while nonidentified fishes and Hymenoptera provided the remaining items. Hemiptera (94.0% IRI) was the main item in summer, followed by C. monoculus, Hymenoptera, Trichoptera, Coleoptera, and Lepidoptera. In autumn 2002, Hemiptera (97.0% IRI) dominated in the diet, followed by Astyanax spp. (3.0% IRI).
Oligosarsus hepsetus consumed both insects and fishes along all the length classes (Table 1). Individuals smaller than 190 mm SL fed heavily on insects, mainly Lepidoptera and Hemiptera, with fishes (restricted to C. monoculus and Astyanax spp.) having minor dietary importance. Fishes predominated in the diet of individuals larger than 190 mm SL, with C. monoculus and Pimelodidae being the major items consumed. Lepidoptera and Hemiptera showed lower contributions. O. hepsetus smaller than 150 mm SL did not eat Hymenoptera, although this food item frequently occurred in stomachs of larger individuals.
DISCUSSION
Oligosarcus hepsetus showed a carnivorous food habit in Lajes reservoir, feeding solely on insects and fishes and changing diet over the seasons and the reservoir zones. These major items were also found in the diet of other congeners in distinct ecosystems. Hahn et al. (1997) and Gealh & Hahn (1998) recorded a carnivorous diet in Oligosarcus longirostris in the Segredo reservoir, Paraná, where juveniles ate fishes, crustaceans, and insects while adults fed exclusively on fishes. Vieira (1994) reported a similar food habit for Oligosarcus solitarius from several lakes of the Doce river, Minas Gerais, with juveniles feeding on insects and prawns, while adults consuming only fishes. HARTZ et al. (1996) found high prey diversity in Oligosarcus jenynsii guts, with fishes, decapod crustaceans, and insects (mainly diptera) being the major items. The exclusive insectivorous-piscivorous habit suggests a specialized diet for O. hepsetus in Lajes reservoir. However, the shifts in major items consumed along the zones and seasons suggests a rather opportunistic strategy for this species, which switches from an insectivorous to piscivorous diet and vice versa according to resource availability. Nikolsky (1963) and Wooton (1992) have affirmed that spatial and seasonal shifts in fishes diet are very common, and usually associated with prey reproductive pulses and environmental changes in an ecosystem; food availability is another important factor dictating these changes in the diet.
The predominantly insectivorous habit in the lower and middle zones, which are comprised by allochthonous resources, probably reflects the relatively low productivity of these zones, which is related to high transparencies and depths, steep margins, and lack of macrophytes. On the other hand, the piscivorous habit developed in upper zone seems to be associated to the higher capacity of this zone to support autochthonous organisms, mainly fishes. Among all food items, Lepidoptera and Cichla monoculus were, respectively, the allochthonous and autochthonous resources most used by O. hepsetus.
O. hepsetus changes its diet markedly according to the season. C. monoculus was the dominant prey in utumn 2001, while insects dominated in the other seasons. Lepidoptera and Hymnenoptera were the main food items in winter, Lepidoptera in spring, and Hemiptera in summer and autumn 2002. High numbers of juvenile C. monoculus (SL < 100 mm) during autumn are common in the reservoir due to summer spawning (Fontenele, 1982), which is possibly linked to the high proportion of C. monoculus found in the diet in autumn 2001. The reasons for no occurrence of C. monoculus in the diet in autumn 2002 are unknown, but delays or weak reproductive peaks in that period could explain this dietary shifts.
Size could be a further constraint on food selection. The individuals caught for this study were > 130 mm SL. Hartz et al. (1996) also reported that Oligosarcus jenynsii (Günther, 1864) ranging from 106 to 264 mm TL fed on insects, fishes, and other smaller items.
There was a trend for the larger-sized O. hepsetus (> 190 mm SL) to feed mainly on fishes when compared to smaller individuals that ate mostly insects. Gealh & Hahn (1998) found similar results for O. longirostris in Segredo reservoir, with smaller individuals eating insects, crustaceans, and fishes, while larger ones (> 160 mm SL) were exclusively piscivorous. Larger-sized O. hepsetus (> 190 mm SL) were apparently able to eat more rewarding prey: larger, heavier, and probably more energetic, since length increase is related to morphological changes (e.g., gape size and swimming speed) that could favor fish capture. Ontogenetic shifts in the diet also can be associated to a decrease in intraspecific competition, with smaller individuals preferentially eating insects and larger ones consuming mostly fishes.
No cannibal behavior was shown for O. hepsetus in Lajes reservoir, while the other carnivorous species, C. monoculus, showed intense cannibalism in certain period of its life cycle (SANTOS et al., 2001). This could mean that this native carnivore is capable of using a greater variety of food resources than is the nonnative piscivorous C. monoculus, thus minimizing negative intra-specific relationships.
These two species also exhibited complex predator-prey interactions. According to SANTOS et al. (2001), O. hepsetus showed a low contribution to the C. monoculus diet (< 3.0% IRI), while in the present study C. monoculus comprised the major prey eaten by O. hepsetus (43.0% IRI). Therefore, O. hepsetus seems to play a key role in the reservoir food web, acting as prey for large piscivorous fishes and predator for smaller ones.
To sum up, the insectivorous-piscivorous habit showed by O. hepsetus, using both autochthonous and allochthonous resources that changed at both spatial and seasonal scales might explain the success of this species in Lajes reservoir. Additionally, O. hepsetus could be an important biologic constraint for C. monoculus.
Acknowledgements This study was partially financed by LIGHT Electricity Services S.A., concessionaire of Lajes reservoir, which supplied the infrastructure essential to the development of this project. We thank the technicians and undergraduate students at the Fish Ecology Laboratory, the University Federal Rural of Rio de Janeiro, for helping in field and laboratory work. We are also grateful to an anonymous referee who contributed valuable comments.
Received May 6, 2003 - Accepted September 29, 2003 - Distributed February 28, 2005
- ARAÚJO, F. G. & SANTOS, L. N., 2001, Distribution of fish assemblages in Lajes reservoir, Rio de Janeiro, Brazil. Rev. Brasil. Biol., 61(4): 563-576.
- BASILE-MARTINS, M. A., CIPÓLI, M. N. & GODINHO, H. M., 1983, Alimentação do mandi Pimelodus maculatus Lacépède, 1803 (Osteichthyes, Pimelodidae) de trechos dos rios Jaguari e Piracicaba, São Paulo, Brasil. Bol. Inst. Pesca, São Paulo, 13(1): 17-19.
- FONTENELE, O., 1982, Contribuição para o estudo da biologia dos tucunarés (Actinopterygii, Cichlidae) em cativeiro - aparelho de reprodução, hábitos de desova e incubação. Colet. de Trab. Técn. DNOCS, Fortaleza, 2: 256-276.
- GEALH, A. M. & HAHN, N. S., 1998, Alimentação de Oligosarcus longirostris do reservatório de Salto Segredo, Paraná, Brasil. Revta. Bras. Zool., 15(4): 985-993.
- HAHN, N. S., FUGI, R., ALMEIDA, V. L. L., RUSSO, M. R. & LOUREIRO, V. E., 1997, Dieta e atividade alimentar de peixes do reservatório de Segredo. pp. 141-153. In: A. A. Agostinho & L. C. Gomes (eds.), Reservatório de Segredo: bases ecológicas para o manejo EDUEM, Maringá, 381p.
- HARTZ, S. M., MARTINS, A., & BARBIERI, G., 1996, Dinâmica da alimentação e dieta de Oligorsacus jenynsii (Günther, 1864) na lagoa Caconde, Rio Grande do Sul, Brasil (Teleoestei, Characidae). Bol. Inst. Pesca, São Paulo, 23: 21-29.
- LIGHT/IESA, 1991, Reservatório de Lajes - estudo de viabilidade para o alteamento do nível da água Relatório final SLP (RMD - 6102 - RE), Rio de Janeiro, 102p.
- LOWE-McCONNEL, R. L., 1975, Fish communities in tropical freshwaters Longman, London, 337p.
- NIKOLSKY, G. V., 1963, The ecology of fishes Academic Press, London, 352p.
- OLIVEIRA, S. L. DE, MENDES, Z. C., CRISÓSTOMO, L. C. & ARAÚJO, F. G., 1986, Resultados preliminares do levantamento ictiológico na represa de Ribeirão das Lajes, estado do Rio de Janeiro. Public. Avulsas Mus. Nac. do Rio de Janeiro, Rio de Janeiro, 65: 87-90.
- PINKAS, L., 1971, Food habits study. pp. 5-10. In: L. Pinkas, M. S. Oliphant, I. L. K. Iverson (eds.), Food habits of albacore bluefin tuna and bonito in California waters. Fish. Bull., 152: 1-105.
- SANTOS, L. N., GONZALEZ, A. F. & ARAÚJO, F. G., 2001, Dieta do tucunaré amarelo Cichla monoculus (Bloch & Schneider) (Osteichthyes, Ciclidae), no reservatório de Lajes, RJ. Revta. Bras. Zool, Curitiba, 18(Supl. 1): 191-204.
- VIEIRA, F., 1994, Estrutura de comunidade e aspectos da alimentação e reprodução dos peixes em dois lagos do médio Rio Doce, MG Editora Universidade Federal de Minas Gerais, Belo Horizonte, 78p.
- WOOTTON, R. J., 1992, Fish ecology Chapman and Hall, New York, 212p.
- ZAVALA-CAMIN, L. A., 1996, Introdução ao estudo sobre alimentação natural em peixes EDUEM, Maringá, 129p. Braz. J. Biol., 65(1): 1-8, 2005
Publication Dates
-
Publication in this collection
16 Nov 2005 -
Date of issue
Feb 2005
History
-
Accepted
29 Sept 2003 -
Received
06 May 2003