Abstracts
Morphological changes in the sexual organs of the pulmonates were observed throughout a year and correlated with reproductive-cycle periods. Reproductive-organ weights of the snail Megalobulimus abbreviatus were recorded seasonally and gonad sections were analyzed morphologically. The weights were used to obtain the organosomatic index. Mean oocytic diameter and oocytic maturation index were based on gonad sections. It was concluded that M. abbreviatus is an iteroparous snail whose annual reproductive cycle is characterized by mating and egg laying throughout spring and early summer, and also by reproductive system preparation, occurring over the remainder of the summer until the end of winter, for a new breeding season.
Pulmonata; Megalobulimus; reproductive cycle; reproductive system; ovotestis
Variações morfológicas nos órgãos do sistema reprodutor dos gastrópodes pulmonados são observadas ao longo do ano e podem ser correlacionadas a fases de seu ciclo reprodutivo. A partir dessa observação, a massa dos órgãos do sistema reprodutor do caracol Megalobulimus abbreviatus foi tomada em cada estação do ano e foram obtidas secções histológicas das gônadas. Os valores de massa foram utilizados para a obtenção do índice organo-somático e as secções, para calcular o diâmetro médio dos ovócitos e o índice de maturação ovocitária. Concluímos que M. abbreviatus é um caracol "iteroparous", apresentando ciclo reprodutivo anual caracterizado por acasalamento e oviposição durante a primavera e o início do verão e pela preparação do sistema reprodutor para um nova fase reprodutiva, que se inicia em janeiro e se estende até o final do inverno.
Pulmonata; Megalobulimus; ciclo reprodutivo; sistema reprodutor; gônada
The annual reproductive cycle of the snail Megalobulimus abbreviatus (Bequaert, 1948) (Gastropoda, Pulmonata)
Ciclo reprodutivo anual do caracol Megalobulimus abbreviatus (Bequaert, 1948) (Gastropoda, Pulmonata)
Horn, A. C. M.I; Achaval, A.I; Zancan, D. M.II
ILaboratório de Histofisiologia Comparada, Departamento de Ciências Morfológicas
IILaboratório de Neurobiologia Comparada, Departamento de Fisiologia, Instituto de Ciências Básicas da Saúde, Universidade Federal do Rio Grande do Sul, Rua Sarmento Leite, 500, Cep 90050-170, Porto Alegre, RS, Brazil
Correspondence Correspondence to Ângelo Cássio Magalhães Horn Laboratório de Histofisiologia Comparada Departamento de Ciências Morfológicas Instituto de Ciências Básicas da Saúde, Universidade Federal do Rio Grande do Sul Rua Sarmento Leite, 500 CEP 90050-170, Porto Alegre, Brazil e-mail: angelocmh@terra.com.br
ABSTRACT
Morphological changes in the sexual organs of the pulmonates were observed throughout a year and correlated with reproductive-cycle periods. Reproductive-organ weights of the snail Megalobulimus abbreviatus were recorded seasonally and gonad sections were analyzed morphologically. The weights were used to obtain the organosomatic index. Mean oocytic diameter and oocytic maturation index were based on gonad sections. It was concluded that M. abbreviatus is an iteroparous snail whose annual reproductive cycle is characterized by mating and egg laying throughout spring and early summer, and also by reproductive system preparation, occurring over the remainder of the summer until the end of winter, for a new breeding season.
Key words: Pulmonata, Megalobulimus, reproductive cycle, reproductive system, ovotestis.
RESUMO
Variações morfológicas nos órgãos do sistema reprodutor dos gastrópodes pulmonados são observadas ao longo do ano e podem ser correlacionadas a fases de seu ciclo reprodutivo. A partir dessa observação, a massa dos órgãos do sistema reprodutor do caracol Megalobulimus abbreviatus foi tomada em cada estação do ano e foram obtidas secções histológicas das gônadas. Os valores de massa foram utilizados para a obtenção do índice organo-somático e as secções, para calcular o diâmetro médio dos ovócitos e o índice de maturação ovocitária. Concluímos que M. abbreviatus é um caracol "iteroparous", apresentando ciclo reprodutivo anual caracterizado por acasalamento e oviposição durante a primavera e o início do verão e pela preparação do sistema reprodutor para um nova fase reprodutiva, que se inicia em janeiro e se estende até o final do inverno.
Palavras-chave: Pulmonata, Megalobulimus, ciclo reprodutivo, sistema reprodutor, gônada.
INTRODUCTION
Megalobulimus abbreviatus is a giant South American snail that can be found in Argentina, Paraguay, and sub-tropical regions of Brazil (Bequaert, 1948; Sawaya & Petersen, 1962). Due to its size (Pitoni et al., 1976), M. abbreviatus and other congenerics have proved to be good experimental models for physiological (De Jorge et al., 1965; Jaeger, 1965; Marques & Pereira, 1970; Romero & Hoffmann, 1991; Belló-Klein et al., 1993; Rossi & Da Silva, 1993; Da Silva & Zancan, 1994; Romero et al., 1994) and morphological (Zancan et al., 1994; Zancan & Achaval, 1995; Zancan et al., 1997; Donelli et al., 1998; Faccioni-Heuser et al., 1999; Santos et al., 2002) studies.
Like all pulmonates, M. abbreviatus is a hermaphroditic snail having three identifiable regions in their reproductive apparatus: hermaphrodite, male, and female (Duncan, 1975; Tompa, 1984; Lutchel et al., 1997). Reproductive-cycle characteristics of stylommatophora based on a morphological analysis of the reproductive system have been established for several species, most of which are of temperate climates (Bett, 1960; Berry, 1963a; Smith, 1966; Hunter, 1968; Runham & Laryea, 1968; Parivar, 1978). Similar reproductive-cycle studies of tropical or sub-tropical snails such as M. abbreviatus are rare (Zannini, 1958; Galangau, 1964).
The use of M. abbreviatus (Gastropoda, Pulmonata) as an experimental animal model requires basic knowledge of its biology. Therefore, the aim of this research was to determine the reproductive cycle of this snail based on the annual variation of the morphological parameters of the reproductive organs and a histological analysis of the ovotestis.
MATERIAL AND METHODS
Adult specimens of the snail Megalobulimus abbreviatus (Pulmonata, Stylommatophora) (Bequaert, 1948) were collected in Charqueadas, Rio Grande do Sul State, Brazil (lat. 30º56'S; long. 51º40'W) every month from March 1991 to July 1992. The snails were then housed in humid cages at the Universidade Federal do Rio Grande do Sul, exposed to ambient temperature and natural light/dark cycles, and fed with lettuce ad libitum twice weekly. The mean ambient temperature was 21.6ºC in spring, 24.3ºC in summer, 16.7ºC in autumn, and 14.9ºC in winter.
Every month, 8 to 17 animals weighing 39.6-101.5 g were anesthetized by immersion in a menthol-saturated solution, following which the ovotestis, spermoviduct, oviduct-vagina, albumen gland, penis complex, Bursa copulatrix, and accessory gland were dissected and removed. Each was immediately isolated, washed with a Ringer solution (Jaeger, 1961), dried on filter paper, and weighed on a precision balance (Sartorius). The weight of each organ was classified by season. At the same time, three ovotestes were obtained every month, fixed in a Bouin's solution, and embedded in paraffin wax. Using a microtome (Minot), sagittal, frontal, or transverse serial sections (10 um) were cut and stained with hematoxylin-eosin (H.E.) as well as by using periodic acid-Schiff (P.A.S.) procedures (Pearse, 1968). Sections were examined and photographed with a Nikon Optiphot-2 microscope and measured with a micrometer ocular lens.
Organ weight was used to calculate the organosomatic index (O.S.I.) by the following formula: OSI = [(organ weight/body weight) ´ 100]. These indices were grouped by season and analysed by means of both one-way ANOVA and Duncan's multiple range test (p < 0.05). A new index was created, referred to by us as the oocytic maturation index (O.M.I.) = [(number of oocytes with diameter equal to or higher than 80 mm/ total number of oocytes) ´ 100], yielding the proportion of mature oocytes found each time. The 80 mm value was based on the diameter of mature oocytes previously found in M. oblongus (Tompa, 1984).
RESULTS
The O.S.I. showed some variation in different organs during the year (Table 1). Spring saw the highest values for all sexual accessory organs when compared to those for the other seasons. With the exception of the albumen gland, these values were significantly different from those found in the other seasons.
When compared to the other reproductive organs, the ovotestis in autumn showed an elevated O.S.I., which differed significantly from those in the other seasons (Table 1).
The ovotestis of M. abbreviatus, which appeared as a large, yellowish organ partially embedded in the digestive gland, is located in the last turn of the shell's apex. A hermaphroditic organ, it consists of four lobes, each of which presented a large number of acini or follicles intermingled with loose connective interacinar tissue. Each acinus was a pear-shaped sac (Fig. 1A) opening into an acinar duct. The ducts of several acini joined together formed the hermaphroditic duct. In each acinus four types of cells were identified: (a) male germ cells, mainly occupying the lumen of the acini; (b) female germ cells, initially found on the wall of the acini; (c) Sertoli cells; and (d) follicle cells. A fifth type of cell was occasionally observed among the others on the acinar wall, which, due to its position and characteristics, corresponds to the cells of the germinal epithelium as described for the ovotestis of other snails and slugs.

Although all the spermatogenetic forms appeared in the ovotestes throughout the year, there was a clear predominance of some forms in specific periods.
In August and September, acini are largely constituted of spermatogonia, and primary and secondary spermatocytes (Fig. 1B). Spermatogonia were large, round cells with a narrow, basophilic cytoplasm around the nucleus. The nucleus of this cell had a granular aspect with fine chromatin particles in which one nucleolus could be observed. These cells were arranged in clusters, each having a variable number of cells assembled around a Sertoli cell in which they were partially embedded.
The primary spermatocytes were similar to the spermatogonia. They were large cells forming clusters with a similar arrangement to that of spermatogonia. However, they presented a slightly eosinophilic cytoplasm, and the nucleus had several forms due to the morphological changes observed during the meiotic process phases. Secondary spermatocytes were small, rounded cells organized in clusters. The highly heterochromatic nucleus, smaller than those of the primary spermatocytes, was spherical in shape containing no observable nucleolus.
Two basic types of Sertoli cells lining the basal lamina of the acini were identified. One was an active Sertoli cell in which spermatogonia, spermatocytes, and spermatids were embedded. Such cells were observed in each acinus, occupying an area from the basal to the apical domain. This cell, attached broadly at one end to the acinar wall, had an irregular shape varying from squamous to columnar and often producing cytoplasmic extensions into the acinar lumen to which the male gametic cells adhered. The large coarsely granulated nucleus with deeply stained chromatin was spherical or oval, depending on cell shape. In contrast, hypertrophied Sertoli cells were found only at the base of some acini. These were large cells devoid of any germ cell, and sometimes occluding the acinar lumen. The cytoplasm of these cells had a foamy appearance with faint staining, while the nucleus presented an irregular shape, and sometimes became lobulated. The chromatin was concentrated and stained strongly with hematoxylin.
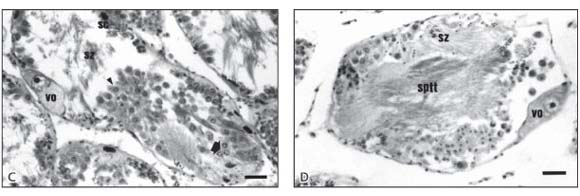
In October, November, and December the gonads exhibited the four stages of the male gametic cells: spermatogonia, spermatocytes I and II, and spermatids (Fig. 1C). The spermatids were organized in clusters enclosing a Sertoli cell and appeared to go through various differentiation stages, so that, for the sake of convenience, only two spermatid classes of cells were considered. When these cells, which appeared to be the same size as secondary spermatocytes, were round or oval and showed a small, condensed, and fairly polarized nucleus, they were considered to be early spermatids. Cells with an elongated cytoplasm and a tiny, round, clearly polarized nucleus were regarded as late spermatids. These were found at the apex of the Sertoli cells in the final stage of spermatogenesis, when the head and the tail became visible. At this stage their tails projected towards, and almost completely occupied, the entire acinar lumen.
In January and part of February, various acini showed numerous spermatozoa in their lumen. However, empty or emptying acini were also observed (Fig. 1D). In effect, in order to classify the male gametic cells, the spermatozoon was considered as an independent cell having a head, and containing a rod-like, condensed, highly heterochromatic nucleus, and a very long eosinophilic tail. Efferent or acinar ducts were full of spermatozoa during these months.
In March, April, May, and June the spermatozoa were abundant inside the acini lumen (Fig. 1E). In the last month a reduction of the spermatozoa mass occurred concurrently. During those months the acinar ducts always contained spermatozoa in their lumen.
Ovotestes fixed in July showed a significant decrease in the spermatozoa mass (Fig. 1F). However, spermatogonia and spermatocytes frequently occurred throughout this period while spermatids were scarce.
As observed in the male gametic cells, acini exhibited maturation of the oocytes throughout the year. Previtellogenic and vitellogenic oocytes were identified in all gonads examined each month. These cells rested on the acinar wall during the development period, attached by squamous follicular cells to the luminal surface. Previtellogenic oocytes (Fig. 1G) were elliptical or oval cells with basophilic cytoplasms, probably due to the large amount of ribosomal RNA synthesized. Their euchromatic germinal vesicles, which were usually located in a central position of the cell, were large and round and presented a prominent, spherical nucleolus. At this stage of oocyte development, lipid droplets and glycogen particles had not yet been identified.
Vitellogenic oocytes (Fig. 1H) were large, rounded cells, reaching a 120 µm diameter. The cytoplasm, which stained less with hematoxylin dye than than did the previtellogenic oocytes, had a foamy appearance, probably due to the accumulation of yolk (glycogen granules and lipid droplets). The oocyte cytoplasm showed a P.A.S. positive reaction. The nucleus of this cell was to a great extent eccentric and invariably showed a ubiquitous nucleolus. No oogonia or pre-meiotic oocytes were identified.
The oocyte diameter and O.M.I. showed some seasonal variation. In spring, the highest mean diameter of oocytes was significantly different from that found in the remaining seasons. In autumn the second highest value was obtained, being significantly different from that registered in summer (Table 2). The highest value for the O.M.I. was that of the spring, with no sizeable variation during the other seasons (Table 2).
DISCUSSION
The results of the histological analysis of the ovotestis suggest that M. abbreviatus can be considered a simultaneous hermaphroditic snail. As with other described stylommatophoran species, such as Arion ater (Lusis, 1961; Smith, 1966; Parivar, 1978), Deroceras reticulatum (Bailey, 1973; Runham, 1978), and Milax gagates (Galangau, 1964), the analysis of the ovotestes showed that both male and female sexual cells develop together.
Despite the lack of experimental evidence, the condition of the M. abbreviatus gonad when compared to what is available in data from the literature appears to indicate that this snail represents a perennial and iteroparous species, with more than one reproductive cycle during its life span. An indication of this is that the gonads of semelparous slugs and snails show that there is a strict correlation between the phase of the reproductive cycle and the set of gametogenic cells observed (Lusis, 1961; Smith, 1966; Runham & Laryea, 1968; Parivar, 1978). But this was not the case with M. abbreviatus.
In addition, the lack of a sign of senescence in the acini, such as a large number of hypertrophied Sertoli cells and the appearance of post-reproductive epithelium, would seem to confirm iteroparity in M. abbreviatus. Furthermore, the results obtained indicate that this species has an annual reproductive cycle, with one period related to the preparation of the reproductive system for breeding and another related to mating and egg laying. Moreover, a clear variation in the O.S.I. of reproductive organs can be observed, as well as an evident alteration of the gametic process throughout the four seasons. These modifications in stylommatophorans are related to changes from a reproductive to a non-reproductive state (Tompa, 1984).
The results of the O.S.I. suggest that spring is the principal reproductive season in the annual reproductive cycle of M. abbreviatus. The significant differences between the O.S.I. values of the reproductive organs obtained in that season is probably related to a number of processes linked to both mating and egg laying, such as the increase in the secretion processes, ova formation, and sperm nutrition. Evidence suggests that these kinds of modifications may happen in Semperula maculata in which the reproductive period of the cycle was characterized by a rise in the biochemical protein level as determining by assaying in its reproductive organs (Nanaware & Varute, 1976). Moreover, both the functioning and the state of the reproductive organs in the reproductive system of stylommatophorans appear strongly correlated with production of gametes by the ovotestis (Lusis, 1961; Berry, 1963a, b; Smith, 1966; Runham & Laryea, 1968). If this were true for M. abbreviatus, spring would be marked by the presence of ripe gametes in preparation for breeding. In fact, spermatozoa are found inside the acini of the ovotestis of M. abbreviatus after September. In the same way, spring is the season in which both the mean oocytic diameter and O.M.I. are highest, suggesting that there would be gametes available for reproductive activity. Besides, increases in the light/dark ratio and the successive temperature rises that characterize spring have a direct influence on the reproductive activity of other Pulmonates such as Helix aspersa (Gomot & Griffond, 1993; Gomot et al., 1989), stimulating mating and egg laying (Bailey, 1981; Enée et al., 1982).
Other evidence indicating spring as the main reproductive period for M. abbreviatus has come from the morphological analysis of the gonadotropic hormone-producing dorsal body. In M. oblongus the dorsal bodies obtained at the end of winter and beginning of spring showed a maximum weight, with their secretory cells having higher synthetic activity (Zancan & Achaval, 1995). An increment in the vitellogenetic process due to the action of the dorsal body hormone could explain the sudden increases in the mean oocytic diameter and O.M.I. observed from winter to spring. In addition, higher lipid levels were found in the ovotestis of M. oblongus in winter and spring (Da Silva & Zancan, 1994). Lipids are used for the production of gametes in the prosobranchs Littorina littoria (Williams, 1970), Haliotis cracheroidii (Webber, 1970), and Thais lamellosa (Voogt, 1983), and in the pulmonate Semperula maculata (Nanaware & Varute, 1976).
In summer there was a significant decrease in the O.S.I. for all the reproductive system organs of, except the ovotestis. This could be related with a reproductive activity reduction of the animal, as has been described for Semperula maculata (Nanaware & Varute, 1976). The ovotestis, on the other hand, did not show O.S.I. values significantly different from those observed in spring. This could be explained by the histological analysis of the gonads obtained in January and February, which showed a large number of spermatids and spermatozoa. The higher weight and volume of ovotestis, when crowded with these cells, were also observed for other stylommatophorans (Lusis, 1961; Smith, 1966; Parivar, 1978; Runham, 1978; Runham & Laryea, 1968).
The reduction of the mean oocytic diameter and of the O.M.I. obtained in the summer compared to the spring is probably due to the ovulation or degeneration of a great number of ripe oocytes at the end of spring and beginning of summer, and to a depression of the vitellogenetic process. Corroborative evidence is found in the result of an analysis of dorsal body cells of M. oblongus in summer, which showed reduced secretory activity (Zancan & Achaval, 1995).
Autumn, as well as spring, is a period characterized by pronounced copulation and egg laying in several species of snails and slugs dwelling in places having temperate or tropical climates (Bett, 1960; Lusis, 1961; Berry, 1963a; Galangau, 1964; Smith, 1966; Hunter, 1968; Runham & Laryea, 1968; Parivar, 1978). However, an analysis of the O.S.I. of M. abbreviatus, as well as histological and morphometrical data did not indicate autumn as a season in which reproductive activity is accentuated, since the O.S.I. values of the reproductive organs in autumn are similar to those observed in the summer. Furthermore, autumn like summer is characterized by a reduction in the photophase of the daily photoperiodic cycle. When Helix aspersa was submitted to a decreased light regime in a 24-hour light/dark cycle or a fixed short day (8 hours light: 16 hours dark), it presented a partial or complete reduction in reproductive activities such as mating, egg-laying, and ovulation (Bailey, 1981; Gomot et al., 1989).
In contrast to the values registered for the other reproductive organs of M. abbreviatus, the ovotestis showed a higher O.S.I. value in autumn. This is due to the presence of a large number of spermatids and spermatozoa inside the acini during the three months of this season. This accentuated spermatogenesis in a period not clearly related to reproduction may be related to storage of male gametes, to be used during breeding at the beginning of spring, in the seminal vesicle of the hermaphroditic duct. The function of the seminal vesicle in autosperm storage is well known in pulmonates (Runham, 1978; Geraert & Joosse, 1984; Lutchel et al., 1997).
The mean oocytic diameter observed in autumn is higher than that in summer, while the O.M.I. is very similar. These results indicate continued oocyte growth during the autumn and a partial unavailability of ripe oocytes during this season. Moreover, the gonadal lipids showed lower levels for M. oblongus in summer (Da Silva & Zancan, 1994). Since part of the lipids are found within the ovotestis as lipid yolk during the vitellogenic phase of oogenesis (Dohmen, 1983; Lutchel et al., 1997), a lower lipid level implies a reduced number of large vitellogenic oocytes.
The reproductive tract organs of M. abbreviatus in winter presented an O.S.I. similar to that of summer and autumn. When compared to the two preceding seasons, these values could indicate a lack of reproductive activity. Ovotestis O.S.I. values in winter were similar to those in spring and summer. Histological analysis of the ovotestis revealed that in July and August only spermatogonia, spermatocytes, and a few spermatozoa produced at the end of autumn and beginning of winter are found inside the acini. It is thought that spermatogenesis may suffer a block during these months, as that described for Helix aspersa and Helix pomatia (Lind, 1973; Bailey, 1981).
Given that the production of new spermatozoa occurs in October and mating is observed from September on, it is believed that the male gametes, stored in the seminal vesicle of the hermaphroditic duct and used in this reproductive event, arise from spermatogenesis occurring in autumn and at the onset of winter.
The mean oocytic diameter in winter is similar to that observed in autumn and lower when compared with that of spring. In contrast, the winter O.M.I. is higher than that of autumn, and increases to appreciable levels from June to September. In this period, the secretory dorsal body cells of M. oblongus show a cell diameter similar to that of autumn and a nuclear diameter like that of spring, indicating an increase in their synthetic activity (Zancan & Achaval, 1995). This suggests the maintenance or a mild elevation of the vitellogenetic process during this period, with which increase in the gonadal lipids of the ovotestis of M. oblongus during winter (Da Silva & Zancan, 1994) could as well be correlated. Thus, the increase in the O.M.I. toward the end of season could be related to an acceleration of the vitellogenetic process and the preparation of ripe oocytes for the spring breeding season.
The annual reproductive cycle of the subtropical snail M. abbreviatus appears to differ profoundly from the reproductive cycle of other iteroparous snails inhabiting temperate or tropical latitudes. Temperate-zone snails such as Helix pomatia and Helix aspersa have both clear hibernation periods with no reproductive activity during the year (autumn and winter) and a breeding period (Bailey, 1981; Lind, 1973). Otherwise, tropical species such as Gyliotrachela depressispira (Berry, 1963a) show continuous spermatogenesis during the year while oocyte diameters, female reproductive system maturation, and oviposition is strictly correlated with periods having increased precipitation (Duncan, 1975). In this way M. abbreviatus has a main period of breeding in spring and the beginning of summer and a separate period, occurring over the remainder of the summer until the end of the winter, to prepare the body for the next reproductive phase. Oogenesis and spermatogenesis are continuous throughout the year, with spermatozoa being produced almost year round, with possible interruption in winter, while great amounts of ripe oocytes occur only in spring.
Acknowledgements The authors are grateful to Dr. Roselis S. M. da Silva for making the Invertebrate Endocrinology Laboratory, in which part of this work was carried out, available to us. We also thank Fernando Capsi Pires and Antonio G. Severino for their technical assistance. This work was supported by CNPq and FAPERGS.
Received June 2, 2003 Accepted November 19, 2003 Distributed August 31, 2005
- BAILEY, T. G., 1973, The in vitro culture of reproductive organs of the slug Agriolimax reticulatus (müll.). Netherlands Journal of Zoology, 1: 72-85.
- BAILEY, S. E. R., 1981, Circannual and circadian rhythms in the snail Helix aspersa Müller and the photoperiodic control of annual activity and reproduction. Journal of Comparative Physiology, 142: 89-94.
- BEQUAERT, J. C., 1948, Monograph of the Strophocheilidae, a neotropical family of terrestrial mollusks. Bulletin: Museum of Comparative Zoology, 100: 1-210.
- BELLÓ-KLEIN, A., WHILHELMS, F. M., SOUZA, D. T., KUCHARSKI, L. C. & BELLÓ, A. A., 1993, Effect of hydrogen peroxide on the isolated heart of the snail Megalobulimus oblongus Medical Science Research, 21: 357-358.
- BERRY, A. J., 1963a, The genital system of the Malayan limestone hill snail Gyliotrachela depressispira, with notes on breeding. Proceedings of the Zoological Society of London, 141: 361-370.
- BERRY, A. J., 1963b, The anatomy of two Malayan limestone hill Streptaxidae, Sinoennea kanchingensis Tomlin and Oophana diaphanopepla Van Bentham Jutting, with special reference to the genital system. Proceedings of the Malacological Society of London, 35: 139-150.
- BETT, J. A., 1960, The breeding seasons of slugs in gardens. Proceedings of the Zoological Society of London, 135: 559-568.
- DA SILVA, R. S. M. & ZANCAN, D. M., 1994, Seasonal variation of the carbohydrate and lipid metabolism in a land pulmonate gastropod Megalobulimus oblongus Comparative Biochemistry and Physiology., 108A: 337-341.
- DE JORGE, F. C., CINTRA, C. A. B., HAESER, P. E. & SAWAYA, P., 1965, Biochemical studies on the snail Strophocheilus oblongus musculus (Becquaert). Comparative Biochemistry and Physiology., 14: 35-42.
- DOHMEN, M. R., 1983, Gametogenesis, pp. 1-48. In: A. S. M. Saleuddin & K. M. Wilbur (ed.), The Mollusca, 3. vol. Academic press, London.
- DONELLI, D. C., ZANCAN, D. M., FACCIONI-HEUSER, M. C. & ACHAVAL, M., 1998, Localization of acid phosphatase activity in the central nervous system of the pulmonate snail Megalobulimus oblongus. Brazilian Journal of Morphological Sciences, 15: 73-83.
- DUNCAN, C. J., 1975, Reproduction, pp. 309-365. In: V. Fretter & J. F. Peake (eds.), Pulmonates, 1. vol. Academic press, London.
- ENÉE, J., BONNEFOY-CLAUDET, R. & GOMOT, L., 1982, Effet de la photopériode artificielle sur la reproduction de l'Escargot Helix aspersa Müll. Comptes Rendus des Séances de L' Académie des Sciences Paris, 294: 357-360.
- FACCIONI-HEUSER, M. C., ZANCAN, D. M., LOPES, C. & ACHAVAL, M., 1999, The pedal muscle of the land snail Megalobulimus oblongus (Gastropoda, Pulmonata): an ultrastructural approach. Acta Zoologica, 80: 325-337.
- GALANGAU, V., 1964, Le cycle annuel de Milax gagates Drap. (Gasterópode, Pulmoné) et ses deux pontes. Bulletin d'Societie de Zoologie, France, 98: 510-513.
- GERAERTS, W. P. M. & JOOSSE, J., 1984, Freshwater snails (Basommatophora), pp. 141-207. In: A. S. M. Saleuddin & K. M. Wilbur (eds.), 7. vol. Academic press, London.
- GOMOT, P., GOMOT, L. & GRIFFOND, B., 1989, Evidence for a light compensation of the inhibition of reproduction by low temperatures in the snail Helix aspersa Ovotestis and albumen gland responsiveness to different conditions of photoperiods and temperatures. Biology of Reproduction, 40: 1237-1245.
- GOMOT, L. & GRIFFOND, B., 1993, Action of epigenetic factors on the expression of hermaphroditism in the snail Helix aspersa Comparative Biochemistry and Physiology, 104A: 195-199.
- HUNTER, P. J., 1968, Studies on slugs of arable ground, II. Life cycles. Malacologia, 6: 379-389.
- JAEGER, C. P., 1961, Action of acetylcholine on the heart of Strophocheilus oblongus Comparative Biochemistry and Physiology, 4: 30-32.
- JAEGER, C. P., 1965, Giant snail Is used for muscle studies. Natural History, 74: 26-27.
- LIND, H., 1973, The functional significance of the spermatophore and the fate of spermatozoa in the genital tract of Helix pomatia (Gastropoda: Stylommatophora). Journal of Zoology, London, 169: 39-64.
- LUSIS, O., 1961, Postembryonic changes in the reproductive system of the slug Arion ater rufus L. Proceedings of the Zoological Society of London, 137: 433-468.
- LUTCHEL, D. L., MARTIN, A. W., DEYUP-OLSEN, I. & BOER, H. H., 1997, Gastropoda: Pulmonata, pp. 624-661. In: F. W. Harrinson & A. J. Kohn (eds.), Microscopic Anatomy of Invertebrates, 6B vol. Wiley liss INC., New York.
- MARQUES, M. & PEREIRA, S., 1970, Seasonal variation in blood glucose and glycogen levels of some tissues of Strophocheilus oblongus (Mollusca, Gastropoda). Revista Brasileira. de Biologia, 30: 43-48.
- NANAWARE, S. G. & VARUTE, A. T., 1976, Biochemical studies on the reproductive organs of a land pulmonate, Semperula maculata (Templeton, 1858; Semper, 1885) During seasonal breeding-aestivation cycle: I. Biochemical seasonal variations in proteins and lipids. The Velinger, 19: 96-106.
- PARIVAR, K., 1978, A histological survey of gonadal development in Arion ater L. (Mollusca, Pulmonata). Journal of Molluscan Studies, 46: 139-147.
- PEARSE, A. G. E., 1968, Histochemistry. Theoretical and applied Churchill Livingstone, Edinburg.
- PITONI, V. L. L., VEITENHEIMER, I. L. & MANSUR, M. C. D., 1976, Moluscos do Rio Grande do Sul. Iheringia Divulgação, 5: 29-30.
- ROSSI, I. C. C. & DA SILVA, R. S. M., 1993, Effects of starvation and a carbohydrate-rich diet on glycogen metabolism in a gastropod mollusc, Megalobulimus oblongus. Comparative Biochemistry and Physiology, 106A: 831-836.
- ROMERO, S. M. B. & HOFFMANN, A., 1991, A technique for cardiac denervation in Megalobulimus sanctipauli (Mollusca, Gastropoda, Pulmonata). Brazilian Journal of Medical and Biological Research, 24: 1185-1188.
- ROMERO, S. M. B., HOFFMANN, A. & MENESCAL-DE-OLIVEIRA, L., 1994, Is there an opiate receptor in the snail Megalobulimus sanctipauli? Action of morphine and naloxone. Comparative Biochemistry and Physiology, 107C: 37-40.
- RUNHAM, N. W., 1978, Reproduction and its control in Deroceras reticulatum Malacologia, 17: 341-350.
- RUNHAM, N. W. & LARYEA, A. A., 1968, Studies on the maturation of the reproductive system of Agriolimax reticulatus (Pulmonata: Limacidae). Malacologia, 7: 93-108.
- SANTOS, P., GEHLEN, G., FACCIONI-HEUSER, M. C., ZANCAN, D. M. & ACHAVAL, M., 2002, Distribution of glial cells in the central nervous system of the pulmonate snail Megalobulimus oblongus identified by means of a glial fibrillary acidic protein marker. Acta Zoologica., 83: 345-351.
- SAWAYA, P. & PETERSEN, J. A., 1962, Sobre a ocorrência de Strophocheilidae (Molusco, Gastrópode) no Rio Grande do Sul. Faculdade de Filosofia, Ciências e Letras da Universidade de São Paulo (Boletim). Zoologia, 261: 31-41.
- SMITH, B. J., 1966, Maturation of the reproductive tract of Arion ater (Pulmonata:Arionidae). Malacologia, 4: 325-349.
- TOMPA, A. S., 1984, Land snails (Stylommatophora), pp. 47-140. In: A. S. M. Saleuddin & K. M. Wilbur (eds.), The Mollusca, 7. vol. Academic Press, London.
- VOOGT, P. A., 1983, Lipids: their distribution and metabolism., pp. 329-370. In: A. S. M. Saleuddin & K. M. Wilbur (ed.), The Mollusca, 1. vol. Academic Press, London.
- WEBBER, H. H., 1970, Changes in metabolite composition during the reproductive cycle of the Abalone Haliotis cracheroidii (Gastropoda: Prosobranchiata). Physiology and Zoology, 43: 213-231.
- WILLIAMS, E. E., 1970, Seasonal variations in the biochemical composition of the edible Winkle Littorina littorea (L.). Comparative Biochemistry and Physiology, 33: 655-661.
- ZANCAN, D. M., NOBLEGA, A. G., SEVERINO, A. G. & ACHAVAL, M., 1994, Acetylcholinesterase distribution in the central nervous system of the Megalobulimus oblongus (Gastropoda, Pulmonata). Archives D'Anatomie D'Histologie & D'Embriologie normales et expérimentales, 75: 75-86.
- ZANCAN, D. M. & ACHAVAL, M., 1995, Morphology of the dorsal body of the pulmonate mollusk Megalobulimus oblongus (Müller, 1774) and its sazonal variations. Revista Brasileira de Biologia, 55: 1-11.
- ZANCAN, D. M., BRAUER, M. & ACHAVAL, M., 1997, Monoamine-containing neurons in the central nervous system of Megalobulimus oblongus (Gastropoda, Pulmonata). Comparative Biochemistry and Physiology A., 118: 765-772.
- ZANNINI, A. M., 1958, Osservazioni sul ciclo annuale della gametogenesi maschile in Helix cincta Müll. Archivio Zoologico Italiano, 43: 283-303.
Correspondence to
Publication Dates
-
Publication in this collection
13 Dec 2005 -
Date of issue
Aug 2005
History
-
Accepted
19 Nov 2003 -
Received
02 June 2003