
Abstracts
Spatial autocorrelation is the lack of independence between pairs of observations at given distances within a geographical space, a phenomenon commonly found in ecological data. Taking into account spatial autocorrelation when evaluating problems in geographical ecology, including gradients in species richness, is important to describe both the spatial structure in data and to correct the bias in Type I errors of standard statistical analyses. However, to effectively solve these problems it is necessary to establish the best way to incorporate the spatial structure to be used in the models. In this paper, we applied autoregressive models based on different types of connections and distances between 181 cells covering the Cerrado region of Central Brazil to study the spatial variation in mammal and bird species richness across the biome. Spatial structure was stronger for birds than for mammals, with R² values ranging from 0.77 to 0.94 for mammals and from 0.77 to 0.97 for birds, for models based on different definitions of spatial structures. According to the Akaike Information Criterion (AIC), the best autoregressive model was obtained by using the rook connection. In general, these results furnish guidelines for future modelling of species richness patterns in relation to environmental predictors and other variables expressing human occupation in the biome.
spatial autoregression; species richness; Cerrado; birds; mammals
Autocorrelação espacial é definida como a falta de independência entre pares de observações a uma dada distância geográfica e é um fenômeno muito freqüente em dados ecológicos. É importante levar em consideração os efeitos de autocorrelação espacial em ecologia geográfica, tanto para realizar uma descrição mais detalhada dos dados quanto para corrigir estimativas enviesadas do erro Tipo I das análises estatísticas convencionais. Entretanto, para resolver efetivamente esses problemas, é preciso avaliar a melhor forma de incorporar estruturas espaciais nos modelos. Neste estudo, modelos autoregressivos, baseados em diferentes tipos de conexões e distâncias entre 181 células de uma rede cobrindo a região do Cerrado brasileiro, foram ajustados para avaliar a variação espacial de riqueza de mamíferos e aves dentro do bioma. A estrutura espacial foi ligeiramente mais forte para aves do que para mamíferos, com valores de R² variando entre 0,77 e 0,94 para mamíferos e 0,77 e 0,97 para aves, em modelos baseados em diferentes formas de conexão espacial. Segundo o Critério de Informação Akaike (AIC), o modelo autoregressivo melhor ajustado foi obtido através da conexão "em torre". Em geral, esses resultados fornecem diretrizes para futuras modelagens dos padrões de riqueza de espécies que estão associados a preditores ambientais e/ou a variáveis que expressam a ocupação humana no Cerrado.
autoregressão espacial; riqueza de espécies; Cerrado; aves; mamíferos
ECOLOGY
Autoregressive modelling of species richness in the Brazilian Cerrado
Modelagem por auto-regressão da riqueza de espécies no Cerrado Brasileiro
Vieira, CM.I, II,* * e-mail: diniz@icb.ufg.br, cleiber.marques@ueg.br ; Blamires, DI, III; Diniz-Filho, JAF.IV,* * e-mail: diniz@icb.ufg.br, cleiber.marques@ueg.br ; Bini, LM.IV; Rangel, TFLVB.V
IDepartamento de Biologia, Unidade Universitária de Ciências Exatas e Tecnológicas UnUCET, Universidade Estadual de Goiás UEG
IIPrograma de Pós-Graduação em Ciências Ambientais, Universidade Estadual de Goiás UEG
IIIUnidade Universitária de Quirinópolis, Universidade Estadual de Goiás UEG, Av. Brasil, Qd. 03, Lt. 01, s/n., Conjunto Hélio Leão, CEP 75860-000, Quirinópolis, GO, Brasil
IVDepartamento de Biologia Geral, Instituto de Ciências Biológicas ICB, Universidade Federal de Goiás UFG, CP 131, CEP 74001-970, Goiânia, GO, Brasil
VDepartamento de Ecologia e Biologia Evolutiva, University of Connecticut Storrs, CT 06269-3043, USA
ABSTRACT
Spatial autocorrelation is the lack of independence between pairs of observations at given distances within a geographical space, a phenomenon commonly found in ecological data. Taking into account spatial autocorrelation when evaluating problems in geographical ecology, including gradients in species richness, is important to describe both the spatial structure in data and to correct the bias in Type I errors of standard statistical analyses. However, to effectively solve these problems it is necessary to establish the best way to incorporate the spatial structure to be used in the models. In this paper, we applied autoregressive models based on different types of connections and distances between 181 cells covering the Cerrado region of Central Brazil to study the spatial variation in mammal and bird species richness across the biome. Spatial structure was stronger for birds than for mammals, with R2 values ranging from 0.77 to 0.94 for mammals and from 0.77 to 0.97 for birds, for models based on different definitions of spatial structures. According to the Akaike Information Criterion (AIC), the best autoregressive model was obtained by using the rook connection. In general, these results furnish guidelines for future modelling of species richness patterns in relation to environmental predictors and other variables expressing human occupation in the biome.
Keywords: spatial autoregression, species richness, Cerrado, birds, mammals.
RESUMO
Autocorrelação espacial é definida como a falta de independência entre pares de observações a uma dada distância geográfica e é um fenômeno muito freqüente em dados ecológicos. É importante levar em consideração os efeitos de autocorrelação espacial em ecologia geográfica, tanto para realizar uma descrição mais detalhada dos dados quanto para corrigir estimativas enviesadas do erro Tipo I das análises estatísticas convencionais. Entretanto, para resolver efetivamente esses problemas, é preciso avaliar a melhor forma de incorporar estruturas espaciais nos modelos. Neste estudo, modelos autoregressivos, baseados em diferentes tipos de conexões e distâncias entre 181 células de uma rede cobrindo a região do Cerrado brasileiro, foram ajustados para avaliar a variação espacial de riqueza de mamíferos e aves dentro do bioma. A estrutura espacial foi ligeiramente mais forte para aves do que para mamíferos, com valores de R2 variando entre 0,77 e 0,94 para mamíferos e 0,77 e 0,97 para aves, em modelos baseados em diferentes formas de conexão espacial. Segundo o Critério de Informação Akaike (AIC), o modelo autoregressivo melhor ajustado foi obtido através da conexão "em torre". Em geral, esses resultados fornecem diretrizes para futuras modelagens dos padrões de riqueza de espécies que estão associados a preditores ambientais e/ou a variáveis que expressam a ocupação humana no Cerrado.
Palavras-chave: autoregressão espacial, riqueza de espécies, Cerrado, aves, mamíferos.
1. Introduction
Autocorrelation is the lack of independence between pairs of observations at given distances in time or space and is commonly found in ecological dataset (Legendre, 1993; Legendre and Legendre, 1998; Fortin and Dale, 2005). Many recent papers have discussed the importance of spatial autocorrelation when evaluating problems in geographical ecology, including gradients in species richness (Badgley and Fox, 2000; Lennon, 2000; Jetz and Rahbek, 2001; Rahbek and Graves, 2001; Diniz-Filho et al., 2003; Tognelli and Kelt, 2004). These papers show that autocorrelation analyses can be useful to provide a more detailed description of spatial structure in species richness data and to allow a better understanding of ecological processes driving richness (Legendre, 1993; Diniz-Filho et al., 2003). At the same time, it is now widely recognized that testing statistical hypotheses using standard methods (e.g., ANOVA, correlation and regression) in the presence of spatial autocorrelation will cause downward bias in the standard errors and, consequently, Type I error rates may be strongly inflated (Haining, 1990, 2003; Cressie, 1993; Legendre, 1993; Fortin and Dale, 2005).
Description of spatial patterns in data using correlograms and variograms is now straightforward (see Legendre and Legendre, 1998; Fortin and Dale, 2005). On the other hand, incorporating the autocorrelation structure into modelling, in a regression framework, may be a more complicated task. Autocorrelation analysis must be based on the spatial relationship between spatial units, but this must be established by taking into account the relationship between the processes underlying diversity and the geographic distances or connectivity among the spatial units analysed. For example, in a stream network, it is important to consider the links between units along the river flows and to take into account ecological barriers (Ganio et al., 2005). Formally, these alternative propositions must be codified into a weighting matrix W. However, for broad-scale patterns in species richness in terrestrial systems, it is difficult to establish these connections assuming spatial dynamics of ecological or biogeographical processes. Empirical evaluation of alternative spatial modelling strategies may be an initial solution, especially considering that models can be sensitive to misspecifications in the W matrix (Cressie, 1993).
In this paper, we evaluated the spatial patterns of mammal and bird species richness in the Brazilian Cerrado. Our goal is to discuss how changes in the definition of the spatial relationship among spatial units (i.e., grid cells) affect the statistical performance of the autoregressive models describing species richness. This may provide a basis for further analyses investigating the relationship between the environmental predictors and richness and consequently allow a better evaluation of the processes driving the spatial patterns in species richness.
2. Material and Methods
2.1. Data
The extents of occurrence of the 138 non-volant mammals species (Marinho-Filho et al., 2002) and 751 birds species (Ridgely and Tudor, 1989, 1994; del Hoyo et al., 1992; 1994; 1996; 1997; 1999; 2001; 2002; Junniper and Parr, 1998; Silva, 1995) found in the Brazilian Cerrado were mapped with a spatial resolution of 1º grid cell, with a total of 181 cells covering the Cerrado Biome (Figure 1). The gathered information on the mammals included 8 orders: Didelphimorphia, Xenarthra, Primates, Carnivora, Rodentia, Perissodactyla, Artiodactyla and Lagomorpha, respectively. Data from the literature (Marinho-Filho et al., 2002; Eisenberg and Redford, 1999; Emons, 1990; Embrapa, 2002; Fonseca et al., 1996) and specifically the following biodiversity websites were used to map the species: The Revista Brasileira de Zoologia (RBZ) site, SpeciesLink site, The Animal Diversity Web site (The University of Michigan Museum of Zoology) and the Site of the Global Biodiversity Information Facility Data Portal (GBIF) (a detailed species list and references are available from the authors upon request). A binary matrix was constructed by recording the geographic ranges of which species overlapped each cell, and species richness was calculated by summing the species present in the cells. Geographical coordinates of cell centroids (latitude and longitude) were also obtained for further spatial analyses.
2.2. Spatial description
Spatial autocorrelation measures the similarity between samples for a given variable as a function of spatial distance (see Legendre and Legendre, 1998). For quantitative variables, such as species richness, the Moran's I coefficient is the most commonly used coefficient in univariate autocorrelation analyses and is given by:

where n is the number of cells, yi and yj are the values of the species richness in cells i and j,  is the average of y and wij is an element of the matrix W. In this matrix, wij = 1 if the pair i,j of cells is within a given distance class interval (indicating cells that are "connected" in this class), and wij = 0 otherwise. S indicates the number of entries (connections) in the W matrix. The value expected under the null hypothesis of absence of spatial autocorrelation is 1/(n1). Detailed descriptions of the computations of the standard error of this coefficient are given in Legendre and Legendre (1998).
is the average of y and wij is an element of the matrix W. In this matrix, wij = 1 if the pair i,j of cells is within a given distance class interval (indicating cells that are "connected" in this class), and wij = 0 otherwise. S indicates the number of entries (connections) in the W matrix. The value expected under the null hypothesis of absence of spatial autocorrelation is 1/(n1). Detailed descriptions of the computations of the standard error of this coefficient are given in Legendre and Legendre (1998).
Moran's I usually varies between 1.0 and 1.0, for maximum negative and positive autocorrelation, respectively. Non-zero values of Moran's I indicate that richness values in cells connected at a given geographic distance are more similar (positive autocorrelation) or less similar (negative autocorrelation) than expected for randomly associated pairs of cells. The geographic distances among cell centroids can be partitioned into discrete classes, creating then successive W matrices and allowing computation of different Moran's I values for the same variable. This allows one to evaluate the patterns of autocorrelation as a function of spatial distance, in a graph called spatial correlogram, which furnishes a spatial description of the species richness. The number and definition of distance classes to be used in the correlograms are arbitrary, but a general methodological criterion is to try to maximise the similarity in the S values (number of connections) for the different Moran's I coefficients, so that they are more comparable. In this paper, correlograms were based on 15 distance classes (see Figure 4).
2.3. Spatial modelling
Spatial autocorrelation in mammal and bird richness (y) was modelled by an autoregressive model of the form:

where W is the row-standardized weighting matrix (not decomposed as in the correlogram), ρ is the autoregressive parameter and ε is the error vector. This model must be fitted by maximum likelihood procedures (Haining, 1990; 2003; Cressie, 2003). Squared correlation between y and the estimated value (ρWy) furnishes the pseudo-R2 of the model, expressing the proportion of variance in Y that is explained by an autoregressive process.
The autoregressive model above was fitted using various W matrices, derived from alternative ways to establish relationships between the spatial units (cells in the Cerrado grid). First, geographic distances among the cell centroids was used, and values in the matrix W were obtained using inverse-powered functions, given by:

where Dij is the geographic distance between centroids of cells i and j. Values of α ranging from 1 to 5, with steps of 0.5, were tested. Large α values indicate that large distances have relatively smaller weighting to model autocorrelation in species richness.
We also used seven different criteria to create binary (0 or 1) matrices W, indicating whether pairs of localities are connected or not. We used the Delaunay triangulation, Gabriel, the Minimum Spanning Tree and the Relative Neighbour networks, and established rook and queen connections among the cell centroids (Figure 2; see also Legendre and Legendre, 1998; Fortin and Dale, 2005, for details). These connections are built using different criteria to establish the links among the cells. In short, according to the Delaunay Triangulation Criterion, for a triplet of points (i.e., cell centroids) to be connected, a circle that circumscribes them (i.e. the circle passing through the three points) must include no other points, whereas in Gabriel connections, two points are connected if the circle in which the diameter is the distance between the points includes no other points. According to the Relative Neighborhood Criterion, two points are connected if, and only if, there is no other points lying on the intersection between the two circles centered in the two points, whereas in the Minimum Spanning Tree all points are inter connected so that the length of this connection is the minimum possible. Rook and Queen connections are designed to match chess' movements (see Figure 2).
The autoregressive models based on these 15 different matrices W (6 binary connectives and distance-based using 9 values of α) were compared using different approaches, for mammal and bird richness. The R2 values of the autoregressive model indicate the ability of each model to explain spatial structure in richness, whereas an autocorrelation analysis base on Moran's I in the ε term indicates the effectiveness in taking autocorrelation structure into account. Akaike information criterion (AIC) was also used to select the best model, within an information theory framework (see Burham and Anderson, 2002, for details). For each model, AIC corrected for small samples was computed as:
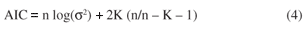
where n is the number of cells, K is the number of parameters in the model and σ2 is the variance of the residuals of each regression model. The variance of the residuals was used here as a proxy for the likelihood of the model given the data (Haining, 2003), whereas the term (n/n K 1) is the small sample correction term and tends to one as n increases. We compared the AIC values of each model using ΔAIC, which is the difference between AIC of each model and the minimum AIC found. A value higher than 10 indicates that a model has a poor fit relative to the best model, whereas a value less than 3 indicates that a model is equivalent to the best model (with the lowest AIC); model. The ΔAIC values were also used to compute Akaike's weighting of each model (w), which provides evidence that the model is actually the best explanatory model. The values of w are usually standardized by their sum among all models evaluated, so they are dependent on the set of models used and are given by:

All spatial analyses were performed in SAM (Spatial Analysis in Macroecology; Rangel et al., 2006), which is a software freely available at www.ecoevol.ufg.br/sam.
3. Results
Both mammal and bird species richness show a clear spatial pattern in the Brazilian Cerrado, with higher richness concentrated in the south-eastern region of the biome, decreasing toward the north (Figure 3). High richness values also appear in the western region of the biome, but this patch is clearer for mammals. Indeed, spatial correlogram confirm this strong spatial structure, with Moran's I coefficients large in the first distance class (0-245 km) and decreasing monotonically with the increasing of geographic distances (Figure 4).
Autoregressive modelling based on the different W matrices (Table 1) reveals a large variation in model fit, both for mammals and birds. As expected, connections based on the minimum spanning tree were not adequate and showed a very poor fit, and will be not considered further. The R2 values ranged from 0.77 to 0.94 for mammals and from 0.77 to 0.97 for birds. Relatively high values of Moran's I (i.e., I > 0.1) remain in the residuals of a few models. In these cases, modelling was not effective in taking the autocorrelation structure into account. In principle, models based on binary connections are better than models based on the inverse of geographic distances.
The AIC analysis allowed a more effective comparison among these alternative models (Table 1). In both mammals and birds, there is no model with ΔAIC smaller than 3, indicating that, in principle, there is a unique solution for modelling richness. The best models were obtained using the rook connection (Figure 2), and the standardized Akaike weights suggest a chance higher than 99.9% that these are the best models among those tested. They yield R2 values of 0.938 and 0.972 for mammals and birds, respectively. Coherent with patterns revealed in the spatial correlograms, birds display stronger spatial structure than mammals, with higher fit of autoregressive models.
However, it is interesting to note that Moran's I in the best model residuals for mammals displays a relatively high negative autocorrelation value 0.193, so a slight over-correction of the spatial structure probably occurred in this case (see Griffith, 2002). For both mammals and birds, the second best models were based on the Gabriel network (Figure 2), although ΔAIC is slightly larger than 10, indicating a low chance that this is the best model and, for mammals, the residual autocorrelation is still relatively high (0.143).
4. Discussion
Different forms of autoregressive models have been recently applied in geographical ecology (Lichstein et al., 2002; Kelt and Tognelli, 2004; Fortin and Dale, 2005). These models have been mainly used as a way to take the spatial structure into account in data and, at the same time, to evaluate how different environmental predictors are related to spatial variations in species richness. However, in most of these papers, researchers assume a given form of matrix W and do not explore alternative scenarios for the relationship among spatial units and the weighting of autoregressive model.
Our results show that the autoregressive model, used here only to analyse spatial structure in richness, is rather sensitive to variations in the definition of W and, consequently, using these models to relate richness to environmental predictors requires more effort around the definition of W.
We recognize that it is difficult to find theoretical arguments to support the use of a given W matrix to evaluate richness patterns at broad scales, so empirical evaluations, as performed here, are important. In our analysis of the mammals and birds in the Cerrado, AIC-based model selection was which effective in establishing a single model as the best one, based on a rook connection among cells. A model based on Gabriel connections followed this, according to AIC. For mammals, although AIC selected the rook connection as the best model, it is important to consider that a negative autocorrelation in the residuals remains, so a more careful evaluation should be performed. In practice, the consequence is that the effect of environmental predictors would be underestimated due to the overestimation of the spatial component in species richness. Of course, more complex models could be tested using alternative scenarios, for example taking into account the different biogeographical or ecological boundaries based on vegetation types or historical barriers, but this is beyond the scope of this paper.
We provide here guidelines for a more effective modelling of the richness patterns of mammals and birds in the Brazilian Cerrado. In some sense, it matches the theoretical evaluation by Griffith (1996), who gave some advice on choosing between alternative W matrices. For predictive purposes, using any of the models discussed here is better than assuming that no spatial autocorrelation exists, especially considering the relative high R2 values of all autoregressive models. This indicates that ignoring spatial components using ordinary regression models (OLS) probably will furnish biased results. Another important guideline is the preference of using low order (= short distance) expressions of spatial structure, in which close localities are more heavily weighted, when compared to distant ones. Indeed, our results show that the AIC values based on connections are usually higher than those obtained with the inverse distances functions. Also, there is a perfect decrease in AIC values when increasing the α values. However, Griffith (1996) indicated that over-specification of the connections in the model (in this case, generating high negative autocorrelation in the residuals of the model) is worse than under-specification. Thus, although rook connections were selected by AIC as the best model, followed by Gabriel connections, future modelling must be aware of inflated negative spatial structures in residuals and, eventually, further models may be tested if more complex autoregressive models are to be obtained (i.e., by adding environmental predictors see below).
Finally, overall spatial patterns of species richness in the Cerrado are not well established for most groups, but there seems to be a general decrease from south to north, that is more accentuated in amphibians than the decrease in bird and mammal richness shown in this paper (see Diniz-Filho et al., 2005a,b). There is currently a consensus that climatic variables, mainly the effects of available energy, water and increasing productivity, explain most of the variation in species richness of endothermic organisms at broad spatial scales (Allen et al., 2002; Hawkins et al., 2003; Currie et al., 2004). At the same time, correlations between richness and components of human occupation can be a consequence of this same energetic response (Balmford et al., 2001; but see Araújo, 2003) or may reflect patterns of knowledge of biodiversity (Diniz-Filho et al., 2005a). In both cases, the pattern has important consequences for biodiversity conservation. Although overall geographic distributions of mammals and birds are relatively well known, their patterns in the Brazilian Cerrado might still reflect knowledge effects, as previously shown for anurans (Diniz-Filho et al., 2005a), due to many reasons. On the one hand, geographic ranges used here, expressed as extents of occurrence, are overestimated because of the high rate of habitat loss in the biome. On the other hand, there may be biases in the estimation of these geographic ranges (and even in the very species richness), mainly because of the paucity of knowledge in the northern part of the biome. Despite these problems, the results obtained here are important because they furnish clear guidelines for future modelling of richness patterns in relation to environmental predictors and other variables expressing human occupation in the biome.
Acknowledgements Financial support for this study came from a PRONEX program of CNPq and SECTEC-GO (proc. 23234156). Work by JAFDF and LMB was also partially supported by other CNPq projects (grants number 300762/94-1 and 300367/96-1; respectively). CAPES and FUNAPE-UFG have also continuously supported our research program in macroecology and biodiversity.
Received March 24, 2006
Accepted May 30, 2007
Distributed May 31, 2008
- ALLEN, AP., BROWN, JH. and GILLOOLY, JF., 2002. Global biodiversity, biochemical kinetics, and the energetic-equivalence rule, Science, vol. 297, no. 5586, p. 1545-1548.
- ARAÚJO, MB., 2003. The coincidence of people and biodiversity in Europe, Global Ecol. Biogeogr., vol. 12, no. 1, p. 5-12.
- BADGLEY, C. and FOX, DL., 2000. Ecological biogeography of North American mammals: species density and ecological structure in relation to environmental gradients, J. Biogeography, vol. 27, no. 6, p. 1437-1467.
- BALMFORD, A., MOORE, JL., BROOKS, T., BURGESS, N., HANSEN, LA., WILLIAM, P., and RAHBEK, C., 2001. Conservation conflicts across Africa, Science, vol. 291, no. 5513, p. 2616-2619.
- BURHAM, KP. and ANDERSON, DR., 2002. Model selection and multimodel inference. A practical information-Theoretical Approach New York: Springer-Verlag.
- CRESSIE, NAC., 1993. Statistics for spatial data New York: John-Wiley and Sons, Inc.
- CURRIE, DJ., MITTELBACH, GG., CORNELL, HV., FIELD, R., GUEGAN, JF., HAWKINS, BA., KAUFMAN, DM., KERR, JT., OBERDORFF, T., O'BRIEN, E. and TURNER, JRG., 2004. Predictions and tests of climate-based hypotheses of broad-scale variation in taxonomic richness. Ecol. Lett., vol. 7, no. 12, p. 1121-1134.
- Del HOYO, J., ELLIOT, A. and SARGATAL, J. (eds.), 1992, Handbook of the birds of the world: ostrichs to ducks. vol. 01. Barcelona: Lynx Edicions.
- -, Handbook of the birds of the world: new world vultures to guineafowl. vol. 02, Barcelona: Lynx Edicions
- -, 1996, Handbook of the birds of the world: hoatzin to auks. vol. 03, Barcelona: Lynx Edicions.
- -, 1997, Handbook of the birds of the world: sandgrouse to cuckoos. vol. 04, Barcelona: Lynx Edicions.
- -, 1999, Handbook of the birds of the world: barn-owls to hummingbirds. vol. 05, Barcelona: Lynx Edicions.
- -, 2001, Handbook of the birds of the world: mousebirds to hornbills. vol. 06, Barcelona: Lynx Edicions.
- -, 2002, Handbook of the birds of the world: mousebirds to woodpeckers. vol. 07, Barcelona: Lynx Edicions.
- DINIZ-FILHO, JAF., BINI, LM., PINTO, MP., RANGEL, TFLVB., CARVALHO, P., and BASTOS, RP., 2006. Anuran species richness, complementarity and conservation conflicts in Brazilian, Cerrado, Acta Oecol, vol. 29, no. 1, p. 9-15.
- DINIZ-FILHO, JAF., BASTOS, RP., RANGEL, TFLVB., BINI, LM., CARVALHO, P. and SILVA, RJ., 2005. Macroecological correlates and spatial patterns of anuran description dates in the Brazilian Cerrado, Global Ecol. Biogeogr., vol. 14, no. 5, p. 469-477.
- DINIZ-FILHO, JAF., BINI, LM. and HAWKINS, BA., 2003. Spatial autocorrelation and red herrings in geographical ecology, Global Ecol. Biogeogr., vol. 12, no. 1, p. 53-64.
- FORTIN, MJ. and DALE, M., 2005. Spatial analysis: a guide for ecologists Cambridge: Cambridge University Press.
- GANIO, LM., TORGERSEN, CE. and GRESSWELL, RE., 2005. A geostatistical approach for describing patterns in stream networks, Front. Ecol. Environ., vol. 3, no. 3, p. 138-144.
- HAINING, R., 2003. Spatial data analysis. Theory and Practice. Cambridge: Cambridge University Press.
- HAINING, R., 1990. Spatial data analysis in the social and environmental sciences Cambridge: Cambridge University Press.
- HAWKINS, BA., PORTER, EE. and DINIZ-FILHO, JAF., 2003. Productivity and history as predictors of the latitudinal diversity gradient of terrestrial birds, Ecology, vol. 31, no. 6, p. 1608-1623.
- JETZ, W. and RAHBEK, C., 2001. Geometric constraints explain much of the species richness pattern in African birds, P. Natl. Acad. Sci.-Biol., vol. 98, no. 10, p. 5661-5666.
- JUNNIPER, T. and PARR, M., 1998. Parrots: a guide to the birds of the world. London: Yale University Press.
- LEGENDRE, P. and LEGENDRE, L., 1998. Numerical ecology Amsterdam: Elsevier.
- LEGENDRE, P., 1993. Spatial autocorrelation: trouble or new paradigm? Ecology, vol. 74, no. 6, p. 1659-1673.
- LENNON, JJ., 2000. Red-shifts and red herrings in geographical ecology, Ecography, vol. 23, no. 1, p. 101-113.
- LICHSTEIN, JW., SIMONS, TR., SHRINER, SA. and FRANZREB, KE., 2002. Spatial autocorrelation and autoregressive models in ecology, Ecol. Monog., vol. 72, no. 3, p. 445-463.
- MARINHO-FILHO, J., RODRIGUES, FHG., and JUAREZ, KM., 2002. The Cerrado mammals: diversity, ecology, and natural history. In OLIVEIRA, PS. and MARQUIS, RJ (eds.), The Cerrados of Brazil New York: Columbia University Press. p. 266-284.
- RAHBEK, C. and GRAVES, GR., 2001, Multiscale assessment of patterns of avian species richness, P. Natl. Acad. Sci.-Biol., vol. 98, no. 8, p. 4534-4539.
- RIDGELY, R. and TUDOR, G., 1989. The birds of South America (vol I- the oscine passerines). Austin: University of Texas Press.
- -, 1994, The birds of South America (vol. II- the suboscine passerines). Austin: University of Texas Press.
- SILVA, JMC., 1995. Birds of the cerrado region, South America, Steenstrupia, vol. 21, no. 1, p. 69-92.
- TOGNELLI, MF. and KELT, DA., 2004. Analysis of determinants of mammalian species richness in South America using spatial autoregressive models, Ecography, vol. 27, no. 4, p. 427-436.
Publication Dates
-
Publication in this collection
21 July 2008 -
Date of issue
May 2008
History
-
Received
24 Mar 2006 -
Accepted
30 May 2007





