Abstracts
The aim of this study was to describe and classify the food preference of fish species in Passa Cinco stream. The grade of feeding preference was applied to stomachs considered replete. This method consists of attributing values to food items found in certain species, according to the participation of each item in the analysed stomach. We analysed 576 full stomachs of 28 species. The autochthonous insects were the main constituents of the diets of these species, and the majority of ingested items classified as occasional. Allochthonous items such as plant debris, seeds and earthworms were associated with higher-order site. Of the total possible combination pairs of species, 29.4% showed high overlap, wich occurred mainly within species that consumed aquatic insect larvae. However, those species showed significant differences in the exploitation of food resources. Omnivory was common, showing the plasticity of the required species that inhabit environments as found in streams.
food preference; seasonality; spatial variation; diet overlap
O objetivo do presente trabalho foi descrever e classificar a preferência alimentar das espécies de peixes do rio Passa Cinco. O grau de preferência alimentar foi aplicado aos estômagos considerados repletos. Este método consiste em atribuir valores aos itens alimentares de uma dada espécie de acordo com a participação relativa de cada item no estômago analisado. Foram analisados 576 estômagos repletos de 28 espécies. Insetos de origem autóctone foram os principais constituintes das dietas dessas espécies, sendo a maioria dos itens ingeridos classificada como ocasional. Itens alóctones como fragmentos vegetais, sementes e minhocas estiveram associados ao ponto de maior ordem. Dentre os possíveis pares de combinações de espécies, 29,4% apresentaram alta sobreposição alimentar, que ocorreu principalmente devido ao consumo de formas imaturas de insetos, no entanto, foram detectadas diferenças significativas na exploração dos recursos por essas espécies. A onivoria foi frequente, mostrando a plasticidade trófica necessária às espécies que habitam ambientes como os encontrados nos riachos.
preferência alimentar; sazonalidade; variação espacial; sobreposição de dieta
BIOLOGY
Diet of fishes in Passa Cinco stream, Corumbataí River sub-basin, São Paulo state, Brazil
Dieta das espécies de peixes no rio Passa Cinco, sub-bacia do rio Corumbataí, São Paulo, Brasil
Rondineli, G.I, * * e-mail: giulianna.rondineli@gmail.com ; Gomiero, LM.II; Carmassi, AL.II; Braga, FMS.II
IDepartamento de Produção Vegetal, Centro de Ciências Agrárias, Universidade Federal do Espírito Santo - UFES, Alto Universitário, s/n, Guararema, CP 16, CEP 29500-000, Alegre, ES, Brazil
IIDepartamento de Zoologia, Instituto de Biociências, Universidade Estadual Paulista "Júlio de Mesquita Filho" - UNESP, Av. 24-A, 1515, CP 199, CEP 13506-900, Rio Claro, SP, Brazil
ABSTRACT
The aim of this study was to describe and classify the food preference of fish species in Passa Cinco stream. The grade of feeding preference was applied to stomachs considered replete. This method consists of attributing values to food items found in certain species, according to the participation of each item in the analysed stomach. We analysed 576 full stomachs of 28 species. The autochthonous insects were the main constituents of the diets of these species, and the majority of ingested items classified as occasional. Allochthonous items such as plant debris, seeds and earthworms were associated with higher-order site. Of the total possible combination pairs of species, 29.4% showed high overlap, wich occurred mainly within species that consumed aquatic insect larvae. However, those species showed significant differences in the exploitation of food resources. Omnivory was common, showing the plasticity of the required species that inhabit environments as found in streams.
Keywords: food preference, seasonality, spatial variation, diet overlap.
RESUMO
O objetivo do presente trabalho foi descrever e classificar a preferência alimentar das espécies de peixes do rio Passa Cinco. O grau de preferência alimentar foi aplicado aos estômagos considerados repletos. Este método consiste em atribuir valores aos itens alimentares de uma dada espécie de acordo com a participação relativa de cada item no estômago analisado. Foram analisados 576 estômagos repletos de 28 espécies. Insetos de origem autóctone foram os principais constituintes das dietas dessas espécies, sendo a maioria dos itens ingeridos classificada como ocasional. Itens alóctones como fragmentos vegetais, sementes e minhocas estiveram associados ao ponto de maior ordem. Dentre os possíveis pares de combinações de espécies, 29,4% apresentaram alta sobreposição alimentar, que ocorreu principalmente devido ao consumo de formas imaturas de insetos, no entanto, foram detectadas diferenças significativas na exploração dos recursos por essas espécies. A onivoria foi frequente, mostrando a plasticidade trófica necessária às espécies que habitam ambientes como os encontrados nos riachos.
Palavras-chave: preferência alimentar, sazonalidade, variação espacial, sobreposição de dieta.
1. Introduction
The feeding habits of fish communities in streams have been addressed in some river basins in Brazil (Castro and Casatti, 1997; Araújo et al., 2001, Castro et al., 2004). These environments are most affected by human activities when compared to other larger freshwater systems such as rivers and lakes. Furthermore, streams are environments subject to periodic disturbances (Gordon et al., 1995), especially during the rainy season, when a large amount of water, resulting from heavy rainfall, is received in a short time, increasing the water level and changing the conditions of existing habitats.
The removal of riparian vegetation, the use of pesticides on crops and uses of marginal resources, such as sand and water itself, causes a serious risk for the ichthyofauna even before they are known and studied (Esteves and Aranha, 1999). The small size (Castro, 1999) and other attributes such as lack of swimbladder, presence of single gonad and thorns for anchor (Braga, 2004) are conspicuous features of fish species in streams that enable species to adapt in these spatially limited environments.
According to Vannote et al. (1980), the increases in community diversity occur with the increase of orders in lotic water bodies: in streams of small orders, allochthonous resources prevail; on the other hand, in the biggest rivers, autochthonous resources prevail. Somehow the items depend indirectly on the native allochthonous material for survival. Thus there is an intricate food web of weak and interdependent links (Alvim and Peret, 2004). Braga and Gomiero (2009) pointed out that for many species that inhabit mountain streams in the watershed of the Ribeirão Grande, in the eastern Serra da Mantiqueira, the use of autochthonous food items were common, especially larvae and nymphs of insects. However, the contribution of allochthonous items from riparian forest is indirect by serving as an energy source for heterotrophic organisms as larvae and nymphs of insects, which serve as food for fish (Uieda and Motta, 2007).
Insectivory seems to be common to most stream fishes both for those with diurnal or nocturnal habits and also for those benthic or nektonic. However, the plasticity of the opportunistic species can ensure survival in severe conditions found in these types of environments (Teixeira et al., 2005). Another important factor is how the food item is presented in the environment (fixed, buried or mobile) and the ability of the fish to catch and eat those items which enable resources division and reduce competition (Braga et al., 2009). The surrounding landscape has great influence on both quality and quantity of food items available to these fish communities (Gomiero et al., 2008). Knowledge of these food chains is of paramount importance for the understanding of these fragile environments still little known and relatively preserved.
The aim of this work was to describe and classify the food preference of diferent species in a community of fish in Passa Cinco stream, belonging to the Corumbataí River sub-basin, a member of the Paraná River basin, São Paulo state.
2. Material and Methods
This work was accomplished in Passa Cinco stream, which is one of the main rivers of the Corumbataí River sub-basin. It presents 525 km2 of drainage area and its course covers about 60 km, from its headwater in serra da Cachoeira, a component of the complex of serra de Itaqueri, located in the local authority of Itirapina, with an altitude of 1,000 m; until its mouth in the Corumbataí river, with an altitude of 480 m. Nowadays, 51.72% of its surface is occupied by planted pasture area, 14.13% by sugarcane plantations, 15.67% by native forest and 0.74% by scrubland (Valente and Vettorazzi, 2004).
Six samplings were accomplished in five different points in Passa Cinco stream, contemplating sites of order two, three, four, five and six (with the following geographical coordinates: site 1 (order 2) - 22° 23' 36'' S and 47° 53' 08" W, site 2 (order 3) - 22° 22' 10" S and 47° 51' 22" W, site 3 (order 4) - 22° 21' 63" S and 47° 48' 48" W, site 4 (order 5) - 22° 24' 74" S and 47° 43' 34" W and site 5 (order 6) - 22° 30' 97" S and 47° 39' 49" W) in May, July, September and November of 2005 and January and March of 2006 (Figure 1), contemplating the drought and rainy seasons.
The sampling sites were characterised according to the predominant substrate type, riparian vegetation, degree of shading, type of current and mean width as shown by Rondineli and Braga (2009).
A sieve, an electric fishery equipment, gill nets and fish-traps were used as fishery equipment. A sieve was used on bank vegetation for ten times in each sample site (except for sample site 1, due to the local characteristics: lack of marginal vegetation and low water volume). Electric fishery equipment was used in sample sites 1, 2 and 3. In these places, the passages of 50 m were determined, where the equipment was placed for a single time, without a contention net. Gill nets (with mesh sizes varying from 3 to 9 cm between adjacent knots) were used in high order sites (sites 3, 4 and 5). The sequence of gill nets was determined at random and remained submerged from the end of the day until the next morning. The fishes were put into plastic bags, and afterwards they were fixed in formalin 10% for 2 days and later transferred to alcohol 70% until the accomplishment of the analysis.
In the laboratory, the fishes were identified until species level. To evaluate the degree of stomach repletion (filling state of the stomach), the following values were attributed: 1 for empty stomachs, 2 for partially full stomachs and 3 for stomachs completely full (Braga, 1990). The stomachs with repletion degree equivalent to 3 were extracted from the visceral cavity for a subsequent study of species diet.
The grade of feeding preference (GFP) method (Braga, 1999) was applied to stomachs considered replete. This method consists of attributing values to food items found in certain species, according to the participation of each item in the analysed stomach. The value 4 is attributed when only one food item exists in the stomach; when there appears to be more than one item for the stomach, the adopted values are 3, 2 or 1, according to its participation in the analysed stomach content. The value of GFP is obtained through the sum of the values attributed to each item, divided by the number of analysed stomachs. Thus, food items can be classified as absolute preferential (GFP = 4), preferential in high degree (3 < GFP < 4), preferential (2 < GFP < 3), secondary (1 < GFP < 2) and occasional (0 < GFP < 1).
The diet width of the species was verified by applying Pielou's evenness index (J ') with a method of re-sampling (bootstrap), that allows comparisons between species through overlaps of the values of confidence intervals. The niche overlap among species was obtained by Pianka's overlap index (Oi,j). This index ranges between 0 (no overlap) to 1 (complete overlap). According to Jaksic and Marti (1981), a low overlap occurs with values between 0 and 0.33, an intermediate overlap between 0.34 and 0.66 and a high overlap between 0.67 and 1.0. For species with high overlap, we tested the existence of significant differences in exploitation of food resources by using the technique of null models with the RA3 randomisation algorithm, which maintains the niche breadth and randomises the structural zeros, as proposed by Gotelli and Entsminger (2004).
The GFP method was used to analyse the food items by season and sample sites. The Spearman test (Siegel, 1975) was applied to verify if there was a correlation between the items ingested by the species and the sampling time (drought and rainy seasons). The Friedman test was used to verify if there existed any correlation between ingested items and sampling sites. To analyse the relationships between species/food items and sites/food items, a correspondence analysis (CA) was applied on two contingency tables (Greenacre, 1984 apud Chabanet et al., 1997), one for the GFP values of food items for each species, and one for the GFP values of food items for each sampling sites.
3. Results
We analysed 576 full stomachs of 28 species (Table 1). The food items found in the stomach contents were immature Diptera (Chironomidae, Simulidae and Chaoboridae), immature Trichoptera, immature insects (not Diptera), immature Plecoptera, immature Ephemeroptera, immature Odonata, terrestrial insects (Coleoptera, Hymenoptera, Diptera, Homoptera), insect remains (locomotor appendages, wings, antennae, elytra), arachnids (spiders and water mites), annelids (Oligochaeta), worms (Nematoda), crustaceans, plant, algae, seeds, detritus and fish.
Grouping these food items into broad ecological categories, we observed that the autochthonous resources were present in most diets of the species studied, corresponding to 58%, while the allochthonous resources accounted for 30%.
The list of food items present in stomach contents of species that had few stomachs analysed is shown in Table 2. Immature insects (with a group of Diptera, Trichoptera and not Diptera) were found in the stomachs of 7 of the 11 species considered, insect remains were present in the stomachs of 5 species, terrestrial insects were common food items to the diet of 4 species and plant in 3. Eigenmannia virescens, Pimelodella sp.1, Prochilodus lineatus and Parodon nasus showed in their stomachs immature Odonata, Nematoda, algae and detritus, respectively.
The values of GFP for the species that had more than five stomachs examined are presented in Table 3. The higher levels of GFP belonged to the following items: immature Diptera (preferencial item for Trichomycterus sp., and Imparfinis mirini and secondary to Cetopsorhamdia iheringi), fish (preferencial item for Rhamdia quelen), detritus (preferencial item for Apareiodon ibitiensis and A. affinis) and insect remains (preferencial item for Odontostilbe microcephala, Astyanax bockmani and Bryconamericus sp. and a secondary item for B. stramineus, A. altiparanae, Piabina argentea, A. fasciatus and A. scabripinnis paranae).
The correspondence analysis (Figure 2) showed the formation of three groups: the first with the two species of Apareiodon, which feeds mainly on detritus, the latter formed only by Rhamdia quelen, which feeds mainly on fish, and the third group formed by other species that consumed mainly insects (immature Diptera and insect remains).
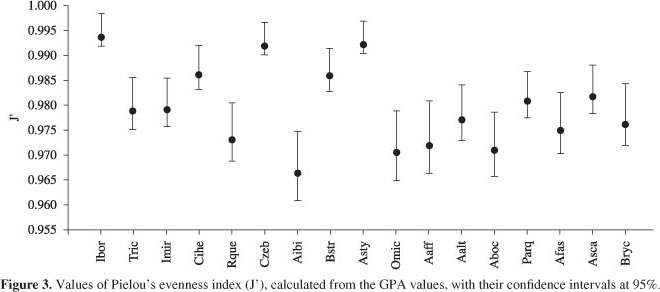
The highest values of Pielou's evenness index were presented for Imparfinis borodini, Characidium zebra and Astyanax sp.: these species consumed a wide variety of food items, being all these items considered occasional. In contrast, the lowest values of this index were found for Rhamdia quelen, Apareiodon ibitiensis, Odontostilbe microcephala, Apareiodon affinis and Astyanax bockmani that consumed less variety of items, showing preference for one food item (Figure 3).
As the confidence intervals for Imparfinis borodini, Characidium zebra and Astyanax sp. were overlapping, the diets of these species showed no significant differences. These three species also had similar diets to Cetopsorhamdia iheringi and Bryconamericus stramineus, which also ate a wide variety of items, but one of these items presented secondary importance. Cetopsorhamdia iheringi and B. stramineus, in turn, had similar diets to the other species, which had a preferential item or secondary items in their diets (Figure 3).
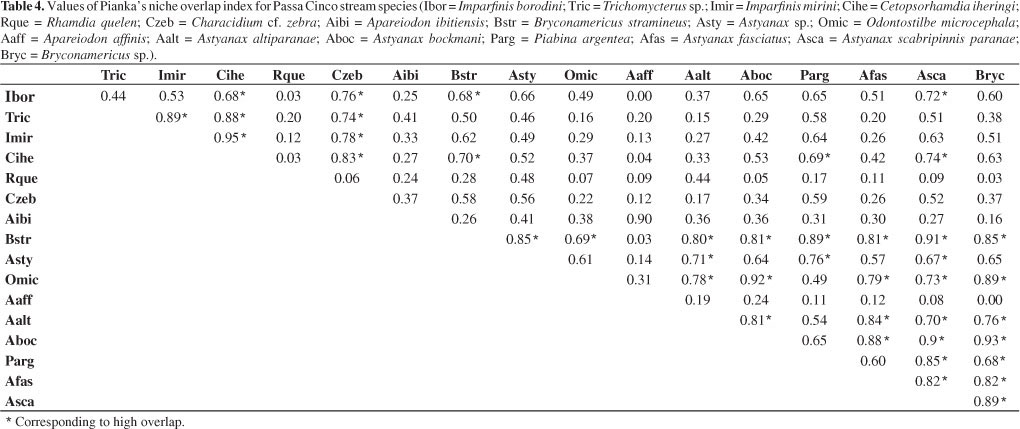
Of the total combination pairs of species, 29.4% showed high overlap (Table 4), which occurred mainly within species that consumed aquatic insect larvae. However, those species showed significant differences in the exploitation of food resources (p < 0.0001). Rhamdia quelen did not show high overlap with any other species, because only their diet includes fish.
The values of GFP for each item found in stomach contents examined in each sampling time are shown in Table 5. The items with higher values of GFP were immature Diptera for the dry season and insect parts for the rainy season. The Spearman's test did not reveal significant difference between the items consumed by the species at different times (r = 0.836, p < 0.05).
The values of GFP for the food items found in stomach contents examined in each sample point are shown in Table 6. In site 1 the item with the highest value of GFP was immature Diptera, while in other sites, insect remains was the item that showed the highest values. The results of the Friedman's test indicated that the food items found in stomach contents of species sampled in different sites were not different (χ2 = 2.36, p > 0.05).
Through the CA (Figure 4) it is possible to observe the presence of an environmental gradient, going from site 1 (headwater area) toward point 5 (mouth), which is confirmed by the physiographic characteristics of sampling sites (Table 7). Plant fragments, seeds and worms were the food items that were associated with the higher-order site (site 5). The other food items consumed by the species were found associated with other points (sites 1, 2, 3 and 4).
4. Discussion
The rivers with native riparian vegetation provide a wide variety of food items, mainly insects and plants, which have great importance in food webs and survival of tropical freshwater fish (Lowe-McConnell, 1999). Alvim and Peret (2004) concluded that both autochthonous items as well as those of allochthonous origin are dependent on riparian forest, hence the importance of conserving the forest to maintain the ecological balance of a river. In the Passa Cinco stream, the analysis of the diets showed a preponderance of autochthonous items for the different species of fish. The autochthonous insects were very important, but the allochthonous items are the base of the food web in low order streams, feeding the prey of most native fish (Castro, 1999). Braga and Gomiero (2009) analysed the feeding of the fish fauna in oligotrophic streams of the eastern Serra da Mantiqueira and found that most species used food sources of autochthonous origin, based mainly on larvae and insect nymphs.
The presentation of food items of uncommon or rare species is of great importance for understanding the biology of these species, even if they are beyond the qualitative and quantitative analysis. The species which had very few stomachs examined showed the same trend, more frequently containing autochthonous food items, except for Cetopsis gobioides which ingested terrestrial insects.
The immature Diptera and other insects were the main food items ingested by fish of the Passa Cinco stream, being a common feature in the streams of the Neotropics (Uieda et al., 1997; Casatti and Castro, 1998; Mazzoni and Lobón-Cervia, 2000). However, most of the items were occasional, showing alternative habits of these species. Omnivory is widespread among stream fishes (Abelha et al., 2001). The omnivorous diet, however, is only possible if the species has morphological and physiological adaptations in order to make use of the food item (Gerking, 1994). The species that directs the feeding habits to certain autochthonous food items such as larvae and nymphs of insects and periphyton, for example, need to have a mouth with specialised adaptations to seize these items (Braga, 2005; Braga et al., 2008, 2009). Cheirodontinae and Tetragonopterinae have visible changes in the mouth in relation to dentition (Britski, 1970), which makes these groups report to an omnivore diet. Therefore, omnivory is not opportunist, which denotes a lack of preference; omnivory is a wide possibility in the intake of items available. The plasticity of species enables the use of a wide variety of food resources in adverse conditions, where less tolerant species would not be able to establish themselves (Teixeira et al., 2005).
The great diversity and variation in community composition of aquatic invertebrates in streams throughout the year may be the explanation for the extensive overlap in the diet of fish (Angermeier and Karr, 1983). The different micro-habitats, periods of activity and tactics used to catch food can minimise the effects of overlap (Casatti, 2002, Esteves et al., 2008). Sabino and Castro (1990) found several cases of high food overlap in the diets of different species and found that they had differences in spatial distribution and feeding periods. Similar results were found by Casatti and Castro (1998) who observed that the foraging activities of two Characidium species occurred during daytime, whereas in Trichomycterus sp., they were predominantly nocturnal. The diets of Apareiodon affinis and A. ibitiensis are very close because of the large taxonomic similarities of the two species, and also the oral apparatus and feeding behaviour, as found for the species of the wetland Curimata (Sazima and Caramaschi, 1989). However, the times of the day which species forage, the preferred sites for feeding and the occurrence of these species can reduce a possible overlap and food competition (Wootton, 1992). On the other hand, if the food source is plentiful there is no competition (Aranha et al., 1993). The catfish, Rhamdia quelen, presented omnivorous habit with a predominance of insects in rivers of Serra do Mar (Gomiero et al., 2007). However, Gomes et al. (2000) characterised this species as omnivore-piscivores and Castro and Casatti (1997) showed that this fish was predominantly insectivorous. These variations occur as a result of ontogenetic changes in the diet of this species, insects are preferencial items for fish in smaller length classes and fish for those of the largest length classes (Gomiero and Braga, 2005).
The presence of a gradient, going from site 1 (headwater area) toward point 5 (mouth), reflected the environmental characteristics present in each of the sampling sites, whereas the bottom type, marginal vegetation, degree of shading, current type and mean width founded for each site was particular as shown by Rondineli and Braga (2009). In the lower orders of stream, the primary production is low due to shading from the riparian forest (Rezende and Mazzoni, 2005). The entire food web is directly or indirectly subject to the contribution of allochthonous material carried into its bed, which is used by organisms that feed at these items, primarily immature insects (Melo et al., 2004), which will be consumed by several species of fish with diurnal and nocturnal habits, benthic or nectonic. With increasing width of the river (higher orders) or lack of riparian forest, the sunlight is more concentrated, increasing the primary production and also increasing the number of fish that consume algae and periphyton (Power, 1984).
Most tropical waters show seasonal variations caused mainly by hydrometric fluctuations (Lowe-McConnell, 1999) that alter water levels and affect the habitat and thus food availability (Aranha et al., 2000). This way, it is expected that during its life cycle, the Neotropical stream fish encounter temporal and spatial variations which are eventually reflected in their diet (Power, 1983). According to Esteves and Aranha (1999), depending on rainfall patterns and local features, seasonal variations cannot be detected in the diet of the species. In the Passa Cinco stream, no significant difference was found when the food items were analysed in spatial and temporal dimensions. Thus, it is believed that in spite of possible seasonal variation in the habitat structure, the food resources should be available throughout the year and along the extension of the stream, as observed by Esteves and Lobón-Cerviá (2001) for Atlantic Rainforest streams. Braga (2006) states that the food resources, while limited in some streams, remain constant throughout the year, since the system remains in equilibrium with the surroundings.
The determination of the importance of allochthonous or autochthonous items in the diet of a fish community is difficult due to extensive interactions and dietary differences of these species, leading to a consumption of a variety of items according to the availability in the environment, which makes the plasticity very common (Gomiero and Braga, 2005).
Acknowledgements - The authors would like to thank FAPESP for the Master's scholarship conceded to first author (proc. n° 04/12669-3), IBAMA (proc. n° 02027.000234/2005-05) for authorising the license for the execution of field works, the designer Jaime R. Somera for map confection and the two anonymous reviewers for suggestions proposed.
Received October 15, 2009
Accepted June 29, 2010
Distributed February 28, 2011
- ABELHA, MCF., AGOSTINHO, AA. and GOULART, E., 2001. Plasticidade trófica em peixes de água doce. Acta Scientiarum, vol. 23, no. 2, p. 425-434.
- ALVIM, MCC. and PERET, AC., 2004. Food resources sustaining the fish fauna in a section of the upper São Francisco River in Três Marias, MG, Brazil. Brazilian Journal of Biology = Revista Brasileira de Biologia, vol. 64, no. 2, p. 195-202.
- ANGERMEIER, PL. and KARR, JR., 1983. Fish communities along environmental gradients in a system of tropical streams. Environmental Biology of Fishes, vol. 9, no. 2, p. 117-135.
- ARANHA, JMR., CARAMASCHI, EP. and CARAMASCHI, U., 1993. Ocupação espacial, alimentação e época reprodutiva de duas espécies de Corydoras Lacépède (Siluroidei, Callichthyidae) coexistentes no rio Alambari (Botucatu, São Paulo). Revista Brasileira de Zoologia, vol. 10, no. 3, p. 453-466.
- ARANHA, JMR., GOMES, JHC. and FOGAÇA, FNO., 2000. Feeding of two sympatric species of Characidium, C. lanei and C. pterostictum (Characidiinae) in a coastal stream of Atlantic Forest (Southern Brazil). Brazilian Archieves of Biology and Technology, vol. 43, no. 5, p. 527-532.
- ARAÚJO, FG., FICHBERG, I., PINTO, BCT. and PEIXOTO, MG., 2001. Variações espaciais na assembléia de peixes do rio Paraíba do Sul (Barra Mansa, Barra do Piraí), Rio de Janeiro, Brasil. Revista Brasileira de Zoologia, vol. 18, no. 2, p. 483-492.
- BRAGA, FMS., 1990. Aspectos da reprodução e alimentação de peixes comuns em um trecho do rio Tocantins entre Imperatriz e Estreito, Estados do Maranhão e Tocantins, Brasil. Revista Brasileira de Biologia = Brazilian Journal of Biology, vol. 50, no. 3, p. 547-558.
- -, 1999. O grau de preferência alimentar: um método qualitativo e quantitativo para o estudo do conteúdo estomacal de peixes. Acta Scientiarum, vol. 21, no. 2, p. 291-295.
- -, 2004. Hábitat, distribuição e aspectos adaptativos de peixes da microbacia do Ribeirão Grande, estado de São Paulo, Brasil. Acta Scientiarum, vol. 26, no. 1, p. 31-36.
- -, 2006. Aspectos da reprodução no gênero Characidium Reinhardt, 1867 (Crenuchidae, Characidiinae) na microbacia do Ribeirão Grande, sudeste do Brasil. Acta Scientiarum, vol. 28, no. 4, p. 365-371.
- BRAGA, FMS. and ANDRADE, PM., 2005. Distribuição de peixes na microbacia do Ribeirão Grande, serra da Mantiqueira oriental, São Paulo, Brasil. Iheringia Série Zoologia, vol. 95, no. 2, p. 121-126.
- BRAGA, FMS. and GOMIERO, LM., 2009. Alimentação de peixes na microbacia do Ribeirão Grande, serra da Mantiqueira oriental, SP. Biota Neotropica, vol. 9, no. 3, p. 1-6.
- BRAGA, FMS., GOMIERO, LM. and SOUZA, UP., 2008. Aspectos da reprodução e alimentação de Neoplecostomus microps, (Loricariidae, Neoplecostominae) na microbacia do Ribeirão Grande, serra da Mantiqueira oriental (SP). Acta Scientiarum, vol. 30, no. 4, p. 455-463.
- -, 2009. Biologia populacional de Pareiorhina rudolphi (Loricariidae, Hypostominae) na microbacia do Ribeirão Grande, serra da Mantiqueira oriental, Estado de São Paulo. Acta Scientiarum, vol. 31, no. 1, p. 79-88.
- BRITSKI, HA., 1970. Peixes de água doce do Estado de São Paulo. In: Comissão Interestadual da bacia Paraná-Uruguai, Poluição e Piscicultura. São Paulo: Faculdade de Saúde Pública USP, Instituto de Pesca (CPRN). p. 79-108
- CASATTI, L. 2002. Alimentação dos peixes em um riacho do Parque Estadual Morro do Diabo, Bacia do Alto Rio Paraná, sudeste do Brasil. Biota Neotropica, vol. 2, no. 2, p. 1-14.
- CASATTI, L. and CASTRO, RMC., 1998. A fish community of the São Francisco Rivers headwaters riffles, southeastern Brazil. Ichthyological Exploration Freshwaters, vol. 9, no. 3, p. 229-242.
- CASTRO, RMC., 1999. Evolução da ictiofauna de riachos sul-americanos: padrões gerais e possíveis processos causais. In: CARAMASCHI, EP., MAZZONI, R. and PERES-NETO, PR., Ed. Ecologia de peixes de riachos. Rio de Janeiro: PPGE-UFRJ. p. 139-155. Série Oecologia Brasiliensis, vol. 6.
- CASTRO, RMC. and CASATTI, L., 1997. The fish fauna from a small forest stream of the upper Paraná River basin, southeastern Brazil. Ichthyological Exploration of Freshwaters, vol. 7, no. 4, p. 337-352.
- CASTRO, RMC., CASATTI, L., SANTOS, HF., MELO, ALA., MARTINS, LSF., FERREIRA, KM., GIBRAN, FZ., BENINE, RC., CARVALHO, M., RIBEIRO, AC., ABREU, TX., BOCKMANN, FA., PELIÇÃO, GZ., STOPLIGIA, R. and LANGEANI, F., 2004. Estrutura e composição da ictiofauna de riachos da bacia do rio Grande no Estado de São Paulo, sudeste do Brasil. Biota Neotropica, vol. 4, no. 1, p. 1-39.
- CHABANET, P., RALAMBONDRAINY, H., AMANIEU, M., FAURE, G. and GALZIN, R., 1997. Relationships between coral reef substrata and fish. Coral Reefs, vol. 16, p. 93-102.
- ESTEVES, KE. and ARANHA, JMR., 1999. Ecologia trófica de peixes de riachos. In: CARAMASCHI, EP., MAZZONI, R. and PERES-NETO, PR., Ed. Ecologia de peixes de riachos. Rio de Janeiro: PPGE-UFRJ. p. 157-182. Série Oecologia Brasiliensis, v. 6.
- ESTEVES, KE. and LOBÓN-CERVIÁ, J., 2001. Composition and trophic structure of a fish community of a clear water Atlantic rainforest stream in southeastern Brazil. Environmental Biology of Fishes, vol. 62, p. 429-440.
- ESTEVES, KE., LOBO, AVP. and FARIA, MDR., 2008. Trophic structure of a fish community along environmental gradientes of a subtropical river (Paraitinga River, Upper Tietê River Basin, Brasil). Hydrobiologia, vol. 598, p. 373-387.
- GARCIA, GJ., HEBERT, HD., PERINOTTO, JAJ. and CASTRO, JFM., Ed. 2004. Atlas ambiental da Bacia do Rio Corumbataí. 2004. Available from: <http://www.rc.unesp.br>
- GERKING, SD., 1994. Feeding ecology of fish. San Diego: Academic Press. 416 p.
- GOMES, LC., GOLOMBIESKI, JI., GOMES, ARC. and BALDISSEROTTO, B., 2000. Biologia do jundiá Rhamdia quelen (Teleostei, Pimelodidae). Ciência Rural, vol. 30, no. 1, p. 179-185.
- GOMIERO, LM. and BRAGA, FMS., 2005. Uso do grau de preferência alimentar para a caracterização da alimentação de peixes na APA de São Pedro e Analândia. Acta Scientiarum, vol. 27, no. 3, p. 265-270.
- GOMIERO, LM., MANZATTO, AG. and BRAGA, FMS., 2008. The role of riverine forests for food supply for the omnivorous fish Brycon opalinus Cuvier, 1819 (Characidae) in the Serra do Mar, southeast Brazil. Brazilian Journal of Biology = Revista Brasileira de Biologia, vol. 68, no. 2, p. 321-328.
- GOMIERO, LM., SOUZA, UP. and BRAGA, FMS., 2007. Reprodução e alimentação de Rhamdia quelen (Quoy & Gaimard, 1824) em rios do Núcleo Santa Virgínia, Parque Estadual da Serra do Mar, São Paulo, SP. Biota Neotropica, vol. 7, no. 3, p. 1-7.
- GORDON, ND., McMAHON, TA. and FINLAYSON, BL., 1995. Stream hidrology: an introduction for ecologists. Chichester: John Wiley & Sons. 526 p.
- GOTELLI, NJ. and ENTSMINGER, GL., 2004. EcoSim: null models software for ecology. Version 7. Jericho, VT: Acquired Intelligence Inc. & Kesey-Bear. 05465. Available from: <http://garyentsminger.com/ecosim/index.htm>. Access in: 05 out. 2009.
- JAKSIC, FM. and MARTI, CD., 1981. Trophic ecology of Athene owls in mediterranean-type ecosystems: a comparative analysis. Canadian Journal of Zoology, vol. 59, p. 2331-2340.
- LOWE-MCCONNELL, RH., 1999. Estudos ecológicos de comunidades de peixes tropicais. São Paulo: Editora da Universidade de São Paulo. 535 p.
- MAZZONI, R. and LOBÓN-CERVIÁ, J., 2000. Longitudinal structure, density and production rates of a neotropical stream fish assemblage: the river Ubatiba in the Serra do Mar, southeast Brazil. Ecography, vol. 23, no. 5, p. 588-602.
- MELO, CE., MACHADO, FA. and PINTO-SILVA, V., 2004. Feeding habits of fish from a stream in the savanna of Central Brazil, Araguaia basin. Neotropical Ichthyology, vol. 2, no. 1, p. 37-44.
- POWER, ME., 1983. Grazing responses of tropical freshwater fishes to different scales of variation in their food. Environmental Biology of Fishes, vol. 9, no. 2, p. 103-115.
- -, 1984. Habitat quality and the distribution of algaegrazing catfish in a Panamanian stream. Journal Animal Ecology, vol. 53, no. 2, p. 357-374.
- REZENDE, CF. and MAZZONI, R., 2005. Seasonal variation in the input of allochthonous matter in an Atlantic Rain Forest stream, Ilha Grande-RJ. Acta Limnologica Brasiliensis, vol. 17, no. 2, p. 167-175.
- RONDINELI, GR. and BRAGA, FMS., 2009. Biologia populacional de Corydoras flaveolus (Siluriformes, Callichthyidae) no rio Passa Cinco, sub-bacia do rio Corumbataí, São Paulo, Brasil. Biota Neotropica, vol. 9, no. 4, p. 1-9.
- SABINO, J. and CASTRO, RMC., 1990. Alimentação, período de atividade e distribuição espacial dos peixes de um riacho da Floresta Atlântica (Sudeste do Brasil). Revista Brasileira de Biologia = Brazilian Journal of Biology, vol. 50, no. 1, p. 23-36.
- SAZIMA, I. and CARAMASCHI, EP., 1989. Comportamento alimentar de duas espécies de Curimata, sintópicas no pantanal de Mato Grosso (Osteichthyes, Characiformes). Brazilian Journal of Biology = Revista Brasileira de Biologia, vol. 49, no. 2, p. 325-333.
- SIEGEL, S., 1975. Estatística não-paramétrica Para as ciências do comportamento. Rio de Janeiro: McGraw-Hill do Brasil. 350 p.
- TEIXEIRA, TP., PINTO, BCT., TERRA, BF., ESTILIANO, EO., GRACIA, D. and ARAÚJO, FG., 2005. Diversidade das assembléias de peixes nas quatro unidades geográficas do rio Paraíba do Sul. Iheringia Série Zoologia, vol. 95, no. 4, p. 347-357.
- UIEDA, VS. and MOTTA, RL., 2007. Trophic organization and food web structure of southeastern Brazilian streams: a review. Acta Limnologica Brasiliensis, vol. 19, no. 1, p. 15-30.
- UIEDA, VS., BUZZATO, P. and KIKUCHI, RM., 1997. Partilha de recursos alimentares em peixes em um riacho de serra do Sudeste do Brasil. Anais Academia Brasileira de Ciências, vol. 69, no. 2, p. 243-252.
- VALENTE, ROA. and VETTORAZZI, CA., 2002. Análise da estrutura da paisagem na bacia do rio Corumbataí, SP. Scientia Florestalis, vol. 62, p. 114-129.
- VANNOTE, RL., MINSHALL, GW., CUMMINS, KW., SEDELL, JR. and CUSHING, CE., 1980. The River Continuum Concept. Canadian Journal of Fisheries and Aquatic Sciences, vol. 37, no. 1, p. 130-137.
- WOOTTON, RJ., 1992. Fish Ecology. New York: Ed. Chapman & Hall, 212 p.
Publication Dates
-
Publication in this collection
11 Mar 2011 -
Date of issue
Feb 2011
History
-
Received
15 Oct 2009 -
Accepted
29 June 2010