Abstracts
Comolia ovalifolia DC Triana (Melastomataceae) and Chamaecrista ramosa (Vog.) H.S. Irwin and Barneby var. ramosa (Leguminosae - Caesalpinioideae) are tropical plant species found in restinga (herbaceous-shrubby, sandy costal ecosystems). They have flowers with poricidal anthers and are pollinated by bees. The study sought to analyse potential pollinators of both plants during visits to their flowers in a restinga area in Bahia. The flowering displayed by both species was considered continuous and long duration, constantly providing pollen to floral visitors. C. ovalifolia was visited by 17 species of bees and C. ramosa by 16 species, predominantly from the Apidae family (with a similarity index of 74%). The behavior displayed by these visiting bees was of vibrating anthers. The small-sized Euglossa sp. Latreille, 1802 and Florilegus similis Urban, 1970 bees played less of a role as pollinators, since they rarely touched the flower stigma during harvests and were thus considered opportunist visitors or casual pollinators. Centris decolorata Lepetier, 1841 (= C. leprieuri) and Xylocopa subcyanea Perez, 1901 are large bees and were considered efficient pollinators of C. ovalifolia and C. ramosa because of the higher frequency and constancy of their visits, and their favourable behaviour and size for pollen transfer between flowers, which guarantees the survival of these native restinga plant species.
buzz pollination; Apidae; Centris decolorata; Xylocopa subcyanea
Comolia ovalifolia (Melastomataceae) e Chamaecrista ramosa (Leguminosae - Caesalpinioideae) são espécies de plantas tropicais que ocorrem na restinga. Estas plantas apresentam flores com anteras poricidas e são polinizadas por abelhas. Este estudo teve como objetivo analisar os polinizadores potenciais de ambas as plantas durante as visitas nas flores em uma área de restinga da Bahia. O florescimento apresentado por ambas as espécies foi considerado contínuo e de longa duração, oferecendo sempre pólen para os visitantes florais. C. ovalifolia foi visitada por 17 espécies de abelhas e C. ramosa por 16 espécies, com predominância da família Apidae (índice de similaridade de 74%). As abelhas visitantes destas flores apresentavam o comportamento de vibrar as anteras. O papel das abelhas de pequeno porte como Euglossa sp. e Florilegus similis, como polinizadores, era pequeno, pois raramente tocavam o estigma das flores durante as coletas, sendo consideradas visitantes oportunistas ou polinizadores casuais. Centris decolorata e Xylocopa subcyanea, são abelhas grandes, e foram consideradas polinizadores eficientes de C. ovalifolia e C. ramosa, por apresentarem elevada frequência e constância de visitas nestas plantas, e comportamento e porte favoráveis à transferência do pólen entre as flores, o que garante a sobrevivência destas espécies de plantas nativas da restinga.
polinização por vibração; Apidae; Centris decolorata; Xylocopa subcyanea
ECOLOGY
Potential pollinators of Comolia ovalifolia DC Triana (Melastomataceae) and Chamaecrista ramosa (Vog.) H.S. Irwin and Barneby var. ramosa (Leguminosae-Caesalpinioideae), in restinga, Bahia, Brazil
Polinizadores potenciais de Comolia ovalifolia DC Triana (Melastomataceae) e Chamaecrista ramosa (Vog.) H.S. Irwin e Barneby var. ramosa (Leguminosae-Caesalpinioideae), na restinga, Bahia, Brasil
Oliveira-Rebouças, P.; Gimenes, M.* * e-mail: mgimenes@uefs.br
Departamento de Ciências Biológicas, Universidade Estadual de Feira de Santana UEFS, Campus Universitário, Rod. BR 116, Km 3, CEP 44031-460, Feira de Santana, BA, Brazil
ABSTRACT
Comolia ovalifolia DC Triana (Melastomataceae) and Chamaecrista ramosa (Vog.) H.S. Irwin and Barneby var. ramosa (Leguminosae - Caesalpinioideae) are tropical plant species found in restinga (herbaceous-shrubby, sandy costal ecosystems). They have flowers with poricidal anthers and are pollinated by bees. The study sought to analyse potential pollinators of both plants during visits to their flowers in a restinga area in Bahia. The flowering displayed by both species was considered continuous and long duration, constantly providing pollen to floral visitors. C. ovalifolia was visited by 17 species of bees and C. ramosa by 16 species, predominantly from the Apidae family (with a similarity index of 74%). The behavior displayed by these visiting bees was of vibrating anthers. The small-sized Euglossa sp. Latreille, 1802 and Florilegus similis Urban, 1970 bees played less of a role as pollinators, since they rarely touched the flower stigma during harvests and were thus considered opportunist visitors or casual pollinators. Centris decolorata Lepetier, 1841 (= C. leprieuri) and Xylocopa subcyanea Perez, 1901 are large bees and were considered efficient pollinators of C. ovalifolia and C. ramosa because of the higher frequency and constancy of their visits, and their favourable behaviour and size for pollen transfer between flowers, which guarantees the survival of these native restinga plant species.
Keywords: buzz pollination, Apidae, Centris decolorata, Xylocopa subcyanea.
RESUMO
Comolia ovalifolia (Melastomataceae) e Chamaecrista ramosa (Leguminosae - Caesalpinioideae) são espécies de plantas tropicais que ocorrem na restinga. Estas plantas apresentam flores com anteras poricidas e são polinizadas por abelhas. Este estudo teve como objetivo analisar os polinizadores potenciais de ambas as plantas durante as visitas nas flores em uma área de restinga da Bahia. O florescimento apresentado por ambas as espécies foi considerado contínuo e de longa duração, oferecendo sempre pólen para os visitantes florais. C. ovalifolia foi visitada por 17 espécies de abelhas e C. ramosa por 16 espécies, com predominância da família Apidae (índice de similaridade de 74%). As abelhas visitantes destas flores apresentavam o comportamento de vibrar as anteras. O papel das abelhas de pequeno porte como Euglossa sp. e Florilegus similis, como polinizadores, era pequeno, pois raramente tocavam o estigma das flores durante as coletas, sendo consideradas visitantes oportunistas ou polinizadores casuais. Centris decolorata e Xylocopa subcyanea, são abelhas grandes, e foram consideradas polinizadores eficientes de C. ovalifolia e C. ramosa, por apresentarem elevada frequência e constância de visitas nestas plantas, e comportamento e porte favoráveis à transferência do pólen entre as flores, o que garante a sobrevivência destas espécies de plantas nativas da restinga.
Palavras-chave: polinização por vibração, Apidae, Centris decolorata, Xylocopa subcyanea.
1. Introduction
The coastal sand plains of Brazil are characterised by herbaceous-shrubby vegetation known as restinga. In this habitat the soil is sandy, acidic and poor, because of the heavy rainfall that washes away nutrients. The fauna and flora that occur in the restinga habitat are adapted to its dominant physical factors such as high salinity, high temperature, winds and salt coming from the ocean, dry sand sediments (due to quick drainage) and intense sunlight exposure (PRODESU, 2001). Human activity along the Brazilian coastline is threatening this habitat and the Brazilian Environment Agency considers restinga a highly fragile ecosystem (Lacerda and Esteves, 2000). Much of this habitat may be lost if it is not protected (Rocha et al., 2007).
There is still very little information about the ecological process that occurs in the restinga habitat, but some relevant information exists regarding interactions between the plants and pollinators that are found in this environment. According to Gottsberger et al. (1988), there is a predominance of melittophily in plant pollination systems in dunes and restinga. Some studies carried out in the restinga of Northeastern Brazil have confirmed that bees are important pollinators of plants (Gottsberger et al., 1988; Silva and Martins, 1999; Costa and Ramalho, 2001; Ramalho and Silva, 2002; Oliveira-Rebouças and Gimenes, 2004; Viana and Kleinert, 2005, 2006; Oliveira et al., 2010).
Certain bee genera, such as Xylocopa and Centris, are very important in the pollination of restinga native plants, mainly because of their larger size which facilitates pollen harvesting in environments where strong winds prevail (Gottsberger et al., 1988; Oliveira-Rebouças and Gimenes, 2004; Viana and Kleinert, 2006; Oliveira et al., 2010). These bees are capable of harvesting pollen from flowers with poricidal anthers through vibrations of their flight musculature (buzz pollination) (Buchmann, 1983). Species of Leguminosae (Caesalpinioideae) and Melastomataceae with poricidal anthers are well represented in an environment of dunes and restinga (Pinto et al., 1984; Queiróz, 2007). Gimenes et al.(2007) studied the interactions between bees and seven plants that are very common in the restinga in Bahia and observed that Comolia ovalifolia DC Triana (Melastomataceae) and Chamaecrista ramosa (Vog.) H.S. Irwin and Barneby var. ramosa (Leguminosae- Caesalpinioideae) showed a high degree of similarity in visiting bees and both have poricidal anthers.
Comolia is a widespread Melastomataceae in tropical South America (Renner, 1993) and C. ovalifolia is an endemic species of the restinga along the Northern coast of Bahia (Britto and Noblick, 1984). Chamaecrista (Caesalpinioideae) has an extensive distribution in Brazil, and occurs in a variety of habitats such as rocky habitats, dry forests (caatingas), tropical savannas (cerrados), open fields, wet areas and sandy dunes, and is quite common in restinga habitats (Pinheiro et al., 1988). C.ramosa is widespread and common in the restinga of northeastern Brazil (Viana and Kleinert, 2006; Silva et al., 2008).
Some authors have suggested that co-occurring plants with poricidal anthers, regardless of genera or family, share their visitors or pollinators (Linsley and Cazier, 1963 apud Thorp and Estes, 1975). Solanum rostratum Dunal and Chamaecrista fasciculata (Michx.) Greene (= Cassia fasciculata), exhibit many differences between their flowers, but have poricidal anthers, and shared large bees, which collected pollen early in the morning (Thorp and Estes, 1975), as their common visitors.
This research aimed to study the sharing, in different months, of visiting bees as potential pollinators of two plant species with poricidal anthers - Comolia ovalifolia and Chamaecrista ramosa - that are very common in the dunes and restinga on the Northern coast of Bahia.
2. Material and Methods
This study took place at the Mingu ranch, a restinga area located in the Environmental Protection Area of the Capivara River, Arembepe, Bahia, Brazil (12º 43' 42" S and 38º 08' 49" W). Data were collected from January 2001 to February 2002 and from May to November 2002. The climate in the area is tropical hot and wet (AM, according to Köppen's classification). The average temperature and relative humidity ranged throughout the study from 21 to 28 °C and 76.7 to 91%, respectively. The annual rainfall in 2001 was 1.528 mm, while in 2002 it was 1.168 mm, and the rainy season occurs between March and October (Figure 1 - Data obtained from CETREL S.A, Environment Protection Company). Microclimatic data (temperature, relative humidity, and light intensity) were collected during field observations using a thermohygrometer and a luximeter.
The study area was about 5 ha, and the vegetation at the Mingu ranch is well preserved, with essentially two types of vegetation; an herbaceous-shrubby layer dominating the area and an arboreal stratum, with sparse trees. The herbaceous-shrubby layer is composed mainly of species of Melastomataceae, Scrophulariaceae, Krameriaceae, Leguminosae, and Lythraceae, while the arboreal stratum has mainly Vochysiaceae and Malpighiaceae (Gimenes et al., 2007). The species studied, C. ovalifolia and C. ramosa are extremely common across the area. Both species tend to form small clusters and are an important part of the herbaceous-shrubby stratum.
The study was conducted in one area (100 m2) where clusters of C. ovalifolia and C. ramosa could be found. C. ovalifolia is a small shrub, approximately 1 m high, and C. ramosa is a small prostrate or decumbent shrub that creeps close to the ground and reaches 20 cm. All the plants were monitored at the same time. The total number of flowers in the area was recorded monthly.
The time of flower opening for both plant species was recorded. Data regarding the diameter, colour, symmetry, and size were recorded from 10 randomly chosen flowers for each species.
On the day before the flower anthesis, 13 buds were enclosed in voile bags in order to test stigma receptivity. When the flower opened, a drop of v10 hydrogen peroxide was applied to the stigma surface (Dafni et al., 2005); if receptivity was positive bubbles were formed. This test was performed on an hourly basis during the time that the flower remained open.
Observations regarding the number of bee visits were carried out on a monthly basis for 30 minutes in every hour between 5:30 AM and 6:00 PM, over two or three consecutive days. The bee visit count was based on the number of times each bee buzzed the flowers for pollen collection.
Similarity between the species of bees collecting pollen from C. ovalifolia and C. ramosa flowers was obtained by means of the Sorensen Similarity Index (Krebs, 1989).
Bee constancy was calculated using the Equation 1:
C = (Number of sampling with species i / Number of sampling) × 100 (1)
According to the values obtained, the species were classified into: C = constant (C > 50%), A = accessory (C = 25-50%), and Ac = accidental (C < 25%) (Silveira-Neto et al., 1976).
The data regarding number of visits was normalised through the square root transformation method (Zar, 2010). The patterns of distribution for the number of bee visits to the plant species studied were compared using the Kolmogorov-Smirnov test, with p < 0.05 (Magurran, 2003).
When analysing the frequency of visits by different bee species to both plants, the accidental category was not considered. Species of the genera Centris and Xylocopa that could not be identified when on the flowers were grouped as Centris spp. and Xylocopa spp. Within this Centris spp. group were C. aenea, C. caxiensis, C. trigonoides, C. analis, C. hyptidis, and C. nitens, all of which are hard to identify when on flowers. The Xylocopa spp. group included X. suspecta and X. cearensis.
To classify bee frequency, a confidence interval for frequency means was established (percentage of the number of visits of a species in relation to the total number of visits observed) with a 5% probability using the following classification: very frequent (frequency greater than the CI upper limit of 5%, greater than 23% for C. ovalifolia and greater than 16.4% for C. ramosa); frequent (frequency within CI of 5%, between 1.6 and 22.9% for C. ovalifolia, and 4.7 and 16.3% for C. ramosa), and not very frequent (frequency below the CI lower limit of 5%, less than 1.59% for C. ovalifolia and less than 4.69% for C. ramosa) (adapted from Thomazini and Thomazini, 2002).
The non parametric Spearman correlation was used to assess a possible association between climatic factors (temperature, relative humidity, or light intensity) and the number of daily visits. The statistical package PRIMER 5 for Windows (Clarke and Warwick, 2001) was used to carry out these analyses.
The collected bees are deposited in the Johan Becker entomological collection of the Museu de Zoologia da Universidade Estadual de Feira de Santana (MZUEFS). The supraspecific classification of the bees follows Michener's proposal (2007). Vouchers for the collected plant species (numbers: 50624 and 50622), are deposited in the Herbarium of the Universidade Estadual de Feira de Santana (HUEFS). The supraspecific classification of the plants follows Cronquist's proposal (1981) for Melastomataceae, and Lewis et al. (2005) and APGII (2003) for Leguminosae.
3. Results
During the study the flowering of C. ovalifolia and C. ramosa in the restinga of Arembepe was continuous. C. ovalifolia displayed a greater number of flowers in March 2001, from September to November 2001, and in October 2002, while C. ramosa flowered mainly from July to October 2001, and in August and October 2002 (Figure 2).
In the months with peak flowering temperatures varied from 23 to 27 °C (for both species) and rainfall from 16 to 224 mm for C. ovalifolia and from 34 to 186 mm for C. ramosa. Both plants had more flowers between the months of September and November 2001 and in October 2002. Rainfall varied during the months of greater flowering for both plants (Figures 1 and 2). The correlation between rainfall and flowering of both plants was not significant, but there was a significant correlation between flowering and temperature for C. ramosa (-0.607).
C. ovalifolia and C. ramosa have zygomorphic flowers and poricidal anthers, and pollen is the only available floral resource. The four purplish petals of C. ovalifolia had a diameter of 22 mm and the five yellow petals of C. ramosa had a diameter of 25 mm. The eight sickle-shaped anthers in C. ovalifolia were joined beneath the style, since the stigma was generally located between the anthers. The pentamerous flowers of C. ramosa displayed four petals that were slightly different from each other and a fifth with red blemishes at its base, generally larger than the others and shaped like a keel; together these petals surrounded the 10 poricidal dehiscence stamen. The style was positioned to the right on one flower and to the left on another, demonstrating enantiostyly.
C. ovalifolia flowers opened between 5:30 AM and 6:45 AM, and closed between 4:00 PM and 5:30 PM. The flowers of C. ramosa opened between 4:30 AM and 6:00 AM and closed between 11:00 AM and 12:00 AM. This data was consistent with the light intensity to which the species responded (from 2 to 81 lux for C. ramosa and from 24 to 272 lux for C. ovalifolia) but there was no difference in response to temperature (from 19 to 28 °C) or relative humidity (from 76 to 94%). The stigmas of both plants were receptive from 7:00 AM and remained so until the end of anthesis. As soon as the flowers of both plants were open they were visited by bees. Visits to C. ramosa flowers ceased at 12:00 AM, while the flowers of C. ovalifolia were visited until 1:00 PM.
A total of 21 species of bees were collected on C. ovalifolia and C. ramosa, of which 18 belonged to the Apidae family (Table 1). Similarity in bee species visiting the flowers was high, with a Sorensen Similarity Index of 0.74%.
The C. ovalifolia flowers were more frequently visited by bees during the months of April, October, and November 2001, as well as in October 2002, while C. ramosa were mostly visited in October 2001 and October 2002 (Figure 2). For both plants there was a drop in bees visiting the flowers from May to August 2001, and from July 2002 to August 2002, although C. ramosa had an increased number of flowers between July and August 2001 and in August 2002. There was a significant correlation between temporal abundance of flowers and visits of bees in C. ramosa (0.702) but not between flowers and visitors in C. ovalifolia (0.105).
In an analysis of the influence of abiotic factors on the daily activity of pollen-collecting bees on C. ovalifolia flowers, light intensity was the factor that showed the highest correlation coefficient (0.75). When looking at visits to C. ramosa flowers, however, the highest correlation (0.66) was with a combination of temperature, humidity and light intensity.
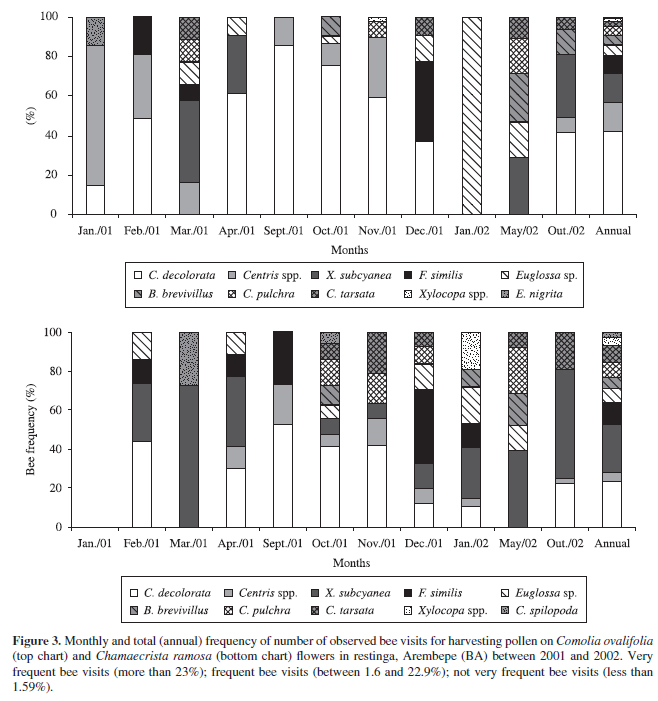
Data regarding the constancy of bees on flowers of both plants are shown on Table 1, and data about monthly or total (annual) frequency of visiting bees can be seen in Figure 3. In terms of bee species, Centris decolorata (= C. leprieuri) represented 41.8% of all visits to C. ovalifolia and 23.1% of visits to C. ramosa; this was considered a very frequent and constant species for both plants. On the other hand, Xylocopa subcyanea represented 14.8% of visits to C. ovalifolia and was considered a frequent and accessory species for this plant, while being a very frequent and constant species for C. ramosa, at 24.7% of visits.
C. decolorata was a very frequent visitor to C. ovalifolia flowers in eight of the 11 months of study, and X. subcyanea was in four of the months studied. C. decolorata were considered very frequent visitors to the C. ramosa flowers in six of the 10 months of observation, and X. subcyanea, in seven of these months. In February and April 2001 and in October 2002, both bee species were very frequent visitors to C. ramosa flowers, and in April 2001 and October 2002 very frequent visitors to C. ovalifolia flowers. During February 2001 and January 2002, X. subcyanea were very frequent visitors to the C. ramosa flowers, but were not observed on C. ovalifolia (Figure 3).
Bees from the Centris spp. group were very frequent in January, February and November 2001 on C. ovalifolia flowers and in September 2001 were very frequent on C. ramosa flowers.
Regarding the annual data, we found other species that were considered frequent visitors to both plant species: Florilegus similis (8.8% in C. ovalifolia and 11.5% in C. ramosa), Centris pulchra (4.3% in C. ovalifolia and 7.8% in C. ramosa), Centris tarsata (2.7% in C. ovalifolia and 9.0% in C. ramosa), Euglossa sp. (5.4% in C. ovalifolia and 7.0% in C. ramosa) and Bombus brevivillus (5.0% in C. ovalifolia and 5.5% in C. ramosa) (Figure 3).
All bees observed visiting C. ovalifolia and C. ramosa flowers buzzed the poricidal anthers for pollen collection. The bees released pollen when buzzing the anthers; this pollen adhered to the insect hairs and was transferred to the scopae or corbiculae (in the case of Euglossa sp.). Differences were observed in how the bees touched the anthers during pollen collection, primarily in terms of the position of the stigma of both plants. In both plants, Euglossa sp. and F. similis landed and buzzed one anther at a time and only rarely touched the flower's stigma. Bees such as C. decolorata and X. subcyanea came together and buzzed all anthers at once, resulting in both plants in the release of larger amounts of pollen, sufficient to cover the bees' bodies. Pollen from C. ovalifolia flowers was released towards the ventral portion of the body which touched the stigma. When this species visited C. ramosa flowers, the pollen was released towards both the ventral and the dorsal parts of the body. Moreover, in this species the stigma sometimes faced to the right and sometimes to the left, thus touching the dorsal or lateral portions of the bees' bodies and favoring pollen transfer.
4. Discussion
Flowering occurs throughout the year and, when we consider other studies of these species carried out in similar habitats (Viana et al., 2006), this suggests a continuous flowering for both species, constantly providing pollen to floral visitors. The greatest flowering occurs during spring and early summer, with climatic factors exerting little influence, as seen in the lack of significance in all the correlation tests except that of temperature, which may have influenced the flowering of C. ramosa.
C. ramosa showed a positive correlation between the number of flowers and bee visits. According to Gentry (1974), the large-scale production of flowers over several months, as was observed in both restinga plants, may be a strategy to ensure the attraction of large numbers of floral visitors and potential pollinators.
The flowers of C. ramosa and C. ovalifolia were visited exclusively by bees. The morphological characteristics of the flowers of these plants, that is their poricidal anthers, zygomorphic form, and yellow and lilac colours, mean that they are considered melittophilous (according to Faegri and Van Der Pijl, 1979). Since pollen is the only floral resource available to the bees in both plant species, they are classified as "pollen flowers", as stated by Vogel (1978).
According to Buchmann (1983), pollen seems to be more selective than nectar as a floral attraction. Thus flowers whose sole resource is pollen may be pollinated more efficiently by specialized bees (Vogel, 1978; Buchmann, 1983). When analysing the relationship between flowers and bees in coastal dunes with sandbank vegetation in Bahia, Ramalho and Silva (2002) observed that specialisations in flower morphology can restrict the visitor's use of floral resources. While this may appear restrictive, it may also enhance efficiency in pollination and use of flower resources.
Although the flowers of C. ramosa and C. ovalifolia have different shapes and colours, both possess poricidal anthers and share many visiting bee species, as is indicated in the high similarity index. These observations suggest that morphological characteristics such as colour and shape do not restrict the bees to one or another plant species, and in our study the pollen liberated by the poricidal anthers attracts bees according to one restriction only: ability to buzz anthers. This shows that anther characteristic is probably an important factor in attracting bees to flowers and in their consequent pollination.
All the bees observed visiting the C. ovalifolia and C. ramosa flowers in the restinga of Arembepe had the ability to buzz these plants' poricidal anthers, and the majority belonged to the Apidae family. The predominance of Apidae bees in restinga and costal dune environments has been noted in other studies undertaken in Northeastern Brazil (Gottsberger et al., 1988; Albuquerque et al., 2007; Silva and Martins, 1999; Costa and Ramalho, 2001; Viana and Kleinert 2005; Gimenes et al., 2007).
Of all the species of bees that visited the C. ovalifolia and C. ramosa flowers, ten belonged to the Centris genus and four to the Xylocopa genus; these bees are considered large and are more common than small bees in the dunes and restinga in the Northeast of Brazil (Gottsberger et al., 1998). In other studies, these bees were observed collecting pollen from C. ovalifolia (Viana and Kleinert, 2006), Comolia lythrarioides (Steudel) Naudin (Ribeiro et al., 2008), Chamaecrista hispidula (Vahl) H.S. Irwin and Barneby (Gottsberger et al., 1988) and C. ramosa (Silva and Martins, 1999; Costa and Ramalho, 2001; Viana and Kleinert, 2006).
The daily activity of bees on the flowers of C. ovalifolia was probably influenced by light intensity. For the flowers of C. ramosa, however, the combined influence of temperature, relative humidity and light intensity attracted the largest number of bee visits to these flowers. Pinheiro et al. (1988), state that adverse climatic conditions (high humidity and temperature and low light intensity) interfere with the pollen availability of C. ramosa flowers.
While many species of bees were observed visiting the flowers in this study, only a small number were considered constant and very frequent for both plants. C. decolorata and X. subcyanea, which are considered to be principally tropical bees (Silveira et al., 2002), visited both plant species frequently and more constantly than the other bees species, but X. subcyanea showed a preference for the flowers of C. ramosa. The morphology of this bee, which is black and large, may be associated with its thermoregulation, which may in turn be related to its preference for visiting a flower that opens early in the morning.
Furthermore, the size of bees and their pollen collection behaviour was adapted to the flowers' dimension and shape, facilitating the transfer of floral resources and thus making them efficient pollinators of these plants. Thorp and Estes (1975) associated the body size and vibration behaviour of bees in poricidal anthers with thermoregulation; this allowed them to be active at low temperatures. Gottsberger et al. (1988) associated the dominance of large bees in the sandy environments of dunes and restinga with the occurrence of the strong winds that interfere in the harvesting activities of smaller-sized bees.
Other studies conducted in the restinga of northeastern Brazil considered Xylocopa spp. and C. decolorata (= C. leprieuri) species as potential pollinators of the flowers of plant species that are found in this environment (Gottsberger et al., 1988; Silva and Martins, 1999; Oliveira-Rebouças and Gimenes, 2004; Ribeiro et al., 2008). C. decolorata was not recorded in the restinga of the southern states of Brazil (Paraná and Rio Grande do Sul) (Zanella et al., 1998; Alves-dos-Santos, 1999), which appears to be consistent with a geographic restriction to Brazil's warmer states.
Other bee species are considered frequent and constant visitors to C. ovalifolia and C. ramosa flowers, although they do not contribute significantly to their pollination. This is the case with Euglossa sp., which is constant and frequent, and F. similis which is frequent in both plants. These are small-sized bees and rarely touch the stigma of the flowers during pollen harvesting and are therefore considered opportunist visitors or casual pollinators (Oliveira-Rebouças and Gimenes, 2004).
Research regarding bees and the interaction between the flowers of plants commonly found in restinga, such as C. ovalifolia and C. ramosa, and their most important pollinators, such as C. decolorata and X. subcyanea, is vital in order to guarantee the survival of both organisms, and to consequently aid preservation and conservation programmes for dune and restinga ecosystems, since these environments have suffered an accelerated process of degradation due to human activity.
Acknowledgements We would like to thank the owners of the Mingu ranch for allowing this field study to be carried out. We would also like to thank Fernando C. V. Zanella (UFCG) and Edinaldo L. das Neves (FJA) for the identification of bee species; and Luciano Paganutti Queiróz (UEFS), for the identification of the plants studied.
Received July 29, 2009
Accepted June 4, 2010
Distributed May 31, 2011
- ALBUQUERQUE, PMC., CAMARGO, JMF. and MENDONÇA, JAC., 2007. Bee community of a beach dune ecosystem on Maranhão Island, Brazil. Brazilian Archives of Biology and Technology, vol. 50, p. 1005-1018.
- ALVES-DOS-SANTOS, I., 1999. Abelhas e plantas melíferas da mata atlântica, restinga e dunas do litoral norte do estado do Rio Grande do Sul. Revista Brasileira de Entomologia, vol. 43, p.191-223.
- Angiosperm Phylogeny Group II APG II, 2003. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG II - The angiosperm phylogeny group. Botanical Journal of the Linnean Society, vol. 141, p. 399-436. doi:10.1046/j.1095-8339.2003.t01-1-00158.x
- BRITTO, IC. and NOBLICK, LR., 1984. A importância de preservar as dunas de Itapoã e Abaeté. In LACERDA, LD., ARAUJO, DCD., CERQUEIRA, R. and TURCQ, B. (Org). Restinga: origem, estrutura e processos. Niterói: CEUFF. p. 269.-273.
- BUCHMANN, SL., 1983. Buzz pollination in angiosperms. In JONES, CE. and LITTLE, RJ. (Eds.), Handbook of experimental pollination biology. New York: Van Nostrand Reinhold Company. p. 73-113
- CLARKE, KR. and WARWICK, RM., 2001. Change in Marine Communities. An approach to statistical analysis and interpretation 2nd ed. Plymouth: Primer-E Ltd. p.190.
- COSTA, JAS. and RAMALHO, M., 2001. Ecologia da polinização em ambientes de Duna Tropical (APA do Abaeté, Salvador, Bahia, Brasil). Sitientibus série Ciências Biológicas, vol. 1, p. 135-145.
- CRONQUIST, A., 1981. An integrated system of classification of flowering plants New York: Columbia University Press. p. 1262.
- DAFNI, A.; KEVAN, P. G. and HUSBAND, B. C., 2005. Practical pollination biology Cambridge: Enviroquest, Ltd., p. 590.
- FAEGRI, K. and van der PIJL, L., 1979. The principles of pollination ecology. 3nd ed. London: Pergamon Press. p. 244.
- GENTRY, H.A. 1974. Flowering phenology and diversity in tropical Bignoniaceae. Biotropica, vol. 6, p.64-68. doi:10.2307/2989698
- GIMENES, M., OLIVEIRA-REBOUÇAS, P. and ALMEIDA, GF., 2007. Estudo das interações abelhas (Hymenoptera-Apoidae) e flores em um ecossistema de restinga no estado da Bahia, Brasil. Sitientibus Ciências Biológicas, vol. 7, p. 347-353.
- GOTTSBERGER, G., CAMARGO, JMF. and SILBERBAUERGOTTSBERGER, I., 1988. A bee-pollinated tropical community: The beach dune vegetation of Ilha de São Luís, Maranhão, Brazil. Botanische Jahrbücher für Systematik, vol. 109, p. 469-500.
- KREBS, CJ., 1989. Ecological Methodology New York: Harper Collins Publishers. 654 p.
- LACERDA, LD. and ESTEVES, FA., 2000. Restingas Brasileiras: Quinze anos de Estudo. Texto de apresentação. In ESTEVES, FA. and LACERDA, LD. (Eds). Ecologia das restingas e das Lagoas Costeiras. Rio de janeiro: NUPEM/UFRJ. p. 446
- LEWIS, G., SCHRIRE, B., MACKINDER, B. and LOCK, M. (Eds.). 2005. Legumes of the World Kew: Royal Botanic Gardens. p. 592.
- MAGURRAN, AE., 2003. Measuring biological diversity. Oxford: Blackwell Publishing. p. 256.
- MICHENER, CD., 2007. The bees of the Word 2nd ed. Baltimore: The Johns Hopkins University Press. 953 p.
- OLIVEIRA-REBOUÇAS, P. and GIMENES, M., 2004. Abelhas (Apoidea) visitantes de flores de Comolia ovalifolia DC Triana (Melastomataceae) em uma área de restinga na Bahia. Neotropical Entomology, vol. 33, p. 315-320.
- OLIVEIRA, FS., MENDONÇA, MWA., VIDIGAL, MCS., REGO, MMC. and ALBUQUERQUE, PMC., 2010. Comunidade de abelhas (Hymenoptera, Apoidea) em ecossistema de dunas na Praia de Panaquatira, São José de Ribamar, Maranhão, Brasil. Revista Brasileira de Entomologia, vol. 54, p. 8290. doi:10.1590/S0085-56262010000100010
- PINHEIRO, MCB., ORMOND, WT., LEITE CO. and LIMA, HA., 1988. Ecologia da polinização de Chamaecrista ramosa (Vog.) Irnn e Barn var. ramosa Revista Brasileira de Biologia = Brazilian Journal of Biology, vol. 48, p. 665-672.
- PINTO, GCP., BAUTISTA, HP. and FERREIRA, JDCA., 1984. A restinga do litoral nordeste do estado da Bahia. In LACERDA, LD., ARAUJO, DCD., CERQUEIRA, R. and TURCQ B. (Org). Restinga: origem, estrutura e processos. Niterói: CEUFF. p. 195- 216.
- Programa de Desenvolvimento Sustentável pra Áreas de proteção Ambiental do litoral Norte da Bahia - PRODESU, 2001. Relatório síntese p. 13-15.
- QUEIRÓZ, EP., 2007. Levantamento florístico e georreferenciamento das espécies com potencial econômico e ecológico em restinga de Mata de São João, Bahia, Brasil. Biotemas, vol. 20, p. 41-47.
- RAMALHO, M. and SILVA, M. 2002. Flores de óleo: Influência sobre a diversidade de abelhas Centridini em uma comunidade de Restinga Tropical. Sitientibus Ciências Biológicas vol. 2, p. 34-43.
- RENNER, SS., 1993. Phylogeny and classification of the Melastomataceae and Memecylaceae. Nordic Journal of Botany , vol. 13, p. 519-540. doi:10.1111/j.1756-1051.1993.tb00096.x
- RIBEIRO, EKMD., REGO, MMC. and MACHADO, ICS., 2008. Cargas polínicas de abelhas polinizadoras de Byrsonima chrysophylla Kunth. (Malpighiaceae): fidelidade e fontes alternativas de recursos florais1. Acta Botânica Brasilica, vol. 22, p. 165-171.
- ROCHA, CFD., BERGALLO, HG., VAN SLUYS, M., ALVES, MAS. and JAMEL, CE., 2007. The remnants of restinga habitats in the brazilian Atlantic Forest of Rio de Janeiro state, Brazil: Habitat loss and risk of disappearance. Brazilian Journal of Biology, vol. 67, p. 263-273.
- SILVA, SSL., ZICKEL CS. and CESTARO LA., 2008. Flora vascular e perfil fisionômico de uma restinga no litoral sul de Pernambuco, Brasil. Acta Botânica Brasilica, vol. 22, p.1123-1135.
- SILVA, MCM. and MARTINS, C., 1999. Flora apícola e relações tróficas de abelhas (Hymenoptera: Apoidea) em uma área de duna (praia de Interlagos, Cabedelo-PB, Brasil). Principia (CEFET-PB) vol. 7, p. 40-50.
- SILVEIRA, FA., MELO, GAR. and ALMEIDA, EAB., 2002. Abelhas Brasileira: Sistemática e Identificação. Belo Horizonte: Composição e Arte. p. 253.
- SILVEIRA NETO, S., NAKANO, O., BARBIN, D. and NOVA, NAV., 1976. Manual de ecologia dos insetos São Paulo: Ed. Agronômica Ceres. p. 419.
- THOMAZINI, MJT. and THOMAZINI, APBW., 2002. Diversidade de Abelhas (Hymenoptera: Apoidea) em Inflorescências de Piper hispidinervum (C.DC.) Neotropical Entomology, vol. 31, p. 27-34.
- THORP, RW. and ESTES, JR., 1975. Intrafloral behavior of bees on flowers of Cassia fasciculate Journal of the Kansas Entomological Society, vol. 48, p. 175- 184.
- VIANA, BF., SILVA, FO. and KLEINERT, AMP., 2005. A community of flower-visiting bees (Hymenoptera: Apoidea) in the coastal sand dunes of northeastern Brazil. Biota Neotropica, vol. 5, p. 1-13.
- -, 2006. Structure of bee-flower system in the coastal sand dune of Abaeté northeastern Brazil. Revista Brasileira de Entomologia, vol. 50, p. 53-63.
- VIANA, BF., SILVA, FO. and KLEINERT, AMP. 2006. A flora apícola de uma área restrita de dunas litorâneas, Abaeté, Salvador, Bahia. Revista Brasileira de Botânica, vol. 29, p.13-25.
- VOGEL, S., 1978. Evolutionary shifts from reward to deception in pollen flowers. In The Pollination of Flowers by Insects. Linnean Society Symposium Series, vol. 6, p. 89-96.
- ZANELLA, FCV., SCHWARTZ FILHO, DL. and LAROCA, S., 1998. Tropical bee island biogeography: diversity and abundance patterns. Biogeographica, vol. 74, no. 3, p. 103-15.
- ZAR, JH., 2010. Biostatistical analysis. 5 nd ed. NewJersey: Prentice-Hall, Upper Saddle River. p. 944.
Publication Dates
-
Publication in this collection
15 July 2011 -
Date of issue
May 2011
History
-
Received
29 July 2009 -
Accepted
04 June 2010