Abstracts
Although crab spiders are common in flowering plants, their relations with plant species and its floral traits have been poorly known in the Neotropics. Observations regarding plant habits, floral visitors and also floral characteristics such as anthesis, odour, shape, colour and floral resources were recorded in flowering plant species of an area of "Cerrado" on a 2 km long trail. Misumenops argenteus and Misumenops pallens accounted for 62.86% of the spiders captured on 22 flowering plant species. The plants Senna rugosa (Fabaceae), Styrax ferrugineus (Styracaceae) and Banisteriopsis campestris (Malpighiaceae), hosted, each one, about 10 to 17% of the total spiders collected and these plants had diurnal anthesis, bee-attractive flower colours such as yellow (S. rugosa), white (S. ferrugineus), and pink (B. campestris), poricidal anthers as well as being visited by bees which evidenced bee-pollination syndrome. This study is the first survey regarding crab spiders and their associations with plant species of the "Cerrado".
ambush spiders; Misumenops; floral biology; floral visitors
Ainda que aranhas Thomisidae sejam comumente encontradas em flores, as associações desses aracnídeos a espécies de plantas e às suas características florais foram pouco registradas na região neotropical. Observações do hábito das plantas, visitantes florais, e também das características florais, tais como antese, odor, forma, cor e recursos da flor, foram assinaladas para espécies floridas de uma área de cerrado presentes em uma trilha de 2 km de extensão. Misumenops argenteus e Misumenops pallens representaram 62,86% das aranhas habitantes de 22 espécies de plantas floridas. As plantas Senna rugosa (Fabaceae), Styrax ferrugineus (Styracaceae) e Banisteriopsis campestris (Malpighiaceae) abrigaram, individualmente, cerca de 10 a 17% do total das aranhas e, nestas plantas, a antese diurna; flores de coloração atrativa a abelhas, como amarela (S. rugosa), branca (S. ferrugineus) e rosa (B. campestris) e as anteras poricidas, bem como a visita das flores por abelhas reforçou a evidência de síndrome de polinização para melitofilia. Este é o primeiro levantamento de espécies de aranhas Thomisidae associadas a plantas do cerrado brasileiro.
aranhas caçadoras de emboscada; Misumenops; biologia floral; visitantes florais
ECOLOGY
Crab spiders (Araneae: Thomisidae) in flowering plants in a Brazilian "Cerrado" ecosystem
Aranhas Thomisidae em plantas floridas de um ecossistema de Cerrado
Rocha-Filho, LC.I,* * e-mail: correiadarocha@yahoo.com.br ; Rinaldi, IMP.II
IDepartamento de Biologia, Faculdade de Filosofia, Ciências e Letras de Ribeirão Preto FFCLRP, Universidade de São Paulo USP, Av. Bandeirantes, 3900, CEP 14040-901, Ribeirão Preto, SP, Brazil
IIDepartamento de Zoologia, Instituto de Biociências, Universidade Estadual Paulista UNESP, CP 510, CEP 18618-000, Botucatu, SP, Brazil
ABSTRACT
Although crab spiders are common in flowering plants, their relations with plant species and its floral traits have been poorly known in the Neotropics. Observations regarding plant habits, floral visitors and also floral characteristics such as anthesis, odour, shape, colour and floral resources were recorded in flowering plant species of an area of "Cerrado" on a 2 km long trail. Misumenops argenteus and Misumenops pallens accounted for 62.86% of the spiders captured on 22 flowering plant species. The plants Senna rugosa (Fabaceae), Styrax ferrugineus (Styracaceae) and Banisteriopsis campestris (Malpighiaceae), hosted, each one, about 10 to 17% of the total spiders collected and these plants had diurnal anthesis, bee-attractive flower colours such as yellow (S. rugosa), white (S. ferrugineus), and pink (B. campestris), poricidal anthers as well as being visited by bees which evidenced bee-pollination syndrome. This study is the first survey regarding crab spiders and their associations with plant species of the "Cerrado".
Keywords: ambush spiders, Misumenops, floral biology, floral visitors.
RESUMO
Ainda que aranhas Thomisidae sejam comumente encontradas em flores, as associações desses aracnídeos a espécies de plantas e às suas características florais foram pouco registradas na região neotropical. Observações do hábito das plantas, visitantes florais, e também das características florais, tais como antese, odor, forma, cor e recursos da flor, foram assinaladas para espécies floridas de uma área de cerrado presentes em uma trilha de 2 km de extensão. Misumenops argenteus e Misumenops pallens representaram 62,86% das aranhas habitantes de 22 espécies de plantas floridas. As plantas Senna rugosa (Fabaceae), Styrax ferrugineus (Styracaceae) e Banisteriopsis campestris (Malpighiaceae) abrigaram, individualmente, cerca de 10 a 17% do total das aranhas e, nestas plantas, a antese diurna; flores de coloração atrativa a abelhas, como amarela (S. rugosa), branca (S. ferrugineus) e rosa (B. campestris) e as anteras poricidas, bem como a visita das flores por abelhas reforçou a evidência de síndrome de polinização para melitofilia. Este é o primeiro levantamento de espécies de aranhas Thomisidae associadas a plantas do cerrado brasileiro.
Palavras-chave: aranhas caçadoras de emboscada, Misumenops, biologia floral, visitantes florais.
1. Introduction
Spiders, an important predator community, are one of the most diverse and abundant groups within Arthropoda, with 40,998 described species (Platnick, 2009) that are found throughout all terrestrial ecosystems and zoogeographical regions of the world, from Arctic islands to arid and desert regions (Turnbull, 1973; Foelix, 1996). The geographical distribution of spiders is intimately associated with various biotic and abiotic factors, such as temperature, humidity, luminosity, vegetation, and food availability (Turnbull, 1973; Wise, 1995; Foelix, 1996; Gonzaga et al., 2007).
The family Thomisidae has 2,093 species, of which the largest are tropical, 12-20 mm in body length. They hunt prey, including large-sized insects, by ambushing (Morse, 1984; Foelix, 1996; Wise, 1995). The genera Misumenoides F. O. Pickard-Cambridge, 1900 and Misumenops F. O. Pickard-Cambridge, 1900 are found on flowers and leaves, and have a very broad range of color patterns (Morse and Fritz, 1982). Females of such spiders are often white or yellow with red and brown dotted patterns, using this camouflage to hide from predators and prey. Gertsch (1939, p. 293) described them as being "[...] commonly found on corolla or white flower petal sets, where they remain waiting for insects that visit flowers to obtain either pollen or nectar [...]". According to some authors (Schick, 1965; Gonzaga et al., 2007), crab spiders are more diverse in temperate, subtropical and tropical areas, such as those of the "Cerrado" ecosystems.
"Cerrado" (Brazilian savanna, sensu stricto) is the second largest Brazilian vegetational formation, with a total area of approximately two million square kilometres (Oliveira-Filho and Ratter, 2002); it is considered a hotspot because of its endemic species and constant threats to its territory (Orme et al., 2005). There are few reports in literature of plants on which Thomisidae spiders may be found. Lowrie (1967) related some associations of Misumena vatia (Clerck, 1757) with vegetal species, and Schick (1965) reported plants associated with some species of Misumenops in California. Preferences for plants have been described by Chew (1961) for Misumenops coloradensis Gertsch, 1933, within a shrub community in Arizona, and Jennings (1971) related associations of such species with plants in New Mexico and researched their seasonality and altitudinal distribution. In the Neotropics, Romero and Vasconcellos-Neto (2003, 2004) reported the phenology of Misumenops argenteus (Mello-Leitão, 1929) on Trichogoniopsis adenantha (DC) (Asteraceae) in a subtropical semideciduous mesophilic forest area in Brazil.
Although some studies have shown a double effect of crab spiders on plants since they can prey both on phytophagous insects and pollinators (Morse and Fritz, 1982; Young, 1989; Romero and Vasconcellos-Neto, 2004; González et al., 2009), their interactions with herbivores, seed predators, pollinators, and floral visitors is still poorly known. In order to better enhance knowledge of the crab spiders of the "Cerrado" domain and its preference for plants species of this biome, we reported the occurrence of Thomisidae spiders on flowering plant species of a Brazilian "Cerrado" sensu stricto ecosystem, evaluating their interaction with vegetal habits and floral characteristics like resources, odour, flower shape and colour, besides anthesis and floral visitors. Furthermore, considering its high levels of endemism and biodiversity, the "Cerrado" is underexplored (Oliveira-Filho and Ratter, 2002) and as pointed out by Del-Claro and Torezan-Silingardi (2009), this biome provides propitious conditions for the study of floral herbivore-predator-pollinator interactions mainly due to its high insect diversity.
2. Material and Methods
The study was carried out within the reserve RPPN do Clube Caça & Pesca Itororó in Uberlândia, Minas Gerais State, Brazil (18° 55' S and 48° 17' W). The reserve has an area of 127 ha with "Cerrado" sensu stricto and a secondary-forest field ("campo sujo") (Del-Claro et al., 1997; Appolinário and Schiavini, 2002). Two well defined seasons, dry and humid, characterise the regional climate. Pluviosity was around 1,550 mm annually, and average temperature is 22 °C (Nimer and Brandão, 1989). Along a 2 km-long patch, visits were made twice a month, from August 2003 to July 2004, from 7:30 AM to 11:30 AM; totalising 24 samplings. All spiders present in flowering plant species found at 1 m on the left and right sides off the patch were photographed, identified by the second author, collected by handling and deposited in the Spider Collection at the Zoology Department, UNESP, Botucatu, state of São Paulo (Coleção UBTU "http://splink.cria.org.br"). The plants were identified and analysed according to the Faegri and Van Der Pijl (1979) criteria and the habit and floral features of each plant were monitored. Observations of five minutes were carried out on the reproductive branches of the flowering plant species found in the sampling area in order to identify its floral visitors.
3. Results
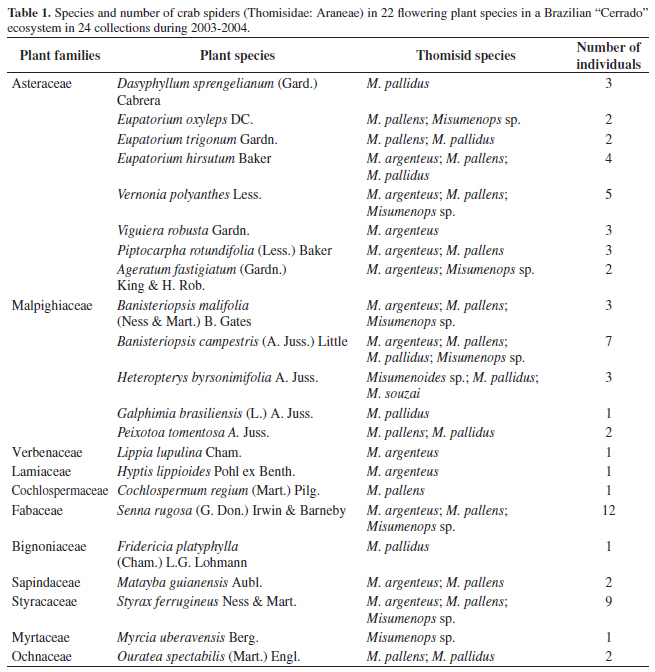
In a total of 70 individuals from six Thomisidae species, 22 "Cerrado" plant species were registered. They were: 23 individuals of Misumenops argenteus (1♀ + 22 sub-adults); 21 of Misumenops pallens (Keyserling, 1880) (3♂ + 18 sub-adults); 11 of Misumenops pallidus (Keyserling, 1880) (1♂ + 10 sub-adults); 1♀ of Misumenops souzai (Soares, 1942), 13 juveniles of Misumenops sp., and one juvenile of Misumenoides sp. (Table 1).
From 22 species of plants in which crab spiders were found, the most diverse families were Asteraceae and Malpighiaceae with eight and five species respectively, and each of the remaining plant species belonged to a single family (Table 1).
Individuals of Misumenops argenteus and M. pallens occurred throughout the entire study period, except for the first two observation months, for M. argenteus, and in October, 2003, for M. pallens (Figure 1). Misumenops souzai and a nymph of Misumenoides sp. occurred on flowers only in August, 2003. Specimens of M. pallidus were collected from August to December, 2003, and only one individual of such species was again reported in April, 2004. Misumenops nymphs were observed from December, 2003 to the last collection month, July, 2004, being absent only in April, 2004 (Figure 1).
Most plants (59.09%) were sub-shrubs; herbs and shrubs represented 36.36% of total and only Styrax ferrugineus Ness. & Mart. (Styracaceae) was a tree (Table 2).
Regarding flower shape, 50% were bell-like and 40.9% were tube-like, whereas 9.09% were throat-like. About 73% of the plant species had odourless flowers, 63.64% produced pollen and 54.55% produced nectar. Floral oils were found in 22.73% of the plants flowering. Five distinct colours were observed prevailing: yellow (31.82%), white (22.73%) and pink (22.73%). The more attractive flowers to bees were beige (13.64%) and lilac (9.09%). Floral traits of all studied plants evidenced melittophilic pollination syndrome, such as daily anthesis (100.0%) and yellow and white colours (54.55%), as established by Faegri and Van Der Pijl (1979) (Table 2).
Asteraceae and Malpighiaceae were the most diverse families, with eight and five species, respectively and 54.14% of spiders were collected on them. Plants with the greatest spider abundance were Senna rugosa (Fabaceae) with 17.14%; Styrax ferrugineus (Styracaceae) with 12.86% and Banisteriopsis campestris (Malpighiaceae) with 10% of the all spiders.
Floral visitors were flies, butterflies, wasps, beetles and predominantly bees which were registered in 15 species of plants (Table 2). Predations in the field area on visitors were observed in inflorescences of Dasyphyllum sprengelianum, where Misumenops pallidus attacked on a Phoridae (Diptera), and in Matayba guianensis, when a Misumenops pallens attacked on an Apis mellifera Linnaeus, 1758 (Apidae: Hymenoptera).
4. Discussion
Distribution, abundance and aggregation patterns of spiders are a function of several factors including the architecture of vegetative branches (Gunnarsson, 1996; Evans, 1997, Halaj et al., 2000; Romero and Vasconcellos-Neto, 2004). Inflorescences are clusters of flowers with a complex arrangement of branches and are suitable for shelter or as reproductive sites (Nentwig, 1993), for basic sources of pollen and nectar to pollinating insects and other phytophagous which can be strongly selected as hunting sites for plant-dwelling spiders (Souza and Martins, 2004; Louda, 1982; Morse and Fritz, 1982; Riechert and Tracy, 1975). Inflorescences also can offer pollen and nectar to spider nymphs in periods of lack or scarcity of prey-insects (Vogelei and Greissel, 1989) or hidden very small floral herbivores (Del-Claro et al., 1997; Del-Claro and Torezan-Silingardi, 2009). All of the studied plant species had dense, large inflorescences instead of isolated flowers and received insect visitors, what is reasonable to explain that crab spiders occurred abundantly in flowering plants.
We observed that some aspects of the inflorescences can be related to the presence of crab spiders. Number of flowers of an inflorescence and flower traits influence the abundance of visiting insects and this is crucial for spiders to select hunting sites (Bernays and Chapman, 1994; Greco and Kevan, 1994). The greatest amount of crab spiders per plant species was registered in Senna rugosa, Styrax ferrugineus and Banisteriopsis campestris. All of these plants have diurnal anthesis, bee-attractive flower colours as yellow (S. Rugosa), white (S. Ferrugineus) and rose (B. Campestris), and poricidal anthers, which are necessary conditions for the bee-pollination syndrome. Flowers of Senna rugosa are pollinated by bees that bear the "buzz-pollination" peciali (Buchmann and Hurley, 1978). Styrax ferrugineus has yellow and conspicuous stamina and poricidal anthers such as in Senna rugosa, which makes pollen accessible only to buzzing bees (Buchmann, 1983). In turn, Banisteriopsis campestris is a species that secretes floral oils through pecialized glands, elaiophores (Simpson and Neff, 1983). Such oils are collected by females of certain bee groups that use them to build and/or feed larvae (Vinson et al., 1997). Our observations suggest that crab spiders are more abundant on melittophilous, bee-visited flowers. Further experiments are needed in order to obtain more information about the ecology of crab spiders in flowering plants of the "Cerrado" areas given that, as Louda (1982) pointed out, predators that dwell on plants also highly influence the plant reproductive biology.
Acknowledgements The authors would like to thank Dr. Ana Angélica Almeida Barbosa, Dr. Jimi Naoki Nakajima (Federal University of Uberlândia) and Dr. Adriana Assis Arantes (University Anhembi Morumbi) for their help in identifying plants, and Dr. Raoul Henry (São Paulo State University) for his critique and suggestions for the manuscript.
Received November 10, 2009
Accepted March 15, 2010
Distributed May 31, 2011
- APPOLINÁRIO, V. and SCHIAVINI, I., 2002. Levantamento fitossociológico de espécies arbóreas de Cerrado (stricto sensu) em Uberlândia - Minas Gerais. Boletim do Herbário Ezechias Paulo Heringer, vol. 10, p. 57-75.
- BERNAYS, EA. and CHAPMAN, RF., 1994. Host-plant selection by phytophagous insects New York: Chapman & Hall. 312 p.
- BUCHMANN, SL., 1983. Buzz pollination in Angionsperms. In JONES, C. and LITTLE, J. (Eds.). Handbook of experimental pollination biology New York: Van Nostrand Reinhold. 558 p.
- BUCHMANN, SL. and HURLEY, JP., 1978. A biological model for buzz pollination in angiosperms. Journal of Theoretical Biology, vol. 72, p. 639-657. doi:10.1016/0022-5193(78)90277-1
- CHEW, RM., 1961. Ecology of the spiders of a desert community. Journal of The New York Entomological Society, vol. 69, p. 5-41.
- DEL-CLARO, K. and TOREZAN-SILINGARDI, HM., 2009. Insect-plant interactions: New pathways to a better comprehension of ecological communities in Neotropical savannas. Neotropical Entomology, vol. 38, 159-164. PMid:19488503. doi:10.1590/S1519-566X2009000200001
- DEL-CLARO, K., MARULLO, R. and MOUND, LA., 1997. A new Brazilian species of Heterothrips (Insecta: Thysanoptera) co-existing with ants in the flowers of Peixotoa tomentosa (Malpighiaceae). Journal of Natural History, vol. 31, p. 1307-1312. doi:10.1080/00222939700770731
- EVANS, TA., 1997. Distribution of social crab spiders in eucalypt forests. Australian Journal of Ecology, vol. 22, p. 107-111. doi:10.1111/j.1442-9993.1997.tb00646.x
- FAEGRI, K. and VAN DER PIJL, L., 1979. The principles of pollination ecology London: Pergamon Press. 244 p.
- FOELIX, RF., 1996. Biology of Spiders New York: Oxford University Press. 330 p.
- GERTSCH, WJ., 1939. A revision of the typical crab-spiders (Misumeninae) of America north of Mexico. Bulletin of the American Museum of Natural History, vol. 76, p. 277-442.
- GONZAGA, MO., SANTOS, AJ. and JAPYASSÚ, HF., 2007. Ecologia e comportamento de aranhas Rio de Janeiro: Editora Interciência. 400 p.
- GONZÁLEZ, A., LILJESTHRÖM, G., MINERVINO, E., CASTRO, D., GONZÁLEZ, S. and ARMENDANO, A., 2009. Predation by Misumenops pallidus (Araneae: Thomisidae) on insect pests of soybean cultures in Buenos Aires Province, Argentina. Journal of Arachnology, vol. 37, p. 282-286. doi:10.1636/T08-63.1
- GRECO, CF. and KEVAN, PG., 1994. Contrasting patch choosing by anthophilous ambush predators: vegetation and floral cues for decisions by a crab spider (Misumena vatia) and males and females of an ambush bug (Phymata americana). Canadian Journal of Zoology, vol. 72, p. 1583-1588. doi:10.1139/z94-210
- GUNNARSSON, B., 1996. Bird predation and vegetation structure affecting spruce-living arthropods in a temperate forest. Journal of Animal Ecology, vol. 65, p. 389-397. doi:10.2307/5885
- HALAJ, J., ROSS, DW. and MOLDENKE, AR., 2000. Importance of habitat structure to the arthropod food-web in Douglas-fir canopies. Oikos, vol. 90, p. 139-152.
- JENNINGS, DT., 1971. Plant associations of Misumenops coloradensis Gertsch (Araneae: Thomisidae) in Central New Mexico. The Southwestern Naturalist, vol. 16, p. 201-207. doi:10.2307/3670499
- LOUDA, SM., 1982. Inflorescence spiders: a cost/benefit analysis for the host plant, Haplopappus venetus Blake (Asteraceae). Oecologia, vol. 55, p. 185-191. doi:10.1007/BF00384486
- LOWRIE, DC., 1967. Effects of grazing and intensive collecting on a population of the green lynx spider. Ecology, vol. 44, p. 777-781.
- MORSE, DH., 1984. How crab spiders (Araneae, Thomisidae) hunt at flowers. Journal of Arachnology, vol. 12, p. 307-316. doi:10.2307/1937042
- MORSE, DH. and FRITZ, RS., 1982. Experimental and observational studies of patch choice at different scales by the crab spider Misumena vatia. Ecology, vol. 63, p. 172-182.
- NENTWIG, W., 1993. Spiders of Panama Gainesville: Sandhill Crane Press. 274 p.
- NIMER, E. and BRANDÃO, AMPM., 1989. Balanço hídrico e clima da região dos cerrados Rio de Janeiro: Fundação IBGE. 166 p.
- OLIVEIRA-FILHO, AT. and RATTER, JA., 2002. Vegetation physiognomies and woody flora of the cerrado biome. In OLIVEIRA, PS. and MARQUIS, RJ. (Eds.). The cerrados of Brazil: Ecology and natural history of a Neotropical savanna. New York: Columbia University Press. 398 p.
- ORME, CDL. et al., 2005. Global hotspots of species richness are not congruent with endemism or threat. Nature, vol. 436, p. 1016-1019. PMid:16107848. doi:10.1038/nature03850
- PLATNICK, NI., 2009. The world spider catalog, version 10.0. Available from:<http://research.amnh.org/entomology/spiders/catalog/index.html>
- RIECHERT, SE. and TRACY, CR., 1975. Thermal balance and prey availability: bases for a model relating web-site characteristics to spider reproductive success. Ecology, vol. 56, p. 265-284. doi:10.2307/1934960
- ROMERO, GQ. and VASCONCELLOS-NETO, J., 2003. Natural history of Misumenops argenteus (Thomisidae): Seasonality and diet on Trichogoniopsis adenantha (Asteraceae). Journal of Arachnology, vol. 31, p. 297-304. doi:10.1636/02-19
- -, 2004. Beneficial effects of flower-dwelling predators on their host plant. Ecology, vol. 85, p. 446-457. doi:10.1890/02-0327
- SCHICK, RX., 1965. The crab spiders of California (Araneidae, Thomisidae). Bulletin of the American Museum of Natural History, vol. 129, p. 1-180.
- SIMPSON, BB. and NEFF, JL., 1983. Evolution and diversity of floral rewards In JONES, C. and LITTLE, J. (Eds.). Handbook of experimental pollination biology New York: Van Nostrand Reinhold. 558 p.
- SOUZA, ALT. and MARTINS, RP., 2004. Distribution of plant-dwelling spiders: inflorescences versus vegetative branches. Austral Ecology, vol. 29, p. 342-349. doi:10.1111/j.1442-9993.2004.01371.x
- TURNBULL, AL., 1973. Ecology of the true spiders (Araneomorphae). Annual Review of Entomology, vol. 18, p. 305-348. doi:10.1146/annurev.en.18.010173.001513
- VINSON, SB., WILLIAMS, HJ., FRANKIE, GW. and SHRUM, G., 1997. Floral lipid chemistry of Byrsonima crassifolia (Malpighiaceae) and a use of floral lipids by Centris bees (Hymenoptera, Apidae). Biotropica, vol. 29, p. 76-83. doi:10.1111/j.1744-7429.1997.tb00008.x
- VOGELEI, A. and GREISSEL, R., 1989. Survival strategies of the crab spider Thomisus onustus Walckenaer, 1806 (Chelicerata, Arachnida, Thomisidae). Oecologia, vol. 80, p. 513-515. doi:10.1007/BF00380075
- WISE, DH., 1995. Spiders in ecological webs New York: Cambridge University Press. 328 p.
- YOUNG, OP., 1989. Relationships between Aster pilosus (Compositae), Misumenops spp. (Araneae: Thomisidae), and Ligus lineolaris (Heteroptera: Miridae). Journal of Entomological Sciences, vol. 24, p. 252-257.
Publication Dates
-
Publication in this collection
15 July 2011 -
Date of issue
May 2011
History
-
Received
10 Nov 2009 -
Accepted
15 Mar 2010