Abstracts
Hummingbird communities tend to respond to variation in resources, having a positive relationship between abundance and diversity of food resources and the abundance and/or diversity of hummingbirds. Here we examined the influence of floral resource availability, as well as seasonality and type of habitat on the composition of hummingbird species. The study was carried out in two habitats of eastern Brazilian mountaintops. A gradient representative of the structure of hummingbird community, based on species composition, was obtained by the ordination of samples using the method of non-metric multidimensional scaling. The composition of hummingbird species was influenced by the type of habitat and floral resource availability, but not by seasonality. Hummingbird communities differ between habitats mainly due to the relative abundance of hummingbird species. The variation in composition of hummingbird species with the variation in floral resource availability may be related to differences in feeding habits of hummingbirds. Hummingbird species with the longest bills visited higher proportions of ornithophilous species, while hummingbirds with shorter bills visited higher proportions of non-ornithophilous species. The results demonstrate that at local-scale the composition of hummingbird species is affected by the type of habitat and floral resources availability, but not by seasonality.
Brazil; hummingbirds; campos rupestres ; habitat preference; seasonality; Serra do Cipó
Comunidades de beija-flores tendem a responder a variação em recursos, havendo uma relação positiva entre a abundância e diversidade de recurso alimentar com a abundância e/ou diversidade de beija-flores. Neste estudo nós analisamos a influencia da disponibilidade de recurso floral, bem como da sazonalidade e do tipo de habitat na composição de espécies de beija-flores. O estudo foi realizado em dois habitas de montanha do leste do Brasil. Um gradiente representativo da estrutura da comunidade de beija-flores, baseado na composição de espécies, foi obtido através da ordenação das amostras usando o método de escalonamento multidimensional não-métrico. A composição de espécies de beija-flores foi influenciada pelo tipo de habitat e disponibilidade de recurso floral, mas não pela sazonalidade. As comunidades de beija-flores variaram entre os habitas, principalmente devido a abundância relativa das espécies de beija-flores. A variação na composição de espécies de beija-flores com a variação na disponibilidade de recurso floral parece estar relacionada a diferenças nos hábitos alimentares dos beija-flores. Espécies de beija-flores com bicos de comprimentos maiores visitaram maiores proporções de espécies ornitófilas, enquanto beija-flores com bicos de comprimento menores visitaram maiores proporções de espécies não-ornitófilas. Os resultados demonstram que em escala local a composição de espécies de beija-flores é afetada pelo tipo de habitat e a disponibilidade de recurso floral, mas não pela sazonalidade.
Brasil; beija-flores; campos rupestres; preferência de habitat; sazonalidade; Serra do Cipó
1 Introduction
In bird communities the species richness and abundance of individuals may vary spatially and
temporally (Malizia, 2001Malizia, LC., 2001. Seasonal fluctuations of birds, fruits, and flowers in a
subtropical forest of Argentina. The Condor, vol. 103, no. 1, p. 45-61.
http://dx.doi.org/10.1650/0010-5422(2001)103[0045:SFOBFA]2.0.CO;2.
http://dx.doi.org/10.1650/0010-5422(2001...
) due to population processes
(births and deaths) and individual movements within and between habitats (Loiselle and Blake, 1991Loiselle, BA. and Blake, JG., 1991. Temporal variation in birds and fruits along
an elevational gradient in Costa Rica. Ecology, vol. 72, no. 1, p. 180-193.
http://dx.doi.org/10.2307/1938913.
http://dx.doi.org/10.2307/1938913...
; Poulin et al.,
1993Poulin, B., Lefebvre, G. and Mcneil, R., 1993. Variations in bird abundance in
tropical arid and semiarid habitats. The Ibis, vol. 135, no. 4, p. 432-441.
http://dx.doi.org/10.1111/j.1474-919X.1993.tb02116.x.
http://dx.doi.org/10.1111/j.1474-919X.19...
). The main reasons for movement are related to variation in resource availability,
habitat structure, climates conditions and competition (Montgomerie and Gass, 1981Montgomerie, RD. and Gass, CL., 1981. Energy limitation of hummingbird
populations in tropical and temperate communities. Oecologia, vol. 50, no. 2, p. 162-165.
http://dx.doi.org/10.1007/BF00348031.
http://dx.doi.org/10.1007/BF00348031...
; Wiens, 1989Wiens, JA., 1989. A case study: interactions among nectarivores. In BARNES,
RSK., BIRKS, HJB., CONNOR, EF. and PAINE, RT. (Eds.). The ecology of bird communities.
Cambridge: Cambridge University Press. 316 p. Processes and variations, vol.
2.; Malizia, 2001Malizia, LC., 2001. Seasonal fluctuations of birds, fruits, and flowers in a
subtropical forest of Argentina. The Condor, vol. 103, no. 1, p. 45-61.
http://dx.doi.org/10.1650/0010-5422(2001)103[0045:SFOBFA]2.0.CO;2.
http://dx.doi.org/10.1650/0010-5422(2001...
). However, food resources are the most
important feature that affects the distribution pattern of many birds (Levey, 1988Levey, DJ., 1988. Spatial and temporal variation in Costa Rican fruit and
fruit-eating bird abundance. Ecological Monographs, vol. 58, no. 4, p. 251-269.
http://dx.doi.org/10.2307/1942539.
http://dx.doi.org/10.2307/1942539...
; Loiselle and Blake,
1991Loiselle, BA. and Blake, JG., 1991. Temporal variation in birds and fruits along
an elevational gradient in Costa Rica. Ecology, vol. 72, no. 1, p. 180-193.
http://dx.doi.org/10.2307/1938913.
http://dx.doi.org/10.2307/1938913...
; Malizia, 2001Malizia, LC., 2001. Seasonal fluctuations of birds, fruits, and flowers in a
subtropical forest of Argentina. The Condor, vol. 103, no. 1, p. 45-61.
http://dx.doi.org/10.1650/0010-5422(2001)103[0045:SFOBFA]2.0.CO;2.
http://dx.doi.org/10.1650/0010-5422(2001...
). For many species, the
capture rates of frugivorous birds have been positively correlated with fruit availability
(Loiselle and Blake, 1991Loiselle, BA. and Blake, JG., 1991. Temporal variation in birds and fruits along
an elevational gradient in Costa Rica. Ecology, vol. 72, no. 1, p. 180-193.
http://dx.doi.org/10.2307/1938913.
http://dx.doi.org/10.2307/1938913...
).
Hummingbirds are very mobile birds (Hadley and Betts,
2009Hadley, AS. and Betts, MG., 2009. Tropical deforestation alters hummingbird
movement patterns. Biology Letters, vol. 5, no. 2, p. 207-210.
http://dx.doi.org/10.1098/rsbl.2008.0691. PMid:19158031
http://dx.doi.org/10.1098/rsbl.2008.0691...
) highly dependent on nectar and have high energy requirements (Diamond et al., 1986Diamond, JM., Karasov, WH., Phan, D. and Carpenter, FL., 1986. Digestive
physiology is a determinant of foraging bout frequency in hummingbirds. Nature, vol. 320, no.
6057, p. 62-63. http://dx.doi.org/10.1038/320062a0. PMid:3951548
http://dx.doi.org/10.1038/320062a0...
). Thus, the availability of flowers may
lead the movements of hummingbirds within and between habitats in search of these resources
(Montgomerie and Gass, 1981Montgomerie, RD. and Gass, CL., 1981. Energy limitation of hummingbird
populations in tropical and temperate communities. Oecologia, vol. 50, no. 2, p. 162-165.
http://dx.doi.org/10.1007/BF00348031.
http://dx.doi.org/10.1007/BF00348031...
).
Moreover, the diversity and composition of hummingbird species may vary between biomes due to
changes in climate conditions, as well as precipitation and temperature (Abrahamczyk and Kessler, 2010Abrahamczyk, S. and Kessler, M., 2010. Hummingbird diversity, food niche
characters, and assemblage composition along a latitudinal precipitation gradient in the
Bolivian lowlands. Journal für Ornithologie, vol. 151, no. 3, p. 615-625.
http://dx.doi.org/10.1007/s10336-010-0496-x.
http://dx.doi.org/10.1007/s10336-010-049...
). Locally, hummingbird community tends to
respond to changes in resource availability. It has been found a positive relationship between
abundance and/or diversity of food resources and abundance and/or diversity of hummingbirds
(Feinsinger, 1976Feinsinger, P., 1976. Organization of a tropical guild of nectarivorous birds.
Ecological Monographs, vol. 46, no. 3, p. 257-291.
http://dx.doi.org/10.2307/1942255.
http://dx.doi.org/10.2307/1942255...
; Des
Granges, 1979Des Granges, JL., 1979. Organization of a tropical nectar feeding bird guild in
a variable environment. Living Bird, vol. 17, p. 199-236.; Montgomerie and Gass, 1981Montgomerie, RD. and Gass, CL., 1981. Energy limitation of hummingbird
populations in tropical and temperate communities. Oecologia, vol. 50, no. 2, p. 162-165.
http://dx.doi.org/10.1007/BF00348031.
http://dx.doi.org/10.1007/BF00348031...
; Stiles, 1985Stiles, FG., 1985. Seasonal pattern and coevolution in the hummingbird-flower
community of a Costa Rican sub-tropical forest. In BUCKLEY, PA., MORTON, MS., RIDGLEY, RS. and
BUCKLEY, FG. (Eds.). Neotropical ornithology. Washington: American Ornithologists Union. p.
757-787.; Malizia,
2001Malizia, LC., 2001. Seasonal fluctuations of birds, fruits, and flowers in a
subtropical forest of Argentina. The Condor, vol. 103, no. 1, p. 45-61.
http://dx.doi.org/10.1650/0010-5422(2001)103[0045:SFOBFA]2.0.CO;2.
http://dx.doi.org/10.1650/0010-5422(2001...
; Cotton, 2007Cotton, PA., 2007. Seasonal resource tracking by Amazonian hummingbirds. The
Ibis, vol. 149, no. 1, p. 135-142.
http://dx.doi.org/10.1111/j.1474-919X.2006.00619.x.
http://dx.doi.org/10.1111/j.1474-919X.20...
). However, except for the
study of Montgomerie and Gass (1981)Montgomerie, RD. and Gass, CL., 1981. Energy limitation of hummingbird
populations in tropical and temperate communities. Oecologia, vol. 50, no. 2, p. 162-165.
http://dx.doi.org/10.1007/BF00348031.
http://dx.doi.org/10.1007/BF00348031...
, who verified that
the number of hummingbirds is positively correlated with daily energy available, measured from
the daily nectar production per flower, no study so far verified the influence of energy
availability of floral resources, as well as rainfall and the type of vegetation on the
composition of hummingbird species.
We aim to test the hypothesis that hummingbird species composition varies according to floral resource availability among habitats and season in the open mountaintops of southeastern Brazil. We describe the seasonal variation of the hummingbird species richness and composition in two different habitats (‘typical campos rupestres’ or open rocky fields, and ‘capões de mata’ or forest patches) and its correlation to floral energetic resources.
2 Methods
2.1 Study site
This study was conducted in a region known as Alto do Palácio (hereafter AP, 19° 15’ S and 43° 31’ W, at approximately 1350 m above sea level), which is located in the northern part of the Serra do Cipó National Park, southeastern Brazil (Rodrigues et al., 2011RODRIGUES, M., Freitas, GHS., Costa, LM., Dias, DF., Varela, MLM. and Rodrigues, LC., 2011. Avifauna of Alto do Palácio, Serra do Cipó National Park, state of Minas Gerais, southeastern Brazil. Check List, vol. 7, p. 151-161.).
The AP region is characterized by campos rupestres habitats. In this region, the landscape is a mosaic consisting of the following characteristics: (1) typical campos rupestres or open rocky fields, which are areas of rocky outcrops with herbaceous vegetation and shrubs; (2) open grasslands composed predominantly of herbaceous species; and (3) capões de mata or forest patches, which are small areas of dense forest-like vegetation associated to wetter areas, such as springs and small streams (Rodrigues and Rodrigues, 2011RODRIGUES, LC. and Rodrigues, M., 2011. Size dimorphism, juvenal plumage, and timing of breeding of the hyacinth Visorbearer (). Augastes scutatusThe Wilson Journal of Ornithology, vol. 123, no. 4, p. 726-733.).
The region experiences extreme variations in rainfall, with particularly wet summers (from November to January) and extremely dry winters (from June to September). Usually, there is a soil water deficit from May to August, which coincides with the coldest months of the year (hereafter referred to as the dry season). In the rainy season, which lasts from November to March, there is excess water in the soil coinciding with the warmest months of the year (Rodrigues et al., 2011RODRIGUES, M., Freitas, GHS., Costa, LM., Dias, DF., Varela, MLM. and Rodrigues, LC., 2011. Avifauna of Alto do Palácio, Serra do Cipó National Park, state of Minas Gerais, southeastern Brazil. Check List, vol. 7, p. 151-161.).
2.2 Data collection
Data were collected monthly from August 2007 to July 2009 (except in December 2008, due to constant rain) in campaigns that lasted four to six days each.
To perform monthly assessments of the hummingbird species composition and richness in an open rocky field and two forest patches, 10 mist nets (12 x 2.6 m, 25-mm mesh) were mounted during two sampling events (one morning for each habitat).
The mist nets were mounted at sunrise (0600 h EST), remained open for six consecutive hours
and were checked every 30 min. The captured hummingbirds were identified and banded. To record
the species that were not captured in the mist nets and to record the hummingbirds’ visits to
flowers, observations were performed during the early morning along transects of 1800 m of
trails with 1200 m in the open grassland and 600 m in the open rocky fields. Each 5-min
observation was performed at 100-m intervals. In the forest patches walks were performed
monthly in the surroundings of two forest patches (total areas of 230 m2 and 170
m2) with one 5-min observation point in each area. All of the hummingbirds spotted
during the walk and at the observation points were recorded (modified from Feinsinger, 1976Feinsinger, P., 1976. Organization of a tropical guild of nectarivorous birds.
Ecological Monographs, vol. 46, no. 3, p. 257-291.
http://dx.doi.org/10.2307/1942255.
http://dx.doi.org/10.2307/1942255...
).
The hummingbirds were categorised as resident if they were registered throughout the year
(even when they were not registered for up to two consecutive months) and as non-resident if
they were absent for three or more consecutive months (modified from Machado, 2009Machado, CG., 2009. Beija-flores (Aves: Trochilidae) e seus recursos florais em
uma área de caatinga da Chapada Diamantina, Bahia, Brasil. Zoologia, vol. 26, no. 2, p.
255-265. http://dx.doi.org/10.1590/S1984-46702009000200008.
http://dx.doi.org/10.1590/S1984-46702009...
).
The plant species visited by the hummingbirds were recorded by direct observation or with the aid of binoculars while walking along the transects. In addition, we conducted monthly focal observations (with minimal and maximum durations of 40 min and 180 min, respectively) on species selected based on the following characteristics: 1) flowering species used by hummingbirds or 2) species that produced nectar and whose morphology allowed visitation by these birds (Rodrigues and Araujo, 2011RODRIGUES, LC. and Araujo, AC., 2011. The hummingbird community and their floral resources in an urban forest remnant in Brazil. Revista Brasileira de Biologia = Brazilian Journal of Biology, vol. 71, no. 3, p. 611-622. PMid:21881784.). We prioritised flowering plants that were more easily accessed, had a greater abundance of flowers and were close to other flowering plants that could also be visited (Rocca-de-Andrade, 2006Rocca-De-Andrade, M., 2006. Recurso floral para aves em uma comunidade de Mata Atlântica de encosta: sazonalidade e distribuição vertical. Campinas: Universidade Estadual de Campinas. Tese de Doutorado em Biologia Vegetal.). The observations were done in different hours of anthesis, mainly between 0600 h and 1100 h and between 1500 h and 1800 h.
The quantity of floral resources available to hummingbirds was recorded monthly at the same trails where the sightings were performed. The number of flowers on all flowering individuals along the trails and in a 10-m wide strip in the open rocky field (resulting in a total of 6000 m2) was recorded. In the two forest patches, sampling was performed using systematic walks at the edge and in the interior to sample the entire area of this environment (400 m2) (Rodrigues and Rodrigues, 2014RODRIGUES, LC. and Rodrigues, M., 2014. Flowers visited by hummingbirds in the open habitats of the southeastern Brazilian moutaintops: species composition and seasonality. Revista Brasileira de Biologia = Brazilian Journal of Biology, vol. 74, no. 3, p. 659-676. PMID: 252962.).
To quantify the volume and concentration of nectar, we bagged flower buds and flowers (in the case of flowers that last more than one day) and measured between 1000 and 1300 h of the following day. The nectar volume was measured using a microsyringe, and the concentration of solutes was measured with a manual refractometer (Atago, 0-32%; Galetto and Bernardello, 2005Galetto, L. and Bernardello, G., 2005. Rewards in flowers: Nectar. In DAFNI, A., Kevan, PG. and Husband, BC. (Eds.). Practical pollination biology. Cambridge: Enviroquest Ltd. 590 p.). The ideal sampling size consisted of 20 flowers of at least three different individuals; however, limitations intrinsic to each species (such as the density of individuals in the study area and/or plants that only produce a single flower every two or three days) resulted in varied sample sizes. To estimate the energetic value of the nectar, the nectar volume and concentration measurements were converted to mg of total sugar according to Galetto and Bernardello (2005)Galetto, L. and Bernardello, G., 2005. Rewards in flowers: Nectar. In DAFNI, A., Kevan, PG. and Husband, BC. (Eds.). Practical pollination biology. Cambridge: Enviroquest Ltd. 590 p.. The data of nectar volume, concentration and mg of total sugar were present in Rodrigues and Rodrigues (2014)RODRIGUES, LC. and Rodrigues, M., 2014. Flowers visited by hummingbirds in the open habitats of the southeastern Brazilian moutaintops: species composition and seasonality. Revista Brasileira de Biologia = Brazilian Journal of Biology, vol. 74, no. 3, p. 659-676. PMID: 252962..
The resource density (RD) of ornithophilous and non-ornithophilous resources available each month for the hummingbirds in each of the sample areas was calculated by the following Equation 1:
where F = total number of open flowers per day; N = average total quantity of mg of sugar in the nectar; x = visited plant; and A = size of the sampled area (m2) in each of the studied habitats.
The in situ total precipitation was measured monthly using a pluviometer located near the sampled areas (Rodrigues and Rodrigues 2014RODRIGUES, LC. and Rodrigues, M., 2014. Flowers visited by hummingbirds in the open habitats of the southeastern Brazilian moutaintops: species composition and seasonality. Revista Brasileira de Biologia = Brazilian Journal of Biology, vol. 74, no. 3, p. 659-676. PMID: 252962.).
2.3 Data analysis
Hummingbirds with evident sexual dimorphism were treated separately in the analyses (Rodrigues and Araujo, 2011RODRIGUES, LC. and Araujo, AC., 2011. The hummingbird community and their floral resources in an urban forest remnant in Brazil. Revista Brasileira de Biologia = Brazilian Journal of Biology, vol. 71, no. 3, p. 611-622. PMid:21881784.; Rodrigues and Rodrigues, 2011RODRIGUES, LC. and Rodrigues, M., 2011. Size dimorphism, juvenal plumage, and timing of breeding of the hyacinth Visorbearer (). Augastes scutatusThe Wilson Journal of Ornithology, vol. 123, no. 4, p. 726-733.). Months in which fewer than three hummingbirds were recorded and hummingbird species that occurred in less than three months in the two habitats were excluded from the ordination and from the non-metric multidimensional scaling (NMDS) analysis.
The quantity of floral resources available for the hummingbirds was represented by the proportion of ornithophilous resources (which is inversely proportional to that of non-ornithophilous resources) calculated monthly for each of the environments based on the resource density. Pearson correlation analyses were performed to verify whether there was a relationship between the ornithophilous and non-ornithophilous resource density and precipitation.
A representative gradient of the hummingbird community structure based on species composition
was obtained by sample ordination. These ordinations were obtained from matrices on the
relative abundance of species (Ferreira, 1997Ferreira, AM., 1997. Transformações de dados em medidas de semelhança e suas
interpretações ecológicas. Revista Brasileira de Ecologia, vol. 1, p. 111-114.). The
Bray-Curtis dissimilarity index (Bray and Curtis, 1957Bray, JR. and Curtis, JT., 1957. An ordination of the upland forest communities
of southern Wisconsin. Ecological Monographs, vol. 27, no. 4, p. 325-349.
http://dx.doi.org/10.2307/1942268.
http://dx.doi.org/10.2307/1942268...
)
was used to obtain the association matrices that were subsequently subjected to NMDS. The
proportion of explained variance (“R2”) of the values of the
original distances in the association matrix relative to those derived from the ordination in
one, two and three dimensions was used to decide the number of dimensions of the ordinations
(e.g.,
Azevedo-Ramos et al., 1999Azevedo-Ramos, C., Magnusson, WE. and Bayliss, P., 1999. Predation as the key
factor structuring tadpole assemblages in a savanna area in central Amazonia. Copeia, vol.
1999, no. 1, p. 22-33. http://dx.doi.org/10.2307/1447381.
http://dx.doi.org/10.2307/1447381...
). Based on
the comparison of the R2 values, an ordination with three
dimensions was selected.
The effect of the variables “habitat”, “quantity of available floral resource”, “precipitation” and “sampling period (month)” on the hummingbird species composition in the sampled environments was tested by multivariate analysis of covariance (Pillai trace statistic). The analyses were performed using R 2.9.1 software (R Development Core Team, 2009R Development Core Team, 2009. R: A language and environment for statistical computing. Vienna: R Foundation for Statistical Computing. Available from: <http://www.R-project.org>.).
3 Results
We recorded 12 hummingbird species during the study period. With the exception of Heliomaster squamosus (Temminck, 1823), male Thalurania glaucopis (Gmelin, 1788) and female Thalurania sp., which were found only in the forests patches, all other species recorded in this study occurred in both of the studied habitats (Chart 1).
Hummingbird species recorded throughout the first study year (August 2007 to July 2008, dotted lines) and the second year (August 2008 to July 2009, continuous lines) in the forest patches and the open rocky field at Alto do Palácio, Serra do Cipó National Park.
In the two sampled habitats, 195 individuals of hummingbirds belonging to 11 species were captured and marked. We captured 114 individuals in the open rocky fields and 81 individuals in the forest patches. Augastes scutatus (Temminck, 1824), was the species with the largest number of captures (N = 107; males = 66, females = 33 and 3 individuals of indeterminate sex), being 76 individuals captured in the open rocky fields (males = 46 and females = 27) and 31 in the forest patches (males = 20 and females = 11). Amazilia versicolor (Vieillot, 1818) and Leucochloris albicollis (Vieillot, 1818) were the species with the smallest number (N = 2 for each species) of captures (Figure 1). With the exception of male and female of A. scutatus (Figure 2A and B) and Campylopterus largipennis (Boddaert, 1783) (Figure 2C), which were more frequently captured in the open rocky fields, the other hummingbirds were more frequently observed in the forest patches or had similar frequencies between the sampled habitats (Figure 1). Three hummingbird species were recorded throughout approximately the entire study period and considered as resident, namely, A. scutatus, Colibri serrirostris (Vieillot, 1816) (Figure 2D) and Phaethornis pretrei (Lesson & Delattre, 1839) (Chart 1 and Figure 2F).

Total number (black bars) of captured hummingbirds per species and the number of hummingbirds captured in the open rocky field (grey bars) and forest patches (empty bars) at Alto do Palácio, Serra do Cipó National Park.

Hummingbird species recorded at Alto do Palácio, Serra do Cipó National Park, MG. A, male Augastes scutatus (photo: Guilherme Freitas); B, female Augastes scutatus; C, Campylopterus largipennis; D, Colibri serrirostris; E, male Chlorostilbon lucidus and F, Phaethornis pretrei visiting Lobelia fistulosa, an ornithophilous species with a long corolla.
The variation in the hummingbird species composition in both habitats was represented by the NMDS ordination in three dimensions (R2 = 0.89). The hummingbird community differed between the sampled habitats (p < 0.01, Table 1), mainly due to the relative abundance of males of A. scutatus, which were negatively correlated with the first dimension of the NMDS ordination (r = –0.96, Table 2). Males of Augastes scutatus were more abundant in the open rocky fields than in the forest patches throughout the study period (Figure 3). Phaethornis pretrei was also important in separating the open rocky fields and forest patches samples (Table 2); in addition, males of A. scutatus were abundant, whereas P. pretrei males were not. A similar pattern was found for females of A. scutatus and C. serrirostris (Figure 3); however, this pattern was related to the second dimension, which did not clearly distinguish between the habitats (Table 2). The remaining hummingbirds had little contribution (r < 0.5, Table 2) to the separation of the samples between the habitats.
Results of the multivariate covariance analysis (df = 3 and 31) used to explain the variation in the hummingbird species composition during 22 sample months in a typical rocky field and in forest patches, at Alto do Palácio, Serra do Cipó National Park. The species composition was represented by sample ordination in three dimensions using non-metric multidimensional scaling.
Correlations between the relative abundances of hummingbird species (separated by sex for species with evident sexual dimorphism) and the three dimensions of the sample ordination by non-metric multidimensional scaling. Absolute values greater than 0.5 are shown in bold.

Number of captured individuals per month in the open rocky field (full bars) and in the forest patches (empty bars) for the most abundant species and that influenced the separation of the samples between the phytophysiognomies according to the NMDS.
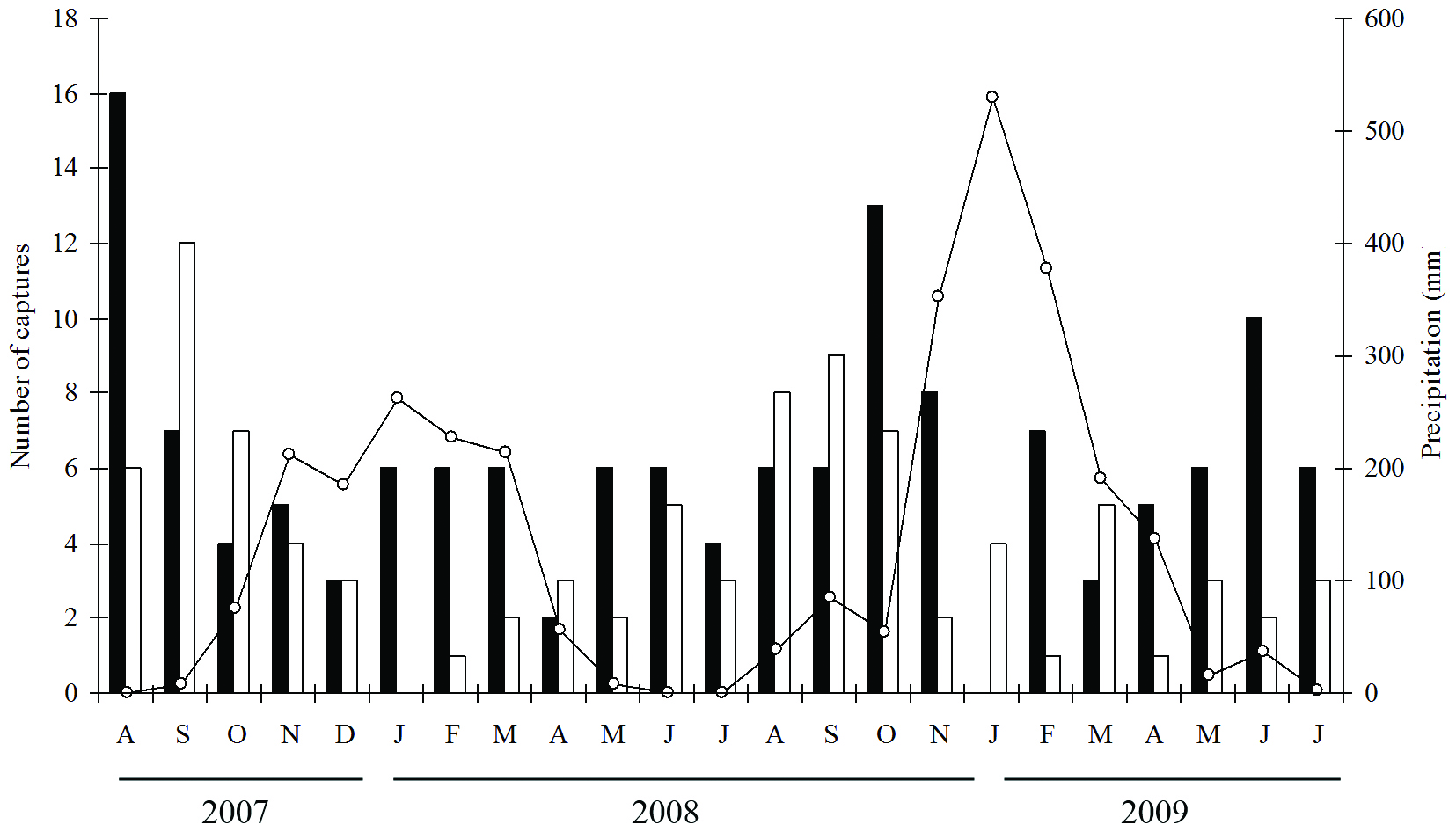
For both habitats, the greatest number of hummingbird species (N = 10) was recorded in October 2008 (Chart 1), and the greatest number of captures occurred in August (N = 22) and September (N = 19) 2007 and October 2008 (N = 20), periods with a low precipitation (Figure 4). However, the hummingbird species composition did not vary significantly in correlation with either the sampling period or precipitation (Table 1).

Precipitation (line) and number of hummingbird individuals captured throughout the study period in the open rocky field (black bars) and in the forest patches (empty bars) at Alto do Palácio, Serra do Cipó National Park.
Hummingbird visited 51 plant taxa when subspecies and varieties were considered separately, with 43% (N = 22) and 57% (N = 29) of these being ornithophilous and non-ornithophilous, respectively (see Rodrigues and Rodrigues, 2014RODRIGUES, LC. and Rodrigues, M., 2014. Flowers visited by hummingbirds in the open habitats of the southeastern Brazilian moutaintops: species composition and seasonality. Revista Brasileira de Biologia = Brazilian Journal of Biology, vol. 74, no. 3, p. 659-676. PMID: 252962.). The open rocky fields area presented a greater number of visited species (N = 38) when compared to the forest patches (N = 17). The largest portion of these species were non-ornithophilous (open rocky fields = 58%, forest patches = 53%).
Male and female A. scutatus visited the largest number of plant species (N = 29 and N = 20, respectively), especially the non-ornithophilous species (males = 76% and females = 60%), whereas P. pretrei visited the largest proportion of ornithophilous species (82%).
The density of ornithophilous resources was not correlated with precipitation (p = 0.1 in the open rocky fields and p = 0.09 in the forest patches), and the same observation was made for the density of non-ornithophilous resources (p = 0.78 for open rocky fields, p = 0.21 for forest patches) (Table 3). However, the proportion of ornithophilous resources explained a good proportion of the variation in the hummingbird species composition (Table 1); i.e., variations in the proportion of ornithophilous resources correspond to variations in the hummingbird species that occur in each sample.
Precipitation (mm) and energy density (mg of sugar/m2) of ornithophilous and non-ornithophilous resources throughout the study period in the open rocky field (ORF) and in the forest patches (FP) at Alto do Palácio, Serra do Cipó National Park.
4 Discussion
This is the first study to show that at a local scale, hummingbird species composition (species that comprise the samples and species relative abundance) is influenced by the type of habitat and the availability of floral resources but not the seasonality (precipitation and sampling period). The hummingbird community of the open rocky fields differs from that of the forest patches, mainly due to the relative abundance of the most common hummingbirds (males and females of A. scutatus, C. serrirostris and P. pretrei) in a given habitat of the Alto do Palácio region. This finding appears to reflect the habitat preferences of these species.
4.1 Species richness and composition
The recorded hummingbird species richness, as well as the number of resident hummingbird
species, was higher than those observed in previous studies performed for at least one year
with monthly sampling in natural forest patches in the Pantanal (Araujo and Sazima, 2003Araujo, AC. and Sazima, M., 2003. The assemblage of flowers visited by
hummingbirds in the capões of southern Pantanal, Mato Grosso do Sul, Brazil. Flora, vol. 198,
no. 6, p. 427-435. http://dx.doi.org/10.1078/0367-2530-00116.
http://dx.doi.org/10.1078/0367-2530-0011...
), in a cerrado area (Rodrigues and Araujo, 2011RODRIGUES, LC. and Araujo, AC., 2011. The hummingbird community and their floral
resources in an urban forest remnant in Brazil. Revista Brasileira de Biologia = Brazilian
Journal of Biology, vol. 71, no. 3, p. 611-622. PMid:21881784.) and in open rocky fields areas (Vasconcelos and Lombardi, 1999Vasconcelos, MF. and Lombardi, JA., 1999. Padrão Sazonal de ocorrência de seis
espécies de beija-flores (Apodiformes: Trochilidae) em uma localidade de campo rupestre na
Serra do Curral, Minas Gerais. Ararajuba, vol. 7, p. 71-79.; Machado et al., 2007Machado, CG., Coelho, AG., Santana, CS. and Rodrigues, M., 2007. Beija-flores e
seus recursos florais em uma área de campo rupestre da Chapada Diamantina, Bahia. Revista
Brasileira de Ornitologia, vol. 15, p. 267-279.). However, it was similar to the
registered in certain Atlantic Forest areas (Araujo,
1996Araujo, AC., 1996. Beija-flores e seus recursos florais numa área de planície
costeira do litoral norte de São Paulo. Campinas: Universidade Estadual de Campinas.
Dissertação de Mestrado.; Rocca-de-Andrade, 2006Rocca-De-Andrade, M., 2006. Recurso floral para aves em uma comunidade de Mata
Atlântica de encosta: sazonalidade e distribuição vertical. Campinas: Universidade Estadual de
Campinas. Tese de Doutorado em Biologia Vegetal.) and in the
Colombian Amazon (Rosero-Lasprilla and Sazima,
2004Rosero-Lasprilla, L. and Sazima, M., 2004. Interacciones planta-colibrí em tres
comunidades vegetales de la parte suroriental Del Parque Nacinal Natural Chiribiquete,
Colombia. Ornitologia Neotropical, vol. 15, p. 183-190.).
Although the number of Phaethornitinae species was similar to values recorded in certain
Atlantic Forest areas (Araujo, 1996Araujo, AC., 1996. Beija-flores e seus recursos florais numa área de planície
costeira do litoral norte de São Paulo. Campinas: Universidade Estadual de Campinas.
Dissertação de Mestrado.; Rocca-de-Andrade, 2006Rocca-De-Andrade, M., 2006. Recurso floral para aves em uma comunidade de Mata
Atlântica de encosta: sazonalidade e distribuição vertical. Campinas: Universidade Estadual de
Campinas. Tese de Doutorado em Biologia Vegetal.), in the Colombian Amazon (Rosero-Lasprilla and Sazima, 2004Rosero-Lasprilla, L. and Sazima, M., 2004. Interacciones planta-colibrí em tres
comunidades vegetales de la parte suroriental Del Parque Nacinal Natural Chiribiquete,
Colombia. Ornitologia Neotropical, vol. 15, p. 183-190.) and in natural forest
patches in the southern Pantanal (Araujo and Sazima,
2003Araujo, AC. and Sazima, M., 2003. The assemblage of flowers visited by
hummingbirds in the capões of southern Pantanal, Mato Grosso do Sul, Brazil. Flora, vol. 198,
no. 6, p. 427-435. http://dx.doi.org/10.1078/0367-2530-00116.
http://dx.doi.org/10.1078/0367-2530-0011...
), the hummingbird P. eurynome was recorded only in larger forest
patches. Therefore, the presence of this hummingbird is associated to the presence of forested
areas at Alto do Palácio, a typical environment for Phaethornitinae species (Stiles, 1981Stiles, FG., 1981. Geographical aspects of bird-flower coevolution, with
particular reference to Central America. Annals of the Missouri Botanical Garden, vol. 68, no.
2, p. 323-351. http://dx.doi.org/10.2307/2398801.
http://dx.doi.org/10.2307/2398801...
).
Augastes scutatus, the most abundant hummingbird and the one with the
highest relative abundance in the open rocky fields (both males and females), is a species
endemic to the Espinhaço Range. Its distribution is restricted to the central-southern portion
of the range, and it occurs only at altitudes greater than 1000 m (Vasconcelos and Rodrigues, 2010Vasconcelos, MF. and Rodrigues, M., 2010. Patterns of geographic distribution
and conservation of the open-habitat avifauna of southeastern Brazilian mountaintops ( and
campos de altitude). campos rupestresPapéis Avulsos de
Zoologia, vol. 50, p. 1-29.). Augastes scutatus appears
to be more dependent on plant species that occur in open rocky fields and becomes locally
extinct in degraded areas, possibly because of the extinction of certain plant species (Vasconcelos and Lombardi, 1999Vasconcelos, MF. and Lombardi, JA., 1999. Padrão Sazonal de ocorrência de seis
espécies de beija-flores (Apodiformes: Trochilidae) em uma localidade de campo rupestre na
Serra do Curral, Minas Gerais. Ararajuba, vol. 7, p. 71-79.). This hypothesis is
supported by the present study, which recorded visits made by this hummingbird to 23 plant
species that occur in the open rocky fields and only 11 species that occur in forest patches
(see Rodrigues and Rodrigues, 2014RODRIGUES, LC. and Rodrigues, M., 2014. Flowers visited by hummingbirds in the
open habitats of the southeastern Brazilian moutaintops: species composition and seasonality.
Revista Brasileira de Biologia = Brazilian Journal of Biology, vol. 74, no. 3, p. 659-676.
PMID: 252962.). In addition,
A. scutatus also appears to prefer open rocky fields as a nesting site (Rodrigues and Rodrigues, 2011RODRIGUES, LC. and Rodrigues, M., 2011. Size dimorphism, juvenal plumage, and
timing of breeding of the hyacinth Visorbearer (). Augastes scutatusThe Wilson
Journal of Ornithology, vol. 123, no. 4, p. 726-733.). Therefore, this species may
be considered highly adapted to open rocky fields, depending on its floral and nesting
resources (Rodrigues et al., 2013RODRIGUES, LC., Martins, FI. and Rodrigues, M. 2013 Survival of a mountaintop
hummingbird, the Hyacinth Visorbearer Augastes scutatus, in Southeastern Brazil. Acta
Ornithologica, vol. 48, no. 2, p. 211-219.
http://dx.doi.org/10.3161/000164513X678801
http://dx.doi.org/10.3161/000164513X6788...
).
Phaethornis pretrei was usually more abundant in the forest patches areas
throughout the study period. The Phaethornitinae are generally more frequently observed in the
understory of closed areas, such as riparian forests (Stiles,
1981Stiles, FG., 1981. Geographical aspects of bird-flower coevolution, with
particular reference to Central America. Annals of the Missouri Botanical Garden, vol. 68, no.
2, p. 323-351. http://dx.doi.org/10.2307/2398801.
http://dx.doi.org/10.2307/2398801...
; Cotton, 1998Cotton, PA., 1998. The hummingbird community of a lowland Amazonian rainforest.
The Ibis, vol. 140, no. 3, p. 512-521.
http://dx.doi.org/10.1111/j.1474-919X.1998.tb04613.x.
http://dx.doi.org/10.1111/j.1474-919X.19...
). In the Alto do Palácio
region, P. pretrei moves among the forest patches. Colibri
serrirostris was also more abundant in the forest patches, which appears to be
related to the territorial behaviour of this species. For nearly the entire study period
(August and September 2007, January to August 2008 and January to July 2009), one vocalising
C. serrirostris individual was observed on a perch in each of the sampled
forest patch. These individuals left these areas only for short periods (5 to 30 min), most
likely to forage in adjacent areas.
4.2 Floral resource availability
The hummingbird species compositions in the studied habitats varied due to the availability
of floral resources, which is most likely related to the hummingbirds’ ability to learn to
locate areas with greater resource availability (Malizia,
2001Malizia, LC., 2001. Seasonal fluctuations of birds, fruits, and flowers in a
subtropical forest of Argentina. The Condor, vol. 103, no. 1, p. 45-61.
http://dx.doi.org/10.1650/0010-5422(2001)103[0045:SFOBFA]2.0.CO;2.
http://dx.doi.org/10.1650/0010-5422(2001...
; Altshuler and Nunn, 2001Altshuler, DL. and Nunn, AM., 2001. Observational Learning in Hummingbirds. The
Auk, vol. 118, no. 3, p. 795-799.
http://dx.doi.org/10.1642/0004-8038(2001)118[0795:OLIH]2.0.CO;2.
http://dx.doi.org/10.1642/0004-8038(2001...
; Cole et al., 1982Cole, S., Hainsworth, FR., Kamil, AC., Mercier, T. and Wolf, LL., 1982. Spatial
learning as an adaptation in hummingbirds. Science, vol. 217, no. 4560, p. 655-657.
http://dx.doi.org/10.1126/science.217.4560.655. PMid:17817537
http://dx.doi.org/10.1126/science.217.45...
). An experimental study by Cole et al. (1982)Cole, S., Hainsworth, FR., Kamil, AC., Mercier, T. and Wolf, LL., 1982. Spatial
learning as an adaptation in hummingbirds. Science, vol. 217, no. 4560, p. 655-657.
http://dx.doi.org/10.1126/science.217.4560.655. PMid:17817537
http://dx.doi.org/10.1126/science.217.45...
showed that the learning of resource
locations by hummingbirds is related to the spatial distribution of resources, which influences
the energy acquisition rate. Therefore, during periods of low energetic availability within a
habitat, individuals of these species may switch locally to another habitat or forage in nearby
areas with greater resource availability, as has been reported for other hummingbird
communities (Wolf et al., 1976Wolf, LL., Stiles, FG. and Hainsworth, FR., 1976. Ecological organization of a
tropical, highland hummingbird community. Journal of Animal Ecology, vol. 45, no. 2, p.
349-379. http://dx.doi.org/10.2307/3879.
http://dx.doi.org/10.2307/3879...
; Montgomerie and Gass, 1981Montgomerie, RD. and Gass, CL., 1981. Energy limitation of hummingbird
populations in tropical and temperate communities. Oecologia, vol. 50, no. 2, p. 162-165.
http://dx.doi.org/10.1007/BF00348031.
http://dx.doi.org/10.1007/BF00348031...
; Kodric-Brown et
al., 1984Kodric-BrowN, A., Brown, JH., Byers, GS. and Gori, DF., 1984. Organization of a
tropical island community of hummingbirds and flowers. Ecology, vol. 65, no. 5, p. 1358-1368.
http://dx.doi.org/10.2307/1939116.
http://dx.doi.org/10.2307/1939116...
; Stiles, 1985Stiles, FG., 1985. Seasonal pattern and coevolution in the hummingbird-flower
community of a Costa Rican sub-tropical forest. In BUCKLEY, PA., MORTON, MS., RIDGLEY, RS. and
BUCKLEY, FG. (Eds.). Neotropical ornithology. Washington: American Ornithologists Union. p.
757-787.). In addition, the
recorded hummingbird species used different floral resources, which reduces the competition
among species (Stiles, 1985Stiles, FG., 1985. Seasonal pattern and coevolution in the hummingbird-flower
community of a Costa Rican sub-tropical forest. In BUCKLEY, PA., MORTON, MS., RIDGLEY, RS. and
BUCKLEY, FG. (Eds.). Neotropical ornithology. Washington: American Ornithologists Union. p.
757-787.).
The hummingbirds with greater bill lengths used a larger proportion of ornithophilous resources in their diets and usually used flowers with greater corolla lengths. Conversely, hummingbirds with smaller bill lengths used a large proportions of non-ornithophilous species and generally used flowers with smaller corolla lengths (L.C. Rodrigues et al., unpublished data). Therefore, when the energetic availability of a given flower type is low within a habitat, the hummingbirds may search for the flower in another habitat or in nearby areas. In this way, hummingbird species composition varies with changes in the energy density of floral resources.
Certain studies have shown that the flowering peak of the species visited by hummingbirds is
concentrated at the end of the dry season and/or during the entire rainy season (Arizmendi and Ornelas, 1990Arizmendi, MC. and Ornelas, JF., 1990. Hummingbirds and their floral resources
in a tropical dry forest in Mexico. Biotropica, vol. 22, no. 2, p. 172-180.
http://dx.doi.org/10.2307/2388410.
http://dx.doi.org/10.2307/2388410...
; Sazima et al., 1996Sazima, I., Buzato, S. and Sazima, M., 1996. An assemblage of
hummingbird-pollinated flowers in a montane forest in southeastern Brazil. Botanica Acta, vol.
109, no. 2, p. 149-160. http://dx.doi.org/10.1111/j.1438-8677.1996.tb00555.x.
http://dx.doi.org/10.1111/j.1438-8677.19...
; Araujo and Sazima,
2003Araujo, AC. and Sazima, M., 2003. The assemblage of flowers visited by
hummingbirds in the capões of southern Pantanal, Mato Grosso do Sul, Brazil. Flora, vol. 198,
no. 6, p. 427-435. http://dx.doi.org/10.1078/0367-2530-00116.
http://dx.doi.org/10.1078/0367-2530-0011...
). However, in this study, no correlation was found between the energy density of
floral resources and precipitation. The constant availability of energy resources for
hummingbirds most likely favours the residence of the three hummingbirds species that were the
most abundant in the study area. In addition, the residence of these species may be one of the
factors explaining the absence of a relationship of hummingbird species composition with
precipitation and sampling period.
Although pronounced seasonality occurred in the study area, the hummingbird species compositions did not correlate with this factor (precipitation and sampling period). However, the compositions varied with the type of habitat, mainly due to the habitat preferences of the species that form the community as well as differences in the availability of ornithophilous and non-ornithophilous resources in the studied phytophysiognomies. The hummingbird species compositions also varied with the availability of floral resources, which is related to the hummingbirds’ capacity to locate the floral resources, as well as their preferences for different flower types.
Acknowledgments
We are grateful to CNPq and Capes for a scholarship awarded to LCR. MR thanks CNPq (473428/2004-0 and 300731/2006-0) and FAPEMIG (PPM APQ-0434-5.03/07). M. L.M. Varela, D. F. Dias, F. C. Diniz and R. Rocha helped in fieldwork. The Chico Mendes Institute (ICMBio) and directors of the Serra do Cipó National Park for logistical support. A. C. Araujo, C. M. Jacobi, M. F. Vasconcelos and M. Araujo greatly improved the manuscript.
-
(With 4 figures)
References
- Abrahamczyk, S. and Kessler, M., 2010. Hummingbird diversity, food niche characters, and assemblage composition along a latitudinal precipitation gradient in the Bolivian lowlands. Journal für Ornithologie, vol. 151, no. 3, p. 615-625. http://dx.doi.org/10.1007/s10336-010-0496-x.
» http://dx.doi.org/10.1007/s10336-010-0496-x - Araujo, AC., 1996. Beija-flores e seus recursos florais numa área de planície costeira do litoral norte de São Paulo. Campinas: Universidade Estadual de Campinas. Dissertação de Mestrado.
- Araujo, AC. and Sazima, M., 2003. The assemblage of flowers visited by hummingbirds in the capões of southern Pantanal, Mato Grosso do Sul, Brazil. Flora, vol. 198, no. 6, p. 427-435. http://dx.doi.org/10.1078/0367-2530-00116.
» http://dx.doi.org/10.1078/0367-2530-00116 - Altshuler, DL. and Nunn, AM., 2001. Observational Learning in Hummingbirds. The Auk, vol. 118, no. 3, p. 795-799. http://dx.doi.org/10.1642/0004-8038(2001)118[0795:OLIH]2.0.CO;2.
» http://dx.doi.org/10.1642/0004-8038(2001)118[0795:OLIH]2.0.CO;2 - Arizmendi, MC. and Ornelas, JF., 1990. Hummingbirds and their floral resources in a tropical dry forest in Mexico. Biotropica, vol. 22, no. 2, p. 172-180. http://dx.doi.org/10.2307/2388410.
» http://dx.doi.org/10.2307/2388410 - Azevedo-Ramos, C., Magnusson, WE. and Bayliss, P., 1999. Predation as the key factor structuring tadpole assemblages in a savanna area in central Amazonia. Copeia, vol. 1999, no. 1, p. 22-33. http://dx.doi.org/10.2307/1447381.
» http://dx.doi.org/10.2307/1447381 - Bray, JR. and Curtis, JT., 1957. An ordination of the upland forest communities of southern Wisconsin. Ecological Monographs, vol. 27, no. 4, p. 325-349. http://dx.doi.org/10.2307/1942268.
» http://dx.doi.org/10.2307/1942268 - Cole, S., Hainsworth, FR., Kamil, AC., Mercier, T. and Wolf, LL., 1982. Spatial learning as an adaptation in hummingbirds. Science, vol. 217, no. 4560, p. 655-657. http://dx.doi.org/10.1126/science.217.4560.655. PMid:17817537
» http://dx.doi.org/10.1126/science.217.4560.655 - Cotton, PA., 1998. The hummingbird community of a lowland Amazonian rainforest. The Ibis, vol. 140, no. 3, p. 512-521. http://dx.doi.org/10.1111/j.1474-919X.1998.tb04613.x.
» http://dx.doi.org/10.1111/j.1474-919X.1998.tb04613.x - Cotton, PA., 2007. Seasonal resource tracking by Amazonian hummingbirds. The Ibis, vol. 149, no. 1, p. 135-142. http://dx.doi.org/10.1111/j.1474-919X.2006.00619.x.
» http://dx.doi.org/10.1111/j.1474-919X.2006.00619.x - Des Granges, JL., 1979. Organization of a tropical nectar feeding bird guild in a variable environment. Living Bird, vol. 17, p. 199-236.
- Diamond, JM., Karasov, WH., Phan, D. and Carpenter, FL., 1986. Digestive physiology is a determinant of foraging bout frequency in hummingbirds. Nature, vol. 320, no. 6057, p. 62-63. http://dx.doi.org/10.1038/320062a0. PMid:3951548
» http://dx.doi.org/10.1038/320062a0 - Feinsinger, P., 1976. Organization of a tropical guild of nectarivorous birds. Ecological Monographs, vol. 46, no. 3, p. 257-291. http://dx.doi.org/10.2307/1942255.
» http://dx.doi.org/10.2307/1942255 - Ferreira, AM., 1997. Transformações de dados em medidas de semelhança e suas interpretações ecológicas. Revista Brasileira de Ecologia, vol. 1, p. 111-114.
- Galetto, L. and Bernardello, G., 2005. Rewards in flowers: Nectar. In DAFNI, A., Kevan, PG. and Husband, BC. (Eds.). Practical pollination biology. Cambridge: Enviroquest Ltd. 590 p.
- Hadley, AS. and Betts, MG., 2009. Tropical deforestation alters hummingbird movement patterns. Biology Letters, vol. 5, no. 2, p. 207-210. http://dx.doi.org/10.1098/rsbl.2008.0691. PMid:19158031
» http://dx.doi.org/10.1098/rsbl.2008.0691 - Kodric-BrowN, A., Brown, JH., Byers, GS. and Gori, DF., 1984. Organization of a tropical island community of hummingbirds and flowers. Ecology, vol. 65, no. 5, p. 1358-1368. http://dx.doi.org/10.2307/1939116.
» http://dx.doi.org/10.2307/1939116 - Levey, DJ., 1988. Spatial and temporal variation in Costa Rican fruit and fruit-eating bird abundance. Ecological Monographs, vol. 58, no. 4, p. 251-269. http://dx.doi.org/10.2307/1942539.
» http://dx.doi.org/10.2307/1942539 - Loiselle, BA. and Blake, JG., 1991. Temporal variation in birds and fruits along an elevational gradient in Costa Rica. Ecology, vol. 72, no. 1, p. 180-193. http://dx.doi.org/10.2307/1938913.
» http://dx.doi.org/10.2307/1938913 - Machado, CG., Coelho, AG., Santana, CS. and Rodrigues, M., 2007. Beija-flores e seus recursos florais em uma área de campo rupestre da Chapada Diamantina, Bahia. Revista Brasileira de Ornitologia, vol. 15, p. 267-279.
- Machado, CG., 2009. Beija-flores (Aves: Trochilidae) e seus recursos florais em uma área de caatinga da Chapada Diamantina, Bahia, Brasil. Zoologia, vol. 26, no. 2, p. 255-265. http://dx.doi.org/10.1590/S1984-46702009000200008.
» http://dx.doi.org/10.1590/S1984-46702009000200008 - Malizia, LC., 2001. Seasonal fluctuations of birds, fruits, and flowers in a subtropical forest of Argentina. The Condor, vol. 103, no. 1, p. 45-61. http://dx.doi.org/10.1650/0010-5422(2001)103[0045:SFOBFA]2.0.CO;2.
» http://dx.doi.org/10.1650/0010-5422(2001)103[0045:SFOBFA]2.0.CO;2 - Montgomerie, RD. and Gass, CL., 1981. Energy limitation of hummingbird populations in tropical and temperate communities. Oecologia, vol. 50, no. 2, p. 162-165. http://dx.doi.org/10.1007/BF00348031.
» http://dx.doi.org/10.1007/BF00348031 - Poulin, B., Lefebvre, G. and Mcneil, R., 1993. Variations in bird abundance in tropical arid and semiarid habitats. The Ibis, vol. 135, no. 4, p. 432-441. http://dx.doi.org/10.1111/j.1474-919X.1993.tb02116.x.
» http://dx.doi.org/10.1111/j.1474-919X.1993.tb02116.x - R Development Core Team, 2009. R: A language and environment for statistical computing. Vienna: R Foundation for Statistical Computing. Available from: <http://www.R-project.org>.
- Rocca-De-Andrade, M., 2006. Recurso floral para aves em uma comunidade de Mata Atlântica de encosta: sazonalidade e distribuição vertical. Campinas: Universidade Estadual de Campinas. Tese de Doutorado em Biologia Vegetal.
- RODRIGUES, LC. and Araujo, AC., 2011. The hummingbird community and their floral resources in an urban forest remnant in Brazil. Revista Brasileira de Biologia = Brazilian Journal of Biology, vol. 71, no. 3, p. 611-622. PMid:21881784.
- RODRIGUES, M., Freitas, GHS., Costa, LM., Dias, DF., Varela, MLM. and Rodrigues, LC., 2011. Avifauna of Alto do Palácio, Serra do Cipó National Park, state of Minas Gerais, southeastern Brazil. Check List, vol. 7, p. 151-161.
- RODRIGUES, LC. and Rodrigues, M., 2011. Size dimorphism, juvenal plumage, and timing of breeding of the hyacinth Visorbearer (). Augastes scutatusThe Wilson Journal of Ornithology, vol. 123, no. 4, p. 726-733.
- RODRIGUES, LC., Martins, FI. and Rodrigues, M. 2013 Survival of a mountaintop hummingbird, the Hyacinth Visorbearer Augastes scutatus, in Southeastern Brazil. Acta Ornithologica, vol. 48, no. 2, p. 211-219. http://dx.doi.org/10.3161/000164513X678801
» http://dx.doi.org/10.3161/000164513X678801 - RODRIGUES, LC. and Rodrigues, M., 2014. Flowers visited by hummingbirds in the open habitats of the southeastern Brazilian moutaintops: species composition and seasonality. Revista Brasileira de Biologia = Brazilian Journal of Biology, vol. 74, no. 3, p. 659-676. PMID: 252962.
- Rosero-Lasprilla, L. and Sazima, M., 2004. Interacciones planta-colibrí em tres comunidades vegetales de la parte suroriental Del Parque Nacinal Natural Chiribiquete, Colombia. Ornitologia Neotropical, vol. 15, p. 183-190.
- Sazima, I., Buzato, S. and Sazima, M., 1996. An assemblage of hummingbird-pollinated flowers in a montane forest in southeastern Brazil. Botanica Acta, vol. 109, no. 2, p. 149-160. http://dx.doi.org/10.1111/j.1438-8677.1996.tb00555.x.
» http://dx.doi.org/10.1111/j.1438-8677.1996.tb00555.x - Stiles, FG., 1981. Geographical aspects of bird-flower coevolution, with particular reference to Central America. Annals of the Missouri Botanical Garden, vol. 68, no. 2, p. 323-351. http://dx.doi.org/10.2307/2398801.
» http://dx.doi.org/10.2307/2398801 - Stiles, FG., 1985. Seasonal pattern and coevolution in the hummingbird-flower community of a Costa Rican sub-tropical forest. In BUCKLEY, PA., MORTON, MS., RIDGLEY, RS. and BUCKLEY, FG. (Eds.). Neotropical ornithology. Washington: American Ornithologists Union. p. 757-787.
- Vasconcelos, MF. and Lombardi, JA., 1999. Padrão Sazonal de ocorrência de seis espécies de beija-flores (Apodiformes: Trochilidae) em uma localidade de campo rupestre na Serra do Curral, Minas Gerais. Ararajuba, vol. 7, p. 71-79.
- Vasconcelos, MF. and Rodrigues, M., 2010. Patterns of geographic distribution and conservation of the open-habitat avifauna of southeastern Brazilian mountaintops ( and campos de altitude). campos rupestresPapéis Avulsos de Zoologia, vol. 50, p. 1-29.
- Wiens, JA., 1989. A case study: interactions among nectarivores. In BARNES, RSK., BIRKS, HJB., CONNOR, EF. and PAINE, RT. (Eds.). The ecology of bird communities. Cambridge: Cambridge University Press. 316 p. Processes and variations, vol. 2.
- Wolf, LL., Stiles, FG. and Hainsworth, FR., 1976. Ecological organization of a tropical, highland hummingbird community. Journal of Animal Ecology, vol. 45, no. 2, p. 349-379. http://dx.doi.org/10.2307/3879.
» http://dx.doi.org/10.2307/3879
Publication Dates
-
Publication in this collection
Jan-Mar 2015
History
-
Received
02 May 2013 -
Accepted
11 Oct 2013