
Abstract
The species of the genus Smilax, popularly known as sarsaparilla, are widely used in folk medicine due to the antirheumatic properties of its underground structures. Smilax fluminensis and S. syphilitica occur in forested areas and form thickened stems called rhizophores from which adventitious roots grow. To provide information for more accurate identification of the commercialised product and for elucidating the process of stem thickening, a morphology and anatomy study of the underground organs of the two species was conducted. The adventitious roots differ in colour and diameter depending on the stage of development. They are white and have a larger diameter in the early stages of development, but as they grow, the adventitious roots become brown and have a smaller diameter due to the disintegration of the epidermis and virtually the entire cortex. In brown roots, the covering function is then performed by the lignified endodermis and the remaining walls of the cells from the last parenchyma cortical layer. These results are similar to those found in studies of other Smilax and suggest that the anatomy of the roots can be useful for identifying fraud in commercialised materials. The thickening process of the nodal regions of the rhizophores in both species involves the activity of axillary buds and pericyclic layers.
Keywords:
sarsaparilla;
Smilax
; underground system; rhizophore; anatomy
Resumo
As espécies de Smilax, conhecidas popularmente como salsaparrilha, são amplamente utilizadas na medicina tradicional devido às propriedades antirreumáticas das estruturas subterrâneas. Smilax fluminensis e S. syphilitica ocorrem em áreas florestais e formam caules espessados denominados rizóforos a partir dos quais são emitidas raízes adventícias. Com o intuito de fornecer informações para a identificação mais precisa do material comercializado e no entendimento do processo de espessamento do caule, foi realizado o estudo morfológico e anatômico dos órgãos subterrâneos das duas espécies. As raízes adventícias apresentam diferenças na coloração e no diâmetro dependendo da fase de desenvolvimento. As raízes nas fases iniciais do desenvolvimento são brancas e possuem diâmetro maior, porém com o desenvolvimento, devido à desintegração da epiderme e de praticamente todo o córtex, as raízes tornam-se marrons e de diâmetro menor. Nas raízes marrons, a função de revestimento passa a ser exercida pela endoderme lignificada e pelas paredes remanescentes das células da penúltima camada cortical. Os resultados são semelhantes aos encontrados nos estudos de outras Smilax e sugerem que a anatomia das raízes pode ser útil na identificação de fraudes em materiais comercializados. O processo de espessamento das regiões nodais dos rizóforos nas duas espécies envolve a atividade das gemas axilares e de camadas pericíclicas.
Palavras-chave:
salsaparrilha;
Smilax
; sistema subterrâneo; rizóforo; anatomia
1 Introduction
Smilax comprises 310 species distributed in temperate and tropical regions (Judd et al., 2009Judd, WS., Campbell, CS., Kellogg, EA., STEVENS, PF. and DONOGHUE, MJ., 2009. Sistemática vegetal: um enfoque filogenético. Porto Alegre: Artmed.). There are 32 species in Brazil (Andreata, 1997Andreata, RHP., 1997. Revisão das espécies brasileiras do gênero Linnaeus (Smilacaceae). SmilaxPesquisa Botânica, vol. 47, p. 7-244.; Andreata, 2009Andreata, RHP., 2009. A new species of and a key to all species of Minas Gerais, Brazil. SmilaxSystematic Botany, vol. 34, no. 1, p. 28-31. http://dx.doi.org/10.1600/036364409787602302.
http://dx.doi.org/10.1600/03636440978760...
), some of which are popularly known as sarsaparillas and have been used in folk medicine since the 16th century (Medeiros et al., 2007Medeiros, MFT., Senna-Valle, L. and Andreata, RHP., 2007. Histórico e o uso da “salsaparrilha” ( spp.) pelos boticários no Mosteiro de São Bento. SmilaxRevista Brasileira de Biociências, vol. 5, p. 27-29.).
For many years, sarsaparilla was identified by how their roots were tied in bundles for commercialization and by their external appearance (Farmacopéia, 1929Farmacopéia, 1929. Codigo pharmaceutico brasileiro: pharmacopeia dos Estados Unidos do Brasil. Rio de Janeiro: Companhia Editora Nacional. 1149 p. Redigida por Rodolpho Albino Dias da Silva.). Studies of the Brazilian Smilax have demonstrated that their roots change in thickness and colour according to the stage of development (Martins and Appezzato-da-Glória, 2006Martins, AR. and Appezzato-Da-Gloria, B., 2006. Morfoanatomia dos órgãos vegetativos Smilax polyantha Griseb. (Smilacaceae). Brazilian Journal of Botany, vol. 29, no. 4, p. 555-567. http://dx.doi.org/10.1590/S0100-84042006000400005.
http://dx.doi.org/10.1590/S0100-84042006...
; Martins et al., 2010Martins, AR., Pütz, N., Soares, AN., Bombo, AB. and Appezzato-Da-Gloria, B., 2010. New approaches to underground system in Brazilian Smilax species (Smilacaceae). The Journal of the Torrey Botanical Society, Athens, vol. 137, no. 2-3, p. 220-235. http://dx.doi.org/10.3159/10-RA-024R.1.
http://dx.doi.org/10.3159/10-RA-024R.1...
; Guimarães et al., 2010Guimarães, A.R., Costa, C.G. and Andreata, R.H.P., 2010. Morfoanatomia do sistema subterrâneo de Smilax subsessiliflora (Smilacaceae). Rodriguésia, vol. 61, no. 2, p. 181-194.), i.e., these features are not useful for identifying the sarsaparilla. However, the anatomical features of the root in cross section, such as the relative thicknesses of cortical, vascular, and medullary regions; the presence of starch; the distribution of phenolic and crystalliferous idioblasts; and the phloem arrangement, can provide a larger number of elements for more accurate identification of the drug (Cunha, 1937Cunha, NS., 1937. Da salsaparrilha a japecanga. Tribuna Farmaceutica, vol. 5, p. 145-150.; Stellfeld, 1938Stellfeld, C., 1938. A salsaparilha do Paraná e a ausência do parênquima cortical. Tribuna Farmaceutica, vol. 6, p. 5-7.; Soares, 2013Soares, MKM., 2013. Anatomia e perfil químico da salsaparrilha comercializada no Estado de São Paulo. Piracicaba: Universidade de São Paulo. 69 p. Dissertação de Mestrado em Fisiologia e Bioquímica de Plantas.). The accurate identification of medicinal plants, including the detailed morphological and anatomical descriptions of its underground organs, is essential for quality control in phytotherapeutic studies (Ming, 1994Ming, LC., 1994. Estudos e pesquisas de plantas medicinais na agronomia. Horticultura Brasileira, vol. 12, p. 3-9.).
Smilax fluminensis and S. syphilitica occur in forested areas and form thickened stems called rhizophores, following the classification proposed by Andreata and Menezes (1999)Andreata, RHP. and MENEZES, NL., 1999. Morfoanatomia do embrião, desenvolvimento pós-seminal e origem do rizóforo de Smilax quinquenervia Vell. Boletim de Botânica da Universidade de São Paulo, vol. 18, p. 39-51. http://dx.doi.org/10.11606/issn.2316-9052.v18i0p39-51.
http://dx.doi.org/10.11606/issn.2316-905...
for S. quinquenervia. According to them, the rhizophores of this species, unlike the rhizomes, have two stem axes during the early stages of development. An aerial stem system of plumular origin and an underground stem system of cotyledonary bud origin have been described, and the root system is completely adventitious and formed by the underground stem axis (Andreata and Menezes, 1999Andreata, RHP. and MENEZES, NL., 1999. Morfoanatomia do embrião, desenvolvimento pós-seminal e origem do rizóforo de Smilax quinquenervia Vell. Boletim de Botânica da Universidade de São Paulo, vol. 18, p. 39-51. http://dx.doi.org/10.11606/issn.2316-9052.v18i0p39-51.
http://dx.doi.org/10.11606/issn.2316-905...
). The presence of two stem axes was also observed in Smilax polyantha, but the underground stem axis originated from axillary buds of the cataphylls at the base of the aerial branching (Martins et al., 2011Martins, AR., Pütz, N., Novembre, ADLC., Piedade, SMS. and Appezzato-Da-Glória, B., 2011. Seed germination and seedling morphology of Smilax polyantha (Smilacaceae). Biota Neotropica, vol. 11, no. 2, p. 1-7. http://dx.doi.org/10.1590/S1676-06032011000200003.
http://dx.doi.org/10.1590/S1676-06032011...
) and not from the cotyledonary bud as described in S. rotundifolia and S. glauca (Holm, 1890Holm, T., 1890. Contribution to the knowledge of the germination of some North American plants. Memoirs of the Torrey Botanical Club, vol. 2, p. 57-108.) and in S. quinquenervia (Andreata and Menezes, 1999Andreata, RHP. and MENEZES, NL., 1999. Morfoanatomia do embrião, desenvolvimento pós-seminal e origem do rizóforo de Smilax quinquenervia Vell. Boletim de Botânica da Universidade de São Paulo, vol. 18, p. 39-51. http://dx.doi.org/10.11606/issn.2316-9052.v18i0p39-51.
http://dx.doi.org/10.11606/issn.2316-905...
).
The rhizophores of Smilax exhibit nodal thickening from which all adventitious roots of the plant are emitted (Martins et al., 2011Martins, AR., Pütz, N., Novembre, ADLC., Piedade, SMS. and Appezzato-Da-Glória, B., 2011. Seed germination and seedling morphology of Smilax polyantha (Smilacaceae). Biota Neotropica, vol. 11, no. 2, p. 1-7. http://dx.doi.org/10.1590/S1676-06032011000200003.
http://dx.doi.org/10.1590/S1676-06032011...
). Moreover, the nodes of the stem branches, when stimulated by fragmentation, also thicken and form adventitious roots, thereby increasing the chances for vegetative propagation (Soares et al.,2011SOARES, AN., NOVEMBRE, ADLC., MARTINS, AR., PIEDADE, SMS. and APPEZZATO-DA-GLÓRIA, B., 2011. Propagation studies in Smilax fluminensis Steud. (Smilacaceae). Ciência Rural, vol. 41, no. 10, p. 1762-1768. http://dx.doi.org/10.1590/S0103-84782011001000014.
http://dx.doi.org/10.1590/S0103-84782011...
).
The primary thickening in monocotyledons has been discussed under a different approach by Menezes et al. (2005Menezes, NL., Silva, DC., Arruda, RCO., Melo-de-Pinna, GFA., Cardoso, VA., Castro, NM., Scatena, VL. and Scremin-Dias, E., 2005. Meristematic activity of the Endodermis and the Pericycle in the primary thickening in monocotyledons: considerations on the “PTM”. Anais da Academia Brasileira de Ciencias, vol. 77, no. 2, p. 259-274. http://dx.doi.org/10.1590/S0001-37652005000200006. PMid:15895162.
http://dx.doi.org/10.1590/S0001-37652005...
, 2012Menezes, NL., Elbl, PM., Cury, G., Sasaki, KLM., Appezzato-Da-Glória, B., Costa, GO. and Silva, CG., 2012. The meristematic activity of the endodermis and the pericycle and its role in the primary thickening of stems in monocotyledonous plants. Plant Ecology & Diversity, vol. 5, no. 2, p. 153-165. http://dx.doi.org/10.1080/17550874.2011.604925.
http://dx.doi.org/10.1080/17550874.2011....
). According to them, in some species, such as Zingiber officinale (Zingiberaceae), the observation of Casparian bands in the endodermis allowed researchers to verify that the pericycle is involved in the tuberisation process of the rhizome. However, in Smilax polyantha, it was not possible to establish the boundary between the cortex and the vascular cylinder and the meristematic band responsible for tuberisation was named the primary thickening meristem (Martins and Appezzato-da-Glória, 2006Martins, AR. and Appezzato-Da-Gloria, B., 2006. Morfoanatomia dos órgãos vegetativos Smilax polyantha Griseb. (Smilacaceae). Brazilian Journal of Botany, vol. 29, no. 4, p. 555-567. http://dx.doi.org/10.1590/S0100-84042006000400005.
http://dx.doi.org/10.1590/S0100-84042006...
).
Therefore, this study presents the morphology and anatomy of the underground organs of Smilax fluminensis and S. syphilitica to provide information for more accurate identification of the commercialised material and elucidating the rhizophore thickening process.
2 Material and Methods
Three individuals were analysed from each species. The S. syphiliticaHumboldt and Bonpland ex Willdenow individuals were collected in the Atlantic Forest, in Santa Tereza, Espírito Santo state, and S. fluminensis Steud. individuals were collected from the riparian forest area in the municipality of Itirapina, São Paulo state, Brazil. The collected material was identified by Dr. Regina Helena Potsch Andreata, an expert in the genus Smilax in Brazil, and the voucher specimens were added to the Herbarium of the School of Agriculture (ESA) under the numbers 107665 (S. syphilitica) and 107633 (S. fluminensis).
Rhizophores and adventitious roots of different diameters in the underground system were analysed. Samples were fixed in FAA 50 (1:1:8 formaldehyde, glacial acetic acid, and 50% ethanol) (Johansen, 1940JOHANSEN, A., 1940. Plant microtechnique. New York: McGraw-Hill.). For better fixation, the samples were subjected to a vacuum to remove the air in the tissues. They were then dehydrated in an ethanol series and embedded in hydroxyethyl methacrylate (LeicaTM Historesin); the obtained blocks were cut into 5-10 μm thick sections with a rotary microtome. Berberine-aniline blue staining was used for detecting suberin and lignin for identifying Casparian bands (Brundrett et al., 1988Brundrett, MC., Enstone, DE. and Peterson, CA., 1988. A berberine-aniline blue fluorescent staining procedure for suberine, lignin, and callose in plant tissue. Protoplasma, vol. 146, no. 2-3, p. 133-142. http://dx.doi.org/10.1007/BF01405922.
http://dx.doi.org/10.1007/BF01405922...
). For the remaining analyses, the material was stained with safranin and astra blue (Burger and Richter, 1991BURGER, LM. and RICHTER, HG., 1991. Anatomia da madeira. São Paulo: Nobel.) or with 0.05% toluidine blue in phosphate buffer and citric acid at pH 4-6 (Sakai, 1973Sakai, WS., 1973. Simple method for differential staining of paraffin embedded plant material using toluidine blue o. Stain Technology, vol. 48, no. 5, p. 247-249. PMid:4126691.). The stained slides were mounted in Entellan synthetic resin.
The images were digitally captured with a Leica DMLB microscope (LeicaTM - Wetzlar, Germany) using a video camera plugged into a computer utilising the IM50 (LeicaTM - Wetzlar, Germany) software for image analysis. The berberine-aniline blue staining was visualised with ultraviolet light.
3 Results
Regarding the rhizophores (Figures 1-4), the two species have in common the rigidity, thickening in the nodal regions, and the formation of white (Figure 2) and brown adventitious roots (Figures 3-4). Smilax fluminensisindividuals due to its elongated internodes (Figure 1) known as “runners”, is distributed in parallel to the soil surface and can reach between 7 and 12 m until a new aerial branch is emitted. The branches are emitted from thickened nodes (Figures 1-3). There is no horizontal development in S. syphilitica, because the internodes are short and the thickened nodes and the aerial branches are too close (Figure 4). In both species, the underground system is located at a depth between 10 and 15 cm.

Underground systems of Smilax fluminensis Steud (1-3) and S. syphilitica Humboldt and Bonpland ex Willdenow (4). 1. Part of an underground system demonstrating that the thickened nodes (arrows) from which the aerial branches are emitted are distant from one another due to the internode elongation. 2-3 Details of the thickened nodes forming white (2, arrow) and brown (3, arrow) adventitious roots. Note that these roots appear at the ends of the tuberous nodal areas. 4. Complete underground system with extremely short internodes and the presence of brown adventitious roots (arrow).
From the nodal area undergoing the tuberisation process (Figures 5-9), it was possible to observe that the swelling process of the rhizophore occurs through the meristematic activity of the axillary buds (Figures 5-7) and through divisions in the pericyclic cell layers underlying the endodermis, which can be recognised due to the parietal thickening in lipophilic and phenolic substances (Figures 8-9), near the bud.

Cross-sections of the nodal region of Smilax fluminensis Steud (5-7) and S.syphilitica Humboldt and Bonpland ex Willdenow (8-9). 5-7 Note the different stages of thickening with no change in the original vascular cylinder of the nodal region (Vc), which demonstrates the meristematic activity underlying the axillary bud. 7. Note the bilateral thickening (arrows). 8. Tuberisation initiates in the region below the axillary bud and divisions of the pericyclic cell layers underlying the endodermis (En) with thickened walls. Ab = axillary bud.
It is notable that tuberisation is generally bilateral (Figure 7), and an adventitious root is formed at the end of each projection (Figure 10) whose pericyclic origin is evidenced by the continuity of the endodermis with U-shaped thickening (Figure 11).

Smilax fluminensis Steud. 10. Cross-section of the end of a nodal tuberisation displaying the emission of an adventitious root. 11. Detail of the section indicated in the previous figure demonstrating the endoderm with thickened walls (arrow).
In both studied species, the adult rhizophore is thickened and exhibits a uniseriate epidermis (Figure 12) with stomata. Phenolic or raphid idioblasts occur between several layers of the cortical parenchyma cells (Figure 12). Peripheral bundles (Figure 13) are commonly found, often surrounded by cells whose parietal thickness is similar to that of the endodermis (compare Figures 12 and 13). The vascular cylinder has a pericycle with several cell layers elongated in the anticlinal direction (Figure 14). There are smaller collateral bundles distributed among the cells of the proliferated pericycle, and the remaining collateral bundles are larger and randomly distributed, as illustrated in Figure 9. There is accumulation of starch grains throughout the vascular cylinder, but with higher concentration in the pericyclic cells (Figure 14) and surrounding the vascular bundles (Figure 15). The endodermis with parietal thickening delimits the cortex and vascular cylinder of the rhizophores (Figure 16).

Cross-sections of the adult rhizophore of SmilaxsyphiliticaHumboldt and Bonpland ex Willdenow (12) and S. fluminensis Steud (13-16). 12. Note the uniseriate epidermis, the cortex with idioblasts between the cortical parenchyma cells, the endodermis with parietal thickening, and two smaller vascular bundles between the pericycle cells (circles). 13. Peripheral vascular bundle with cells similar to the endodermal cells (arrows). 14-15. Accumulation of starch grains evidenced by iodinated zinc chloride in the pericyclic layers (14) and surrounding the vascular bundle (15). 16. Note the emission of an adventitious root and the endodermis of the adult rhizophore. Ar = adventitious root; En = endodermis; Id = idioblasts; Pe = pericyclic cells; Vb = vascular bundles.
In both species, the adventitious root system of the rhizophore consists of roots at different stages of development. There are white tender roots of a larger diameter and brown rigid roots of a variable diameter (Figures 2-4). The white roots represent the youngest stage, characterised by root hairs and a uniseriate epidermis (Figure 17). The cortex displays an exodermis with Casparian bands (Figures 18-19), followed by the cortical parenchyma consisting of isodiametric cells. The cells of the last parenchyma cortical layer exhibit thickening in the inner periclinal wall and in the radial walls (Figure 20). The endodermis has more elongated cells in the periclinal direction and walls with Casparian bands (Figure 20). The vascular cylinder is a polyarch, and the pericycle is formed by approximately four cell layers with walls that become gradually thicker (Figures 22-24). Lysis occurs throughout the development and is accompanied by gradual removal of the epidermis and the entire cortical parenchyma (Figure 21). The root becomes brown and rigid at this stage, and the covering function is then performed by periclinal cell walls of the last cortical parenchyma layer and by the endodermis, which at this stage has thick and highly lignified walls (Figures 23-24). These walls have a brown colour due to the accumulation of phenolic compounds, which, in S. syphilitica, can be also present within the endodermal cells (Figure 24). The pith contains spherical and polyhedral starch grains (Figure 25).
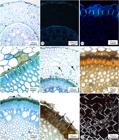
Cross-sections of the root of Smilax fluminensis Steud (17-23) and S.syphilitica Humboldt and Bonpland ex Willdenow (24-25). 17. White root covered by the epidermis. 18. White root stained with berberine-aniline blue, observed under ultraviolet light. 19. Detail of the exodermis with Casparian bands. 20. Cells of the last cortical parenchyma layer with thick walls and tall endodermal cells. 21. Beginning of the lysigenous process (arrows) of the cortical cells. 22. Detail of the parietal thickening of the endodermis with a U-shaped thickening and pericycle. 23. Brown root covered by the cell wall of the last cortical parenchyma layer and endodermis. 24. Detail of the brown root lining with phenolic compounds evidenced by the ferric chloride on the wall and lumen (arrows) of the endodermal cells. 25. Spherical and polyhedral starch grains located in the medulla, observed under polarised light.
4 Discussion
The white and brown adventitious roots of the root system, as well as their ontogenetic processes, are extremely similar to those described for S. polyantha (Martins and Appezzato-da-Glória, 2006Martins, AR. and Appezzato-Da-Gloria, B., 2006. Morfoanatomia dos órgãos vegetativos Smilax polyantha Griseb. (Smilacaceae). Brazilian Journal of Botany, vol. 29, no. 4, p. 555-567. http://dx.doi.org/10.1590/S0100-84042006000400005.
http://dx.doi.org/10.1590/S0100-84042006...
), S. subsessifolia (Guimarães et al., 2010Guimarães, A.R., Costa, C.G. and Andreata, R.H.P., 2010. Morfoanatomia do sistema subterrâneo de Smilax subsessiliflora (Smilacaceae). Rodriguésia, vol. 61, no. 2, p. 181-194.), S. brasiliensis, S. campestris, and S. cissoides (Martins et al., 2010Martins, AR., Pütz, N., Soares, AN., Bombo, AB. and Appezzato-Da-Gloria, B., 2010. New approaches to underground system in Brazilian Smilax species (Smilacaceae). The Journal of the Torrey Botanical Society, Athens, vol. 137, no. 2-3, p. 220-235. http://dx.doi.org/10.3159/10-RA-024R.1.
http://dx.doi.org/10.3159/10-RA-024R.1...
). The distribution of the phenolic and crystalliferous idioblasts, the circular arrangement of the primary phloem, and the absence of metaxylem elements in the centre of the structure are also common among other species already described in the literature and have allowed identification of the adulteration of products being sold as sarsaparilla (Soares, 2013Soares, MKM., 2013. Anatomia e perfil químico da salsaparrilha comercializada no Estado de São Paulo. Piracicaba: Universidade de São Paulo. 69 p. Dissertação de Mestrado em Fisiologia e Bioquímica de Plantas.).
The elongated internodes in the underground system were only verified in S. fluminensis, similar to that described in S. rufescens (Martins et al., 2010Martins, AR., Pütz, N., Soares, AN., Bombo, AB. and Appezzato-Da-Gloria, B., 2010. New approaches to underground system in Brazilian Smilax species (Smilacaceae). The Journal of the Torrey Botanical Society, Athens, vol. 137, no. 2-3, p. 220-235. http://dx.doi.org/10.3159/10-RA-024R.1.
http://dx.doi.org/10.3159/10-RA-024R.1...
).
The epidermis with stomata in rhizophores, most likely resulting from the derivation from aerial structures (Andreata and Menezes, 1999Andreata, RHP. and MENEZES, NL., 1999. Morfoanatomia do embrião, desenvolvimento pós-seminal e origem do rizóforo de Smilax quinquenervia Vell. Boletim de Botânica da Universidade de São Paulo, vol. 18, p. 39-51. http://dx.doi.org/10.11606/issn.2316-9052.v18i0p39-51.
http://dx.doi.org/10.11606/issn.2316-905...
; Appezzato-da-Glória, 2003APPEZZATO-DA-GLÓRIA, B., 2003. Morfologia de sistemas subterrâneos: histórico e evolução do conhecimento no Brasil. Ribeirão Preto: A.S. Pinto.), is a pattern already recorded in other Smilax species (Andreata and Menezes, 1999Andreata, RHP. and MENEZES, NL., 1999. Morfoanatomia do embrião, desenvolvimento pós-seminal e origem do rizóforo de Smilax quinquenervia Vell. Boletim de Botânica da Universidade de São Paulo, vol. 18, p. 39-51. http://dx.doi.org/10.11606/issn.2316-9052.v18i0p39-51.
http://dx.doi.org/10.11606/issn.2316-905...
; Palhares and Silveira, 2005Palhares, D. and Silveira, CES., 2005. Estudo anatômico do rizoma de A. DC. (Smilacaceae). Smilax goyazanaRevista Brasileira de Plantas Medicinais, vol. 8, no. 1, p. 52-62.; Guimarães et al., 2010Guimarães, A.R., Costa, C.G. and Andreata, R.H.P., 2010. Morfoanatomia do sistema subterrâneo de Smilax subsessiliflora (Smilacaceae). Rodriguésia, vol. 61, no. 2, p. 181-194.). However, in other Smilax rhizophores, the covering function is performed by peridermis (Caponetti and Quimby, 1956Caponetti, JD. and Quimby, MW., 1956. The comparative anatomy of certain species of Smilax. Journal of the American Pharmaceutical Association, vol. 45, no. 10, p. 691-696. http://dx.doi.org/10.1002/jps.3030451011. PMid:13366812.
http://dx.doi.org/10.1002/jps.3030451011...
) or by epidermis and cortex (Martins and Appezzato-da-Glória, 2006Martins, AR. and Appezzato-Da-Gloria, B., 2006. Morfoanatomia dos órgãos vegetativos Smilax polyantha Griseb. (Smilacaceae). Brazilian Journal of Botany, vol. 29, no. 4, p. 555-567. http://dx.doi.org/10.1590/S0100-84042006000400005.
http://dx.doi.org/10.1590/S0100-84042006...
).
The large amount of raphid idioblasts found in the cortex of the analysed rhizophores suggests an investment in an anti-herbivore strategy, as observed in the parenchyma of the tuberous roots of Asphodelusaestivus (Xanthorrhoeaceae) (Sawidis et al., 2005Sawidis, T., KalyVa, S. and Delivopoulos, S., 2005. The root-tuber anatomy of Asphodelus aestivus. Flora, vol. 200, no. 4, p. 332-338. http://dx.doi.org/10.1016/j.flora.2004.10.002.
http://dx.doi.org/10.1016/j.flora.2004.1...
). In addition, because the rhizophore is an organ capable of expanding the rhizosphere and conferring resistance and vegetative propagation (Andreata and Menezes, 1999Andreata, RHP. and MENEZES, NL., 1999. Morfoanatomia do embrião, desenvolvimento pós-seminal e origem do rizóforo de Smilax quinquenervia Vell. Boletim de Botânica da Universidade de São Paulo, vol. 18, p. 39-51. http://dx.doi.org/10.11606/issn.2316-9052.v18i0p39-51.
http://dx.doi.org/10.11606/issn.2316-905...
), it is understandable that the rhizophore accumulates large amounts of starch grains, such as the ones regularly found in the cortex and throughout the vascular cylinder of the structure in both studied species.
In this study, unlike reports in other species of Smilax (Martins and Appezzato-da-Glória, 2006Martins, AR. and Appezzato-Da-Gloria, B., 2006. Morfoanatomia dos órgãos vegetativos Smilax polyantha Griseb. (Smilacaceae). Brazilian Journal of Botany, vol. 29, no. 4, p. 555-567. http://dx.doi.org/10.1590/S0100-84042006000400005.
http://dx.doi.org/10.1590/S0100-84042006...
; Martins et al., 2010Martins, AR., Pütz, N., Soares, AN., Bombo, AB. and Appezzato-Da-Gloria, B., 2010. New approaches to underground system in Brazilian Smilax species (Smilacaceae). The Journal of the Torrey Botanical Society, Athens, vol. 137, no. 2-3, p. 220-235. http://dx.doi.org/10.3159/10-RA-024R.1.
http://dx.doi.org/10.3159/10-RA-024R.1...
), it was possible to observe the endodermis with parietal thickening delimiting the cortex and vascular cylinder of the rhizophores. This analysis made it possible to determine that the cell proliferation leading to nodal thickening occurs in the proximity of the axillary bud in the layers underlying the endodermis, i.e., there is a collaboration from the pericycle in this process. Although it was not possible to establish the exact boundary between the cortex and vascular cylinder in S. campestris, S. goyazana, and S. oblongifolia (Martins et al., 2010Martins, AR., Pütz, N., Soares, AN., Bombo, AB. and Appezzato-Da-Gloria, B., 2010. New approaches to underground system in Brazilian Smilax species (Smilacaceae). The Journal of the Torrey Botanical Society, Athens, vol. 137, no. 2-3, p. 220-235. http://dx.doi.org/10.3159/10-RA-024R.1.
http://dx.doi.org/10.3159/10-RA-024R.1...
), the cell proliferation is extremely similar to that observed in our study. Furthermore, in the analysed species and in S. campestris, S. goyazana, and S. oblongifolia (Martins et al., 2010Martins, AR., Pütz, N., Soares, AN., Bombo, AB. and Appezzato-Da-Gloria, B., 2010. New approaches to underground system in Brazilian Smilax species (Smilacaceae). The Journal of the Torrey Botanical Society, Athens, vol. 137, no. 2-3, p. 220-235. http://dx.doi.org/10.3159/10-RA-024R.1.
http://dx.doi.org/10.3159/10-RA-024R.1...
) there is no establishment of a meristematic area similar to that described in S. polyantha by Martins and Appezzato-da-Glória (2006)Martins, AR. and Appezzato-Da-Gloria, B., 2006. Morfoanatomia dos órgãos vegetativos Smilax polyantha Griseb. (Smilacaceae). Brazilian Journal of Botany, vol. 29, no. 4, p. 555-567. http://dx.doi.org/10.1590/S0100-84042006000400005.
http://dx.doi.org/10.1590/S0100-84042006...
. The data presented here support the proposal by Menezes et al. (2012)Menezes, NL., Elbl, PM., Cury, G., Sasaki, KLM., Appezzato-Da-Glória, B., Costa, GO. and Silva, CG., 2012. The meristematic activity of the endodermis and the pericycle and its role in the primary thickening of stems in monocotyledonous plants. Plant Ecology & Diversity, vol. 5, no. 2, p. 153-165. http://dx.doi.org/10.1080/17550874.2011.604925.
http://dx.doi.org/10.1080/17550874.2011....
that, in monocotyledons, the pericycle itself is involved in the thickening process with the formation of new vascular bundles and parenchyma. Moreover, the continuity of the endodermis of the rhizophores and adventitious roots, observed on their emission sites, corroborates the statements that the endodermis is a continuous tissue present in roots and stems (Menezes et al., 2005Menezes, NL., Silva, DC., Arruda, RCO., Melo-de-Pinna, GFA., Cardoso, VA., Castro, NM., Scatena, VL. and Scremin-Dias, E., 2005. Meristematic activity of the Endodermis and the Pericycle in the primary thickening in monocotyledons: considerations on the “PTM”. Anais da Academia Brasileira de Ciencias, vol. 77, no. 2, p. 259-274. http://dx.doi.org/10.1590/S0001-37652005000200006. PMid:15895162.
http://dx.doi.org/10.1590/S0001-37652005...
). The involvement of the axillary bud in the nodal thickening process and consequent rooting in the studied species had already been observed in stem cuttings of S. fluminensis (Soares et al., 2011SOARES, AN., NOVEMBRE, ADLC., MARTINS, AR., PIEDADE, SMS. and APPEZZATO-DA-GLÓRIA, B., 2011. Propagation studies in Smilax fluminensis Steud. (Smilacaceae). Ciência Rural, vol. 41, no. 10, p. 1762-1768. http://dx.doi.org/10.1590/S0103-84782011001000014.
http://dx.doi.org/10.1590/S0103-84782011...
). The authors found that adventitious root formation did not involve callus formation and neither occur in the basal region of the cuttings, but instead occurred in the basal region of the swollen bud.
Regarding the peripheral bundles of the rhizophores, only a few cells had parietal thickening similar to that of the endodermis, but in Smilax goyazana, the endodermis was described as surrounding all the vascular bundles (Palhares and Silveira, 2005Palhares, D. and Silveira, CES., 2005. Estudo anatômico do rizoma de A. DC. (Smilacaceae). Smilax goyazanaRevista Brasileira de Plantas Medicinais, vol. 8, no. 1, p. 52-62.).
In conclusion, the axillary bud and the pericycle participate in the process of rhizophore nodal thickening. The adventitious roots have common features among the Smilax species, which may facilitate the diagnosis of adulteration in drugs commercialised as sarsaparilla.
-
(With 25 figures)
References
- Andreata, RHP. and MENEZES, NL., 1999. Morfoanatomia do embrião, desenvolvimento pós-seminal e origem do rizóforo de Smilax quinquenervia Vell. Boletim de Botânica da Universidade de São Paulo, vol. 18, p. 39-51. http://dx.doi.org/10.11606/issn.2316-9052.v18i0p39-51.
» http://dx.doi.org/10.11606/issn.2316-9052.v18i0p39-51 - Andreata, RHP., 1997. Revisão das espécies brasileiras do gênero Linnaeus (Smilacaceae). SmilaxPesquisa Botânica, vol. 47, p. 7-244.
- Andreata, RHP., 2009. A new species of and a key to all species of Minas Gerais, Brazil. SmilaxSystematic Botany, vol. 34, no. 1, p. 28-31. http://dx.doi.org/10.1600/036364409787602302.
» http://dx.doi.org/10.1600/036364409787602302 - APPEZZATO-DA-GLÓRIA, B., 2003. Morfologia de sistemas subterrâneos: histórico e evolução do conhecimento no Brasil. Ribeirão Preto: A.S. Pinto.
- Brundrett, MC., Enstone, DE. and Peterson, CA., 1988. A berberine-aniline blue fluorescent staining procedure for suberine, lignin, and callose in plant tissue. Protoplasma, vol. 146, no. 2-3, p. 133-142. http://dx.doi.org/10.1007/BF01405922.
» http://dx.doi.org/10.1007/BF01405922 - BURGER, LM. and RICHTER, HG., 1991. Anatomia da madeira. São Paulo: Nobel.
- Caponetti, JD. and Quimby, MW., 1956. The comparative anatomy of certain species of Smilax. Journal of the American Pharmaceutical Association, vol. 45, no. 10, p. 691-696. http://dx.doi.org/10.1002/jps.3030451011. PMid:13366812.
» http://dx.doi.org/10.1002/jps.3030451011 - Cunha, NS., 1937. Da salsaparrilha a japecanga. Tribuna Farmaceutica, vol. 5, p. 145-150.
- Farmacopéia, 1929. Codigo pharmaceutico brasileiro: pharmacopeia dos Estados Unidos do Brasil. Rio de Janeiro: Companhia Editora Nacional. 1149 p. Redigida por Rodolpho Albino Dias da Silva.
- Guimarães, A.R., Costa, C.G. and Andreata, R.H.P., 2010. Morfoanatomia do sistema subterrâneo de Smilax subsessiliflora (Smilacaceae). Rodriguésia, vol. 61, no. 2, p. 181-194.
- Holm, T., 1890. Contribution to the knowledge of the germination of some North American plants. Memoirs of the Torrey Botanical Club, vol. 2, p. 57-108.
- JOHANSEN, A., 1940. Plant microtechnique. New York: McGraw-Hill.
- Judd, WS., Campbell, CS., Kellogg, EA., STEVENS, PF. and DONOGHUE, MJ., 2009. Sistemática vegetal: um enfoque filogenético. Porto Alegre: Artmed.
- Martins, AR. and Appezzato-Da-Gloria, B., 2006. Morfoanatomia dos órgãos vegetativos Smilax polyantha Griseb. (Smilacaceae). Brazilian Journal of Botany, vol. 29, no. 4, p. 555-567. http://dx.doi.org/10.1590/S0100-84042006000400005.
» http://dx.doi.org/10.1590/S0100-84042006000400005 - Martins, AR., Pütz, N., Novembre, ADLC., Piedade, SMS. and Appezzato-Da-Glória, B., 2011. Seed germination and seedling morphology of Smilax polyantha (Smilacaceae). Biota Neotropica, vol. 11, no. 2, p. 1-7. http://dx.doi.org/10.1590/S1676-06032011000200003.
» http://dx.doi.org/10.1590/S1676-06032011000200003 - Martins, AR., Pütz, N., Soares, AN., Bombo, AB. and Appezzato-Da-Gloria, B., 2010. New approaches to underground system in Brazilian Smilax species (Smilacaceae). The Journal of the Torrey Botanical Society, Athens, vol. 137, no. 2-3, p. 220-235. http://dx.doi.org/10.3159/10-RA-024R.1.
» http://dx.doi.org/10.3159/10-RA-024R.1 - Medeiros, MFT., Senna-Valle, L. and Andreata, RHP., 2007. Histórico e o uso da “salsaparrilha” ( spp.) pelos boticários no Mosteiro de São Bento. SmilaxRevista Brasileira de Biociências, vol. 5, p. 27-29.
- Menezes, NL., Elbl, PM., Cury, G., Sasaki, KLM., Appezzato-Da-Glória, B., Costa, GO. and Silva, CG., 2012. The meristematic activity of the endodermis and the pericycle and its role in the primary thickening of stems in monocotyledonous plants. Plant Ecology & Diversity, vol. 5, no. 2, p. 153-165. http://dx.doi.org/10.1080/17550874.2011.604925.
» http://dx.doi.org/10.1080/17550874.2011.604925 - Menezes, NL., Silva, DC., Arruda, RCO., Melo-de-Pinna, GFA., Cardoso, VA., Castro, NM., Scatena, VL. and Scremin-Dias, E., 2005. Meristematic activity of the Endodermis and the Pericycle in the primary thickening in monocotyledons: considerations on the “PTM”. Anais da Academia Brasileira de Ciencias, vol. 77, no. 2, p. 259-274. http://dx.doi.org/10.1590/S0001-37652005000200006. PMid:15895162.
» http://dx.doi.org/10.1590/S0001-37652005000200006 - Ming, LC., 1994. Estudos e pesquisas de plantas medicinais na agronomia. Horticultura Brasileira, vol. 12, p. 3-9.
- Palhares, D. and Silveira, CES., 2005. Estudo anatômico do rizoma de A. DC. (Smilacaceae). Smilax goyazanaRevista Brasileira de Plantas Medicinais, vol. 8, no. 1, p. 52-62.
- Sakai, WS., 1973. Simple method for differential staining of paraffin embedded plant material using toluidine blue o. Stain Technology, vol. 48, no. 5, p. 247-249. PMid:4126691.
- Sawidis, T., KalyVa, S. and Delivopoulos, S., 2005. The root-tuber anatomy of Asphodelus aestivus. Flora, vol. 200, no. 4, p. 332-338. http://dx.doi.org/10.1016/j.flora.2004.10.002.
» http://dx.doi.org/10.1016/j.flora.2004.10.002 - SOARES, AN., NOVEMBRE, ADLC., MARTINS, AR., PIEDADE, SMS. and APPEZZATO-DA-GLÓRIA, B., 2011. Propagation studies in Smilax fluminensis Steud. (Smilacaceae). Ciência Rural, vol. 41, no. 10, p. 1762-1768. http://dx.doi.org/10.1590/S0103-84782011001000014.
» http://dx.doi.org/10.1590/S0103-84782011001000014 - Soares, MKM., 2013. Anatomia e perfil químico da salsaparrilha comercializada no Estado de São Paulo. Piracicaba: Universidade de São Paulo. 69 p. Dissertação de Mestrado em Fisiologia e Bioquímica de Plantas.
- Stellfeld, C., 1938. A salsaparilha do Paraná e a ausência do parênquima cortical. Tribuna Farmaceutica, vol. 6, p. 5-7.
Publication Dates
-
Publication in this collection
25 Aug 2015 -
Date of issue
Aug 2015
History
-
Received
26 Nov 2013 -
Accepted
04 Feb 2014





