
Abstract
Microclimatic conditions of tropical forest favour the high richness of bryophytes, which by being sensitive to environmental changes, are important indicators of habitat conditions. The aim of this study was to determine the richness and species composition of the bryophyte flora in fragments of terra firme forest on the great curve of the Xingu River, Pará state, Brazil. The collections were made in August and September 2012 in 14 fragments, in which were installed two plots per fragment, one at the edge and one inside, measuring 10 × 10 m each. The results showed 77 species in 45 genera and 18 families. Lejeunea setiloba Spruce and Marchesinia brachiata (Sw.) Schiffn. are new records for Pará state. The richness families in this study were the ones typically found in tropical forest surveys. A high richness of rare species in comparison to common ones, a pattern usually observed for plants in tropical forests was not reported in this study, probably due to historical fragmentation and disturbance in the area. The richness and species composition were determined mainly by the physiognomic characteristics of the studied forest fragments.
Keywords:
brioflora; Amazonian flora; liverworts; mosses
Resumo
As condições microclimáticas das florestas tropicais favorecem elevada riqueza de briófitas, que por serem sensíveis às alterações ambientais, são importantes indicadoras das condições dos habitats. O objetivo deste trabalho foi determinar a riqueza e a composição florística da brioflora de fragmentos florestais de terra firme na região da Volta Grande do Xingu, Pará, Brasil. As coletas foram realizadas em Agosto e Setembro de 2012, em 14 fragmentos e em cada um destes foram plotadas duas parcelas, uma na borda e outra no interior, medindo 10 × 10 m (cada). Foram registradas 77 espécies, distribuídas em 45 gêneros e 18 famílias. Lejeunea setiloba Spruce e Marchesinia brachiata (Sw.) Schiffn. são novos registros para o Pará. As famílias mais ricas neste estudo foram aquelas tipicamente encontradas em levantamentos em florestas tropicais. A elevada riqueza de espécies raras em comparação às comuns, padrão comumente reportado para plantas em florestas tropicais, não foi observada, provavelmente devido ao histórico de fragmentação e distúrbios na área de estudo. A riqueza e a composição de espécies foram determinadas principalmente pelas características fisionômicas dos fragmentos estudados.
Palavras-chave:
brioflora; flora da Amazônia; hepáticas; musgos
1 Introduction
In several regions of the world, bryophytes are an important component of vegetation (Glime, 2007Glime, JM., 2007. Economic and ethnic uses of bryophytes. In Editorial Committee (Ed.). Flora of North America North of Mexico: bryophyta, part 1. New York: Oxford Universit Press. p. 14-41. vol. 27.). Large variations in landscapes and climates favour the richness and diversity of bryophytes (Gradstein et al., 2001Gradstein, SR., Churchill, SP. and Salazar, NA., 2001. Guide to the Bryophytes of Tropical America. Memoirs of the New York Botanical Garden, vol. 86, p. 1-577.), particularly in tropical forests (Gradstein and Pócs, 1989Gradstein, SR. and Pócs, T., 1989. Bryophytes. In LIETH, H. and WERGER, MJA. (Eds.). Tropical rain forest ecosystems. Amsterdam: Elseiver Science Publishers. p. 311-325. http://dx.doi.org/10.1016/B978-0-444-42755-7.50022-5.
http://dx.doi.org/10.1016/B978-0-444-427...
). In tropical regions their exuberance and diversity is especially noticed, increasing in altitudes above 1,500 m (Gradstein et al., 2001Gradstein, SR., Churchill, SP. and Salazar, NA., 2001. Guide to the Bryophytes of Tropical America. Memoirs of the New York Botanical Garden, vol. 86, p. 1-577.).
The Amazon and Atlantic Forest are the two most significant phytogeographic regions in terms of richness for bryophytes in Brazil (Santos et al., 2011Santos, ND., Costa, DP., Kinoshita, LS. and Shepherd, GJ., 2011. Aspectos brioflorísticos e fitogeográficos de duas formações costeiras de Floresta Atlântica da Serra do Mar, Ubatuba/SP, Brasil. Biota Neotropica, vol. 11, no. 2, p. 425-438. http://dx.doi.org/10.1590/S1676-06032011000200040.
http://dx.doi.org/10.1590/S1676-06032011...
). Even the Amazon being the largest extension of humid tropical forest characterized by a remarkable richness of species and high levels of endemism (Mittermeier et al., 1992Mittermeier, RA., Werner, T., Ayres, JM. and Fonseca, GAB., 1992. O país da megadiversidade. Campinas. Available from: <http://www.bdpa.cnptia.embrapa.br>. Access in: June 2012.
http://www.bdpa.cnptia.embrapa.br...
), it is the second most important region for the bryoflora in the country. The higher diversity and endemism of bryophytes in Brazil is in the Atlantic forest, greatly due to its altitudinal and latitudinal range (Gradstein et al, 2001Gradstein, SR., Churchill, SP. and Salazar, NA., 2001. Guide to the Bryophytes of Tropical America. Memoirs of the New York Botanical Garden, vol. 86, p. 1-577.).
The landscape of the Amazon is very diverse and different forest types can be distinguished, among which stands out the terra firme forest that occupies about 90% of the Amazon territory (Pires, 1973Pires, JM., 1973. Tipos de vegetação da Amazônia. Belém: Museu Goeldi. p. 179-202. Publicações Avulsas, vol. 20.; Pires and Prance, 1985Pires, JM. and Prance, GT., 1985. The vegetation types of the brazilian Amazon. In PRANCE, GT. and LOVEJOY, TE. (Eds.). Key environments: Amazonia. Oxford: Pergamon Press. p. 109-145.; Gradstein and Costa, 2003Gradstein, SR. and Costa, DP., 2003. The Hepaticae and Anthocerotae of Brazil. Memoirs of the New York Botanical Garden, vol. 87, p. 1-336.). This forest type is characterized mostly by the floristic heterogeneity, with high richness and diversity, as well as high complexity regarding its composition, distribution and density of species (Oliveira and Mori, 1999Oliveira, AA. and Mori, SA., 1999. A central Amazonian terra firme forest. I. High tree species richness on poor soils. Biodiversity and Conservation, vol. 8, no. 9, p. 1219-1244. http://dx.doi.org/10.1023/A:1008908615271.
http://dx.doi.org/10.1023/A:100890861527...
; Ter Steege et al., 2000Ter Steege, H., Sabatier, D., Castellanos, H., Van Andel, T., Duivenvoorden, J., Adalardo De Oliveira, A., Ek, R., Lilwah, R., Maas, P. and Mori, S., 2000An analysis of Amazonian floristic composition, including those of the Guiana Shield. Journal of Tropical Ecology, vol. 16, no. 6, p. 801-828. http://dx.doi.org/10.1017/S0266467400001735.
http://dx.doi.org/10.1017/S0266467400001...
; Gama et al., 2005Gama, JRV., Souza, AL., Martins, SV. and Souza, DR., 2005. Comparação entre florestas de várzea e de terra firme do estado do Pará. Revista Árvore, vol. 29, no. 4, p. 607-616. http://dx.doi.org/10.1590/S0100-67622005000400013.
http://dx.doi.org/10.1590/S0100-67622005...
).
Studies on bryophytes in the state of Pará were performed in different forest formations and various parts of the state as in the surroundings of the Lake Tucuruí (Ilkiu-Borges et al., 2004Ilkiu-Borges, AL., Tavares, ACC. and Lisboa, RCL., 2004. Briófitas da Ilha de Germoplasma, reservatório de Tucuruí, Pará, Brasil. Acta Botanica Brasilica, vol. 18, no. 3, p. 689-692. http://dx.doi.org/10.1590/S0102-33062004000300026.
http://dx.doi.org/10.1590/S0102-33062004...
; Garcia et al., 2014Garcia, ET., Ilkiu-Borges, AL. and Tavares-Martins, ACC., 2014. Brioflora de Florestas de terra firme na Área de Proteção Ambiental do Lago de Tucuruí PA, Brasil. Hoehnea, vol. 41, no. 4, p. 499-514. http://dx.doi.org/10.1590/2236-8906-10/2013.
http://dx.doi.org/10.1590/2236-8906-10/2...
), Serra dos Carajás (Moraes and Lisboa, 2006Moraes, ENR. and LISBOA, RCL., 2006. Musgos (Bryophyta) da Serra dos Carajás, Estado do Pará, Brasil. Boletim do Museu Paraense Emilio Goeldi. Série Ciências Naturais, vol. 1, p. 39-68.), Caxiuanã (Alvarenga and Lisboa, 2009Alvarenga, LDP. and Lisboa, RCL., 2009. Contribuição para o conhecimento da taxonomia, ecologia e fitogeografia de Briófitas da Amazônia Oriental. Acta Amazonica, vol. 39, no. 3, p. 495-504. http://dx.doi.org/10.1590/S0044-59672009000300003.
http://dx.doi.org/10.1590/S0044-59672009...
; Ilkiu-Borges et al., 2013Ilkiu-Borges, AL., Macêdo, LPC.,PEREIRA, MAV. and LISBOA, RCL., 2013. Briófitas em Caxiuanã: resultados do levantamento em duas parcelas da grade do PPBIO. In LISBOA, PLB. (Ed.). Caxiuanã: paraíso ainda preservado. Pará: Museu Paraense Emilio Goeldi. p. 287-295.), metropolitan region of Belém (Lisboa and Ilkiu-Borges, 1995Lisboa, RCL. and ILKIU-BORGES, AL., 1995. Diversidade das briófitas de Belém (PA) e seu potencial como indicadoras de poluição. Boletim do Museu Paraense Emilio Goeldi. Série Ciências Naturais, vol. 11, p. 199-225.; Moura et al., 2013Moura, OS., Ilkiu-Borges, AL. and Brito, ES., 2013. Brioflora (Bryophyta e Marchantiophyta) da Ilha do Combu, Belém, PA, Brasil. Hoehnea., vol. 40, no. 1, p. 143-165. http://dx.doi.org/10.1590/S2236-89062013000100008.
http://dx.doi.org/10.1590/S2236-89062013...
), Marajó Island (Lisboa et al., 1999Lisboa, RCL., Lima, MJL. and Maciel, UN., 1999. Musgos da Ilha de Marajó - II – Anajás, Pará, Brasil. Acta Amazonica, vol. 29, no. 2, p. 201-206. http://dx.doi.org/10.1590/1809-43921999292206.
http://dx.doi.org/10.1590/1809-439219992...
; Brito and Ilkiu-Borges, 2013Brito, ES. and Ilkiu-Borges, AL., 2013. Bryoflora of the municipalities of Soure and Cachoeira do Arari, on Marajó Island, in the state of Pará, Brazil. Acta Botanica Brasilica, vol. 27, no. 1, p. 124-141. http://dx.doi.org/10.1590/S0102-33062013000100013.
http://dx.doi.org/10.1590/S0102-33062013...
) and northeastern Pará (Santos and Lisboa, 2008Santos, RCP. and LISBOA, R., 2008. Musgos (Bryophyta) da Microrregião do Salgado Paraense e sua utilização como possíveis indicadores de ambientes perturbados. Rodriguésia, vol. 59, no. 2, p. 361-368.; Lisboa and Tavares, 2008Lisboa, RCL. and TAVARES, ACC., 2008. Briófitas de Santarém Novo, Pará. In JARDIM, MAG. and ZOGHBI, MGB. (Eds.). Flora Resex Chocoaré-Mato Grosso (PA): diversidades e usos. Museu Paraense Emílio Goeldi. p. 51-61.; Tavares-Martins et al., 2014Tavares-Martins, ACC., Lisboa, RCL. and Costa, DP., 2014. Bryophyte flora in upland forests at different successional stages and in the various strata of host trees in northeastern Pará, Brazil. Acta Botanica Brasilica, vol. 28, no. 1, p. 46-58. http://dx.doi.org/10.1590/S0102-33062014000100005.
http://dx.doi.org/10.1590/S0102-33062014...
). In the southwestern state, it is recorded only the inventory held by Lisboa and Ilkiu-Borges (2001)Lisboa, RCL. and ILKIU-BORGES, AL., 2001. Briófitas de São Luís do Tapajós, Município de Itaituba, com novas adições para o Estado do Pará. Boletim do Museu Paraense Emilio Goeldi. Série Ciências Naturais, vol. 17, p. 75-91. in São Luiz do Tapajós, in the municipality of Itaituba.
Over the past 40 years, the region of the great curve of the Xingu River, located in the southwestern Pará, went through an intense process of occupation, which resulted in the change of its original forest cover and about 60% were converted into areas for family farms, pastures, among others (Mausel et al., 1993Mausel, P., Wu, Y., Li, Y., Moran, EF. and Brondizio, ES., 1993. Spectral identification of successional stages following deforestation in the Amazon. Geocarto International, vol. 8, no. 4, p. 61-71. http://dx.doi.org/10.1080/10106049309354430.
http://dx.doi.org/10.1080/10106049309354...
; Salomão et al., 2007Salomão, RP., Vieira, ICG., Suemitsu, C., Rosa, NA., Almeida, SS., Amaral, DD. and Menezes, MPM., 2007. As florestas de Belo Monte na grande curva do rio Xingu, Amazônia Oriental. Boletim do Museu Paraense Emilio Goeldi. Série Ciências Naturais, vol. 2, p. 57-153.). As a result, the forests in the region are quite fragmented and poorly known regarding its richness and composition, especially about bryophytes.
The aim of this study was to determine the richness and species composition of the bryophyte flora in fragments of terra firme forest on the great curve of the Xingu River, Pará state.
2 Material and Methods
2.1 Study area
The study area corresponds to the region called great curve of the Xingu River, in the lower course of the river (Figure 1), southwest of Pará (Salomão et al., 2007Salomão, RP., Vieira, ICG., Suemitsu, C., Rosa, NA., Almeida, SS., Amaral, DD. and Menezes, MPM., 2007. As florestas de Belo Monte na grande curva do rio Xingu, Amazônia Oriental. Boletim do Museu Paraense Emilio Goeldi. Série Ciências Naturais, vol. 2, p. 57-153.). The region includes areas classified as dense and open ombrophilous forest and alluvial vegetation, directly influenced by the hydrological system of the Xingu River, in addition to recent and old secondary forests (Salomão et al., 2007Salomão, RP., Vieira, ICG., Suemitsu, C., Rosa, NA., Almeida, SS., Amaral, DD. and Menezes, MPM., 2007. As florestas de Belo Monte na grande curva do rio Xingu, Amazônia Oriental. Boletim do Museu Paraense Emilio Goeldi. Série Ciências Naturais, vol. 2, p. 57-153.). The predominant climate is humid tropical, with an average temperature of 26 °C and annual rainfall of 2,289 mm. A short dry season occurs during the months of August and September (Cunha and Ferreira, 2012Cunha, DA. and Ferreira, LV., 2012. Impacts of the Belo Monte hydroelectric dam construction on pioner vegetation formations along the Xingu River, Pará State, Brazil. Brazilian Journal of Botany, vol. 35, no. 2, p. 159-167. http://dx.doi.org/10.1590/S1806-99592012000200005.
http://dx.doi.org/10.1590/S1806-99592012...
; INMET, 2012Instituto Nacional de Meteorologia - INMET, 2012. Brasília. Available from: <http://www.inmet.gov.br>. Access in: Mar 2012.
http://www.inmet.gov.br...
).

Location map of forest fragments in the region of Volta Grande do Xingu, Pará state, Brazil. Source: Marcelo Cordeiro Thalês (MPEG).
The 14 fragments studied are located on the right bank of the river, in Vitória do Xingu, which has the largest forest cover in the region (Salomão et al., 2007Salomão, RP., Vieira, ICG., Suemitsu, C., Rosa, NA., Almeida, SS., Amaral, DD. and Menezes, MPM., 2007. As florestas de Belo Monte na grande curva do rio Xingu, Amazônia Oriental. Boletim do Museu Paraense Emilio Goeldi. Série Ciências Naturais, vol. 2, p. 57-153.). The fragment dimensions range from 7.96 to 3,784.20 hectares and altitudes from 57 to 187 m (Figure 1).
The fragments show similar physiognomies, with few clearings and humid environments, and were characterised by large sized trees, such as the species Theobroma speciosum Willd. ex Spreng. (Malvaceae), Inga spp. (Fabaceae), Pouteria spp. (Sapotaceae), Sterculia spp. (Malvaceae), Tachigali spp. (Fabaceae), Vouacapoua americana Aubl. (Fabaceae), Cenostigma tocantinum Ducke (Fabaceae), Bertholletia excelsa Bonpl. (Lecythidaceae), Alexa grandiflora Ducke (Fabaceae), and Schefflera morototoni (Aubl.) Maguire et al. (Araliaceae). In the edges, the dossal is usually a more open and formed by fewer trees species.
2.2 Collection and taxonomic identification
The collections were made in August and September 2012, following the techniques described by Yano (1989)Yano, O., 1989. Briófitas. In FIDALGO, O. and BONONI, VLR (Eds.). Técnicas de coleta, preservação e herborização de material botânico. São Paulo: Instituto de Botânica. p. 27-30.. The selection of the 14 terra firme forest fragments was carried out with the aim of including the largest possible number of areas along the great curve of the Xingu River, taking as selection criteria the same physiognomies. In each fragment were established two plots, one on edge and other inside, measuring 10 × 10 m (each). This method was applied with the intention to engage a larger species composition within the fragments, since it is known that the edge have effect in floristic composition. Plot size was based on the subplots of Santos et al. (2011)Santos, ND., Costa, DP., Kinoshita, LS. and Shepherd, GJ., 2011. Aspectos brioflorísticos e fitogeográficos de duas formações costeiras de Floresta Atlântica da Serra do Mar, Ubatuba/SP, Brasil. Biota Neotropica, vol. 11, no. 2, p. 425-438. http://dx.doi.org/10.1590/S1676-06032011000200040.
http://dx.doi.org/10.1590/S1676-06032011...
and for the collection of material, each plot was subdivided into five corridors 2 × 10 m, where traversing each one, the material were collected on all types of substrates only reaching the understory.
In preparing the material for identification were adopted the usual techniques for bryophytes and the determination of taxa was performed with the aid of taxonomic keys and descriptions found in the specialised literature. The taxonomic classifications adopted were those of Goffinet et al. (2009)Goffinet, B., Buck, WR. and Shaw, AJ., 2009. Morphology, anatomy, and classification of the Bryophyta. In GOFFINET, B. and SHAW, AJ. (Eds.). Bryophyte biology. 2nd ed. Cambridge: University Press Cambridge. p. 55-138. for the Bryophyta and Crandall-Stotler et al. (2009)Crandall-Stotler, B., Stotler, RE. and Long, DG., 2009. Morphology and classification of the Marchantiophyta. In GOFFINET, B. and SHAW, AJ. (Eds.). Bryophyte biology. 2nd ed. Cambridge: University Press Cambridge. p. 1-54. for Marchantiophyta. The testimony material was incorporated in the João Murça Pires Herbarium (MG).
2.3 Data analysis
The richness, floristic composition, geographic distribution, the absolute frequency of the species were analyzed, and also calculated the similarity between fragments. The existence of significant differences in richness and in the guilds of tolerance of the species between fragments was also verified.
To verify sample sufficiency, a species accumulation curve was generated using the Mao Tau function (Colwell et al., 2004COLWELL, RK., Mao, CX. and Chang, J., 2004. Interpolating, extrapolating, and comparing incidence-based species accumulation curves. Ecology, vol. 85, no. 10, p. 2717-2727. http://dx.doi.org/10.1890/03-0557.
http://dx.doi.org/10.1890/03-0557...
) with a confidence interval of 95%, for this was elaborate an matrix of presence/absence of species per fragment, subjected to analysis in EstimateS 8.2.0 program (Colwell, 2009Colwell, RK., 2009. EstimateS: Statistical estimation of species richness and shared species from samples. Storrs: University of Connecticut. Available from: <http://viceroy.eeb.uconn.edu/EstimateS>. Access in: Aug. 2012.
http://viceroy.eeb.uconn.edu/EstimateS...
).
The floristic composition was determined by the families, genera and species present in fragments and species richness described by the total of species inventoried in the study area.
The absolute frequency of the species was determined based on the number of occurrences (incidence/occurrence) of species in fragments. Three categories of frequency were defined, adapted from Silva and Pôrto (2007)SILVA, MPP. and PÔRTO, KC., 2007. Composição e riqueza de briófitas epíxilas em fragmentos florestais da Estação Ecológica de Murici, Alagoas. Revista Brasileira de Biologia, vol. 5, p. 243-245. and Garcia (2012)Garcia, ET., 2012. Briófitas (Bryophyta e Marchantiophyta) de Remanescentes Florestais no Reservatório de Tucuruí, Pará, Brasil. Pará: Universidade Federal Rural da Amazônia, Museu Paraense Emilio Goeldi. 87 p. Masters Dissertation.: rare (one to five occurrences), common (six to 19 occurrences) and constant (20 or more occurrences).
A matrix of presence and absence of 77 species listed in 14 fragments was prepared and calculated the floristic similarity by the Jaccard coefficient, followed by a cluster analysis (Hair Junior et al., 2006Hair JUNIOR, JF., Anderson, RE., Tathan, RL. and Black, WC., 2006. Análise multivariada de dados. Porto Alegre: Bookman. 593 p.) from the average linkage clustering method (UPGMA) using the MVSP 3.0 software.
To verify the existence of significant difference in species richness of bryophytes between fragments was applied a chi-squared test, which is the number of different species found in the sample from each fragment analyzed, calculated in the Biostat 5.0 software (Ayres et al., 2007Ayres, M., AYRES JÚNIOR, M., Ayres, DL. and Santos, AAS., 2007. BIOESTAT: aplicações estatísticas nas áreas das ciências bio-médicas. Belém: Instituto de Desenvolvimento Sustentável Mamirauá. 364 p.).
In the analysis of tolerance guilds, species were classified according to their tolerance to sunlight in: sun epiphytes (Es), shade epiphytes (Esh) and generalists (Gen). This classification was based on the works of Richards (1984)Richards, PW., 1984. The ecology of tropical forest bryophytes. In SCHUSTER, RM. (Ed.). New Manual of Bryology. Nichinan: Hattori Botanical Laboratory. p. 1233-1269. vol. 2., Cornelissen and Ter Steege (1989)Cornelissen, JH. and TER STEEGE, H., 1989. Distribuition and ecology of epiphytic bryophytes and lichens in dry evergreen Forest of Guyana. Journal of Tropical Ecology, vol. 5, no. 2, p. 131-150. http://dx.doi.org/10.1017/S0266467400003400.
http://dx.doi.org/10.1017/S0266467400003...
, Gradstein et al. (2001)Gradstein, SR., Churchill, SP. and Salazar, NA., 2001. Guide to the Bryophytes of Tropical America. Memoirs of the New York Botanical Garden, vol. 86, p. 1-577., Alvarenga and Pôrto (2007)Alvarenga, LDP. and PÔRTO, KC., 2007. Patch size and isolation effects on epiphytic and epiphyllous bryophytes in the fragmented Brazilian Atlantic forest. Biological Conservation, vol. 34, p. 415-427., Gradstein and Ilkiu-Borges (2009)Gradstein, SR. and Ilkiu-Borges, AL., 2009. Guide to the Plants of Central French Guiana. Part 4. Liverworts and Hornworts. Memoirs of the New York Botanical Garden. vol. 76, p. 1-140., Silva and Pôrto (2009Silva, MPP. and Pôrto, KC., 2009. Effect of fragmentation on the community structure of epixylic bryophytes in Atlantic Forest remnants in the Northeast of Brazil. Biodiversity and Conservation, vol. 18, no. 2, p. 317-337. http://dx.doi.org/10.1007/s10531-008-9487-0.
http://dx.doi.org/10.1007/s10531-008-948...
, 2013Silva, MPP. and Pôrto, KC., 2013. Bryophyte communities along horizontal and vertical gradients in a human-modified Atlantic Forest remnant. Botany, vol. 91, no. 3, p. 155-166. http://dx.doi.org/10.1139/cjb-2012-0194.
http://dx.doi.org/10.1139/cjb-2012-0194...
), Alvarenga et al. (2010)Alvarenga, LDP., Pôrto, KC. and Oliveira, JRPM., 2010. Habitat loss effects on spatial distribution of non-vascular epiphytes in a Brazilian Atlantic forest. Biodiversity and Conservation, vol. 19, no. 3, p. 619-635. http://dx.doi.org/10.1007/s10531-009-9723-2.
http://dx.doi.org/10.1007/s10531-009-972...
, Oliveira et al. (2011)Oliveira, JRPM., Pôrto, KC. and Silva, MPP., 2011. Richness preservation in a fragmented landscape: a study of epiphytic bryophytes in an Atlantic forest remnant in Northeast Brazil. Journal of Bryology, vol. 33, no. 4, p. 279-290. http://dx.doi.org/10.1179/1743282011Y.0000000017.
http://dx.doi.org/10.1179/1743282011Y.00...
and Santos et al. (2011)Santos, ND., Costa, DP., Kinoshita, LS. and Shepherd, GJ., 2011. Aspectos brioflorísticos e fitogeográficos de duas formações costeiras de Floresta Atlântica da Serra do Mar, Ubatuba/SP, Brasil. Biota Neotropica, vol. 11, no. 2, p. 425-438. http://dx.doi.org/10.1590/S1676-06032011000200040.
http://dx.doi.org/10.1590/S1676-06032011...
.
To check whether there is significant difference in the guilds of tolerance (Es, Esh and Gen) among different forest fragments was also used a chi-squared test.
3 Results
There were recorded 1,595 occurrences of bryophytes, totalling 77 species, belonging to 45 genera and 18 families (Table 1). The species accumulation curve (Figure 2) did not stabilise.
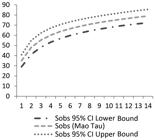
Species accumulation curve generated for the 14 fragments analysed in the region of Volta Grande do Xingu, Pará state, Brazil.
Liverwort were represented by 48 species (62%), 26 genera and six families, and therefore, with highest species richness compared to mosses (38%). Lejeuneaceae was the most representative family with 40 species (52%). Mosses accounted for 29 species (38%), 19 genera and 12 families, among which stood out Calymperaceae (7 spp.), Fissidentaceae (4 spp.), Pilotrichaceae (4 spp.), and Sematophyllaceae (4 spp.).
The richest genera among liverworts were Lejeunea (7 spp.), Cheilolejeunea (5 spp.) and Ceratolejeunea (4 spp.), and among mosses were Fissidens (4 spp.), Calymperes (3 spp.) and Syrrhopodon (3 spp.). The most occurring species were Archilejeunea parviflora, Ceratolejeunea coarina, Cololejeunea camilli, C. subcardiocarpa, Stictolejeunea squamata, Plagiochila disticha, P. subplana, Fissidens guianensis, Calymperes erosum, Callicostella pallida, Isopterygium tenerum, Pilosium chlorophyllum, and Pelekium scabrosulum.
Regarding the frequency of the species, 27 (35%) were classified as rare, 23 (30%) common, and 27 (35%) constant. Among the species classified as rare, the following species stood out by occurring once in this study, Calypogeia miquelii, Cheilolejeunea aneogyna, Harpalejeunea oxyphylla, Lejeunea setiloba, Marchesinia brachiata, Radula javanica, Leucobryum martianum, Lepidopilum scabrisetum, and L. surinamense.
In the similarity analysis (Figure 3), F7 and F14 showed the lowest degree of similarity to the further fragments, which resulted in their location at different branches. They also presented the lowest species richness, compared to all other studied fragments. In F7 were found bryophytes colonizing pieces of plastics in the area of the fragment edge.
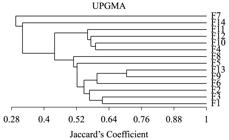
Floristic similarity between fragments in the region of Volta Grande do Xingu, Pará state, Brazil.
The cladogram of similarity also showed the formation of two big groups of fragments, one containing four fragments (F4, F10, F12, and F11) and the other more heterogeneous, consisting of eight fragments (F1, F3, F2, F6, F9, F13, F5 and F8). In the first group, F4 and F10 were more similar than F11 and F12. In the second group, F5 and F8 were less similar than the grouping formed by F1, F3 and F2, and F6, F9 and F13.
The chi-squared test indicated the existence of significant differences (p = 0. 0151) in the number of species of bryophytes among the studied areas (Figure 4).
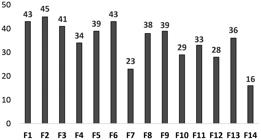
Species richness of bryophytes in different forest fragments studied in the region of Volta Grande do Xingu, Pará state, Brazil.
The listed species were generalists in their majority (48%), followed by shade epiphytes (30%) and sun epiphytes (21%), one species has not been classified.
In the result of chi-square applied to the guilds of tolerance (Table 2), no significant differences were observed (p = 0.9907) between the categories analyzed.
Distribution of the number of different species of bryophytes by the guilds in the studied fragments in the region of Volta Grande do Xingu, Pará state, Brazil, according to the categories: generalist, epiphytes of sun and shade.
4 Discussion
The species richness found in the study area (77 spp.) included two species to the list of bryophytes reported by Costa (2014)Costa, DP., 2014. Briófitas. In Jardim Botânico do Rio de Janeiro - JBRJ. Lista de espécies da flora do Brasil. Rio de Janeiro. Available from: <http://floradobrasil.jbrj.gov.br>. Access in: July 2014.
http://floradobrasil.jbrj.gov.br...
for the Pará State, which happens to be of 333 species. In the study area, the species accumulation curve was not stabilized, but according to Schilling and Batista (2008)Schilling, AC. and Batista, JLF., 2008. Curva de acumulação de espécies e suficiência amostral em florestas tropicais. Revista Brasileira de Botanica. Brazilian Journal of Botany, vol. 31, no. 1, p. 179-187. http://dx.doi.org/10.1590/S0100-84042008000100016.
http://dx.doi.org/10.1590/S0100-84042008...
the use of this method in tropical forest is controversial, due to the high richness inherent to this environment. In accordance with Zartman and Nascimento (2006)Zartman, CE. and Nascimento, H., 2006. Are patch-tracking metacommunities dispersal limited? Inferences from abundance-occupancy patterns of epiphylls in Amazonian forest fragments. Biological Conservation, vol. 127, no. 1, p. 46-54. http://dx.doi.org/10.1016/j.biocon.2005.07.012.
http://dx.doi.org/10.1016/j.biocon.2005....
and Zartman and Shaw (2006)Zartman, CE. and Shaw, AJ., 2006. Metapopulation extinction thresholds in rain forest remnants. American Naturalist, vol. 167, no. 2, p. 177-189. http://dx.doi.org/10.1086/499376. PMid:16670979.
http://dx.doi.org/10.1086/499376...
, the fragmentation causes loss of richness and diversity of bryophytes. We believe that the sampling on the great curve of the Xingu River was sufficient to represent the bryoflora of the area, since it is not possible to appoint the necessary sampling effort to achieve the richness of an area (Vanzolini, 1992Vanzolini, PE., 1992. Paleoclimas e especiação em animais da américa do sul tropical. Estudos Avançados, vol. 6, no. 15, p. 1-25. http://dx.doi.org/10.1590/S0103-40141992000200003.
http://dx.doi.org/10.1590/S0103-40141992...
), and because the richness found corresponds to 23.3% of the bryoflora reported for Pará state.
Lejeuneaceae was the most representative family in this study and this is in agreement with other surveys carried out in the Amazon (Gradstein and Costa, 2003Gradstein, SR. and Costa, DP., 2003. The Hepaticae and Anthocerotae of Brazil. Memoirs of the New York Botanical Garden, vol. 87, p. 1-336.; Ilkiu-Borges et al., 2004Ilkiu-Borges, AL., Tavares, ACC. and Lisboa, RCL., 2004. Briófitas da Ilha de Germoplasma, reservatório de Tucuruí, Pará, Brasil. Acta Botanica Brasilica, vol. 18, no. 3, p. 689-692. http://dx.doi.org/10.1590/S0102-33062004000300026.
http://dx.doi.org/10.1590/S0102-33062004...
, 2009Ilkiu-Borges, AL., LISBOA, RCL. and MORAES, ENR., 2009. Avanços no conhecimento da brioflora. In LISBOA, PLB. (Ed.). Caxiuanã: desafios para a conservação de uma Floresta Nacional na Amazônia. Pará: Museu Paraense Emílio Goeldi. p. 313-330.; Lisboa and Tavares, 2008Lisboa, RCL. and TAVARES, ACC., 2008. Briófitas de Santarém Novo, Pará. In JARDIM, MAG. and ZOGHBI, MGB. (Eds.). Flora Resex Chocoaré-Mato Grosso (PA): diversidades e usos. Museu Paraense Emílio Goeldi. p. 51-61.; Alvarenga and Lisboa, 2009Alvarenga, LDP. and Lisboa, RCL., 2009. Contribuição para o conhecimento da taxonomia, ecologia e fitogeografia de Briófitas da Amazônia Oriental. Acta Amazonica, vol. 39, no. 3, p. 495-504. http://dx.doi.org/10.1590/S0044-59672009000300003.
http://dx.doi.org/10.1590/S0044-59672009...
; Moura et al., 2013Moura, OS., Ilkiu-Borges, AL. and Brito, ES., 2013. Brioflora (Bryophyta e Marchantiophyta) da Ilha do Combu, Belém, PA, Brasil. Hoehnea., vol. 40, no. 1, p. 143-165. http://dx.doi.org/10.1590/S2236-89062013000100008.
http://dx.doi.org/10.1590/S2236-89062013...
; Brito and Ilkiu-Borges, 2013Brito, ES. and Ilkiu-Borges, AL., 2013. Bryoflora of the municipalities of Soure and Cachoeira do Arari, on Marajó Island, in the state of Pará, Brazil. Acta Botanica Brasilica, vol. 27, no. 1, p. 124-141. http://dx.doi.org/10.1590/S0102-33062013000100013.
http://dx.doi.org/10.1590/S0102-33062013...
; Tavares-Martins et al., 2014Tavares-Martins, ACC., Lisboa, RCL. and Costa, DP., 2014. Bryophyte flora in upland forests at different successional stages and in the various strata of host trees in northeastern Pará, Brazil. Acta Botanica Brasilica, vol. 28, no. 1, p. 46-58. http://dx.doi.org/10.1590/S0102-33062014000100005.
http://dx.doi.org/10.1590/S0102-33062014...
). The results confirmed it as one of the richest families in the Neotropics, representing approximately 70% of the liverwort in tropical forests (Gradstein et al., 2001Gradstein, SR., Churchill, SP. and Salazar, NA., 2001. Guide to the Bryophytes of Tropical America. Memoirs of the New York Botanical Garden, vol. 86, p. 1-577.).
The higher richness of Calymperaceae, Fissidentaceae, Pilotrichaceae, and Sematophyllaceae was expected and can be explained by their high incidence among the richest families in the Neotropics (Gradstein et al., 2001Gradstein, SR., Churchill, SP. and Salazar, NA., 2001. Guide to the Bryophytes of Tropical America. Memoirs of the New York Botanical Garden, vol. 86, p. 1-577.). Fissidentaceae is mainly found in tropical regions, but its richness decreases with higher latitude (Pursell, 2007Pursell, AR., 2007. Fissidentaceae. Flora Neotropica Monograph, vol. 1, p. 278.).
The genera Lejeunea, Cheilolejeunea, Ceratolejeunea, Fissidens, Calymperes and Syrrhopodon are often the most representative in Pará (Santos and Lisboa, 2008Santos, RCP. and LISBOA, R., 2008. Musgos (Bryophyta) da Microrregião do Salgado Paraense e sua utilização como possíveis indicadores de ambientes perturbados. Rodriguésia, vol. 59, no. 2, p. 361-368.; Moraes and Lisboa, 2009Moraes, EN. and Lisboa, RCL., 2009. Diversidade, taxonomia e distribuição por estados brasileiros das famílias Bartramiaceae, Brachytheciaceae, Bryaceae, Calymperaceae, Fissidentaceae, Hypnaceae e Leucobryaceae (Bryophyta) da Estação Científica Ferreira Penna, Caxiuanã, Pará, Brasil. Acta Amazonica, vol. 39, no. 4, p. 773-792. http://dx.doi.org/10.1590/S0044-59672009000400006.
http://dx.doi.org/10.1590/S0044-59672009...
; Brito and Ilkiu-Borges, 2013Brito, ES. and Ilkiu-Borges, AL., 2013. Bryoflora of the municipalities of Soure and Cachoeira do Arari, on Marajó Island, in the state of Pará, Brazil. Acta Botanica Brasilica, vol. 27, no. 1, p. 124-141. http://dx.doi.org/10.1590/S0102-33062013000100013.
http://dx.doi.org/10.1590/S0102-33062013...
; Moura et al., 2013Moura, OS., Ilkiu-Borges, AL. and Brito, ES., 2013. Brioflora (Bryophyta e Marchantiophyta) da Ilha do Combu, Belém, PA, Brasil. Hoehnea., vol. 40, no. 1, p. 143-165. http://dx.doi.org/10.1590/S2236-89062013000100008.
http://dx.doi.org/10.1590/S2236-89062013...
), and therefore this result was also predictable.
Among the most occurring species, Calymperes erosum is considered quite tolerant, and may be potentially indicative of disturbed habitats (Santos and Lisboa, 2008Santos, RCP. and LISBOA, R., 2008. Musgos (Bryophyta) da Microrregião do Salgado Paraense e sua utilização como possíveis indicadores de ambientes perturbados. Rodriguésia, vol. 59, no. 2, p. 361-368.), other species such as Octoblepharum albidum, Taxitelium planum, Sematophyllum subsimplex and Calymperes palisotii which also occurred in the region of the great curve of the Xingu River were considered as indicators of disturbed habitats by Lisboa and Ilkiu-Borges (1995)Lisboa, RCL. and ILKIU-BORGES, AL., 1995. Diversidade das briófitas de Belém (PA) e seu potencial como indicadoras de poluição. Boletim do Museu Paraense Emilio Goeldi. Série Ciências Naturais, vol. 11, p. 199-225..
Studies in tropical forests pointed to a greater richness of rare species (Silva and Pôrto, 2007SILVA, MPP. and PÔRTO, KC., 2007. Composição e riqueza de briófitas epíxilas em fragmentos florestais da Estação Ecológica de Murici, Alagoas. Revista Brasileira de Biologia, vol. 5, p. 243-245.; Garcia, 2012Garcia, ET., 2012. Briófitas (Bryophyta e Marchantiophyta) de Remanescentes Florestais no Reservatório de Tucuruí, Pará, Brasil. Pará: Universidade Federal Rural da Amazônia, Museu Paraense Emilio Goeldi. 87 p. Masters Dissertation.). Therefore, the equal rates of rare and constant species is unusual and indicates a change in the establishment of bryophytes community in the region of the great curve of the Xingu River. The strategies of dispersal and establishment of bryophytes are associated with spatial patterns of local richness and environmental conditions (favorable and stable), thus, many species have small populations and heavily dependent on interactions with neighboring populations (Husband and Barrett, 1996Husband, BC. and Barrett, SCH., 1996. Metapopulation perspective in plant population biology. Journal of Ecology, vol. 84, no. 3, p. 461-469. http://dx.doi.org/10.2307/2261207.
http://dx.doi.org/10.2307/2261207...
; Glime, 2007Glime, JM., 2007. Economic and ethnic uses of bryophytes. In Editorial Committee (Ed.). Flora of North America North of Mexico: bryophyta, part 1. New York: Oxford Universit Press. p. 14-41. vol. 27.; Mota de Oliveira et al., 2009Mota de Oliveira, S., ter Steege, H., Cornelissen, JHC. and Robbert Gradstein, S., 2009. Niche assembly of epiphytic bryophyte communities in the Guianas: a regional approach. Journal of Biogeography, vol. 36, no. 11, p. 2076-2084. http://dx.doi.org/10.1111/j.1365-2699.2009.02144.x.
http://dx.doi.org/10.1111/j.1365-2699.20...
).
In this study, the mosses Leucobryum martianum, Lepidopilum surinamense and L. scabrisetum as well as the liverworts Cheilolejeunea aneogyna, Harpalejeunea oxyphylla and Radula javanica were considered rare.
Leucobryum martianum, Lepidopilum surinamense and L. scabrisetum are characteristic epiphytes of humid tropical forest (Lisboa and Ilkiu-Borges, 1996Lisboa, RCL. and ILKIU-BORGES, AL., 1996. Briófitas da Serra de Carajás e sua possível utilização como indicadoras de metais. Boletim do Museu Paraense Emilio Goeldi. Série Ciências Naturais, vol. 12, p. 161-181.; Santos and Lisboa, 2008Santos, RCP. and LISBOA, R., 2008. Musgos (Bryophyta) da Microrregião do Salgado Paraense e sua utilização como possíveis indicadores de ambientes perturbados. Rodriguésia, vol. 59, no. 2, p. 361-368.; Moraes and Lisboa, 2009Moraes, EN. and Lisboa, RCL., 2009. Diversidade, taxonomia e distribuição por estados brasileiros das famílias Bartramiaceae, Brachytheciaceae, Bryaceae, Calymperaceae, Fissidentaceae, Hypnaceae e Leucobryaceae (Bryophyta) da Estação Científica Ferreira Penna, Caxiuanã, Pará, Brasil. Acta Amazonica, vol. 39, no. 4, p. 773-792. http://dx.doi.org/10.1590/S0044-59672009000400006.
http://dx.doi.org/10.1590/S0044-59672009...
; Moura et al., 2013Moura, OS., Ilkiu-Borges, AL. and Brito, ES., 2013. Brioflora (Bryophyta e Marchantiophyta) da Ilha do Combu, Belém, PA, Brasil. Hoehnea., vol. 40, no. 1, p. 143-165. http://dx.doi.org/10.1590/S2236-89062013000100008.
http://dx.doi.org/10.1590/S2236-89062013...
), and the later one seems to be confined to this type of environment (Florschütz-de Waard, 1986Florschütz-De Waard, J., 1986. Musci (Part II). In STOFFERS, AL. and LINDEMAN, JC. (Eds.). Flora of Suriname. Leiden: Brill. p. 274-361.). In Pará, only L. surinamense was found in canga vegetation, an open and dry environment (Moraes and Lisboa, 2006Moraes, ENR. and LISBOA, RCL., 2006. Musgos (Bryophyta) da Serra dos Carajás, Estado do Pará, Brasil. Boletim do Museu Paraense Emilio Goeldi. Série Ciências Naturais, vol. 1, p. 39-68.). Leucobryum martianum was also considered abundant in the Amazon, forming large mats on the floor of terra firme forest (Yano, 1992Yano, O., 1992. Leucobryaceae (Bryopsida) do Brasil. São Paulo: Universidade de São Paulo. 318 p. Doctoral Thesis.),
Cheilolejeunea aneogyna, Harpalejeunea oxyphylla and Radula javanica are common species in lowland tropical forest, and the two former species can also be found in submontane forest (Gradstein and Costa, 2003Gradstein, SR. and Costa, DP., 2003. The Hepaticae and Anthocerotae of Brazil. Memoirs of the New York Botanical Garden, vol. 87, p. 1-336.). In Pará, Cheilolejeunea aneogyna was registered in both primary and secondary forests (Garcia et al., 2014Garcia, ET., Ilkiu-Borges, AL. and Tavares-Martins, ACC., 2014. Brioflora de Florestas de terra firme na Área de Proteção Ambiental do Lago de Tucuruí PA, Brasil. Hoehnea, vol. 41, no. 4, p. 499-514. http://dx.doi.org/10.1590/2236-8906-10/2013.
http://dx.doi.org/10.1590/2236-8906-10/2...
; Moura et al., 2013Moura, OS., Ilkiu-Borges, AL. and Brito, ES., 2013. Brioflora (Bryophyta e Marchantiophyta) da Ilha do Combu, Belém, PA, Brasil. Hoehnea., vol. 40, no. 1, p. 143-165. http://dx.doi.org/10.1590/S2236-89062013000100008.
http://dx.doi.org/10.1590/S2236-89062013...
; Tavares-Martins et al., 2014Tavares-Martins, ACC., Lisboa, RCL. and Costa, DP., 2014. Bryophyte flora in upland forests at different successional stages and in the various strata of host trees in northeastern Pará, Brazil. Acta Botanica Brasilica, vol. 28, no. 1, p. 46-58. http://dx.doi.org/10.1590/S0102-33062014000100005.
http://dx.doi.org/10.1590/S0102-33062014...
). Tavares-Martins et al. (2014)Tavares-Martins, ACC., Lisboa, RCL. and Costa, DP., 2014. Bryophyte flora in upland forests at different successional stages and in the various strata of host trees in northeastern Pará, Brazil. Acta Botanica Brasilica, vol. 28, no. 1, p. 46-58. http://dx.doi.org/10.1590/S0102-33062014000100005.
http://dx.doi.org/10.1590/S0102-33062014...
registered H. oxyphylla and R. javanica exclusively in canopy of terra firme forest and emphasized that these species are typical of open environments.
According to Söderström and During (2005)Söderström, L. and During, HJ., 2005. Bryophyte rarity viewed from the perspectives of life history strategy and metapopulation dynamics. Journal of Bryology, vol. 27, no. 3, p. 261-268. http://dx.doi.org/10.1179/174328205X70010.
http://dx.doi.org/10.1179/174328205X7001...
, there are various types of species rarity and a species can be rare in different scales. Since the species above mentioned were collected in different areas and environments of the Pará state, also being considered common, the rarity of these species in the study area can be related to the fragmentation and the isolation of the fragment, which decrease the rates of species colonization (Zartman and Shaw, 2006Zartman, CE. and Shaw, AJ., 2006. Metapopulation extinction thresholds in rain forest remnants. American Naturalist, vol. 167, no. 2, p. 177-189. http://dx.doi.org/10.1086/499376. PMid:16670979.
http://dx.doi.org/10.1086/499376...
).
The less similar fragments, F7 and F14, were located very close to the access roads. Although not visible during the fragment selection for this study, the canopy of the two fragments was more open in relation to the others, as well as possessing lower tree species richness and signs of disturbance inside, such as cut trees and plastic waste.
The conservation status of the fragments was the factor that exerted the greatest influence on the similarity between areas in the region of the great curve of the Xingu River. The fragments F9 and F13 were those who had a higher degree of similarity, followed by F3 and F1 and by F10 and F4. These showed structures with similar canopies (more closed), without signs of disturbance inside and greater presence of tree species.
The species composition was also influenced by the conservation status of the fragments, because as stated Zartman (2003)Zartman, CE., 2003. Habitat fragmentation impacts on epiphyllous bryophyte communities in central Amazonia. Ecology, vol. 84, no. 4, p. 948-954. http://dx.doi.org/10.1890/0012-9658(2003)084[0948:HFIOEB]2.0.CO;2.
http://dx.doi.org/10.1890/0012-9658(2003...
and Zartman and Nascimento (2006)Zartman, CE. and Nascimento, H., 2006. Are patch-tracking metacommunities dispersal limited? Inferences from abundance-occupancy patterns of epiphylls in Amazonian forest fragments. Biological Conservation, vol. 127, no. 1, p. 46-54. http://dx.doi.org/10.1016/j.biocon.2005.07.012.
http://dx.doi.org/10.1016/j.biocon.2005....
, the maintenance of communities depends on neighboring populations, which may be present in near fragments.
The fragments with the higher species richness were F1, F2, F3, and F6, which possibly generated the significant difference observed. They shared physiognomic characteristics, present enclosed canopy and few clearings and, therefore, were more humid. According to Oliveira et al. (2011)Oliveira, JRPM., Pôrto, KC. and Silva, MPP., 2011. Richness preservation in a fragmented landscape: a study of epiphytic bryophytes in an Atlantic forest remnant in Northeast Brazil. Journal of Bryology, vol. 33, no. 4, p. 279-290. http://dx.doi.org/10.1179/1743282011Y.0000000017.
http://dx.doi.org/10.1179/1743282011Y.00...
, the best physionomically structured fragments harbor a rich bryoflora, because they have a higher number of trees, with larger diameters and height, which are important characteristics for their maintenance. The low-light environments and with more humidity are, according to Richards (1984)Richards, PW., 1984. The ecology of tropical forest bryophytes. In SCHUSTER, RM. (Ed.). New Manual of Bryology. Nichinan: Hattori Botanical Laboratory. p. 1233-1269. vol. 2., the favorable microclimate for the maintenance of the bryoflora.
Regarding to the light tolerance guilds, the result was similar to that found by Silva and Pôrto (2009Silva, MPP. and Pôrto, KC., 2009. Effect of fragmentation on the community structure of epixylic bryophytes in Atlantic Forest remnants in the Northeast of Brazil. Biodiversity and Conservation, vol. 18, no. 2, p. 317-337. http://dx.doi.org/10.1007/s10531-008-9487-0.
http://dx.doi.org/10.1007/s10531-008-948...
, 2010Silva, MPP. and Pôrto, KC., 2010. Spatial structure of bryophyte communities along an edge-interior gradient in an Atlantic Forest remnant in Northeast Brazil. Journal of Bryology, vol. 32, no. 2, p. 101-112. http://dx.doi.org/10.1179/037366810X12578498136110.
http://dx.doi.org/10.1179/037366810X1257...
, 2013Silva, MPP. and Pôrto, KC., 2013. Bryophyte communities along horizontal and vertical gradients in a human-modified Atlantic Forest remnant. Botany, vol. 91, no. 3, p. 155-166. http://dx.doi.org/10.1139/cjb-2012-0194.
http://dx.doi.org/10.1139/cjb-2012-0194...
) and Santos et al. (2011)Santos, ND., Costa, DP., Kinoshita, LS. and Shepherd, GJ., 2011. Aspectos brioflorísticos e fitogeográficos de duas formações costeiras de Floresta Atlântica da Serra do Mar, Ubatuba/SP, Brasil. Biota Neotropica, vol. 11, no. 2, p. 425-438. http://dx.doi.org/10.1590/S1676-06032011000200040.
http://dx.doi.org/10.1590/S1676-06032011...
, conducted in lowland forests in Atlantic Forest. In a study conducted in submontane forest, in Bolivia, Acebey et al. (2003)Acebey, A., Gradstein, SR. and Krömer, T., 2003. Species richness and habitat diversification of bryophytes in submontane rain forest and fallows of Bolivia. Journal of Tropical Ecology, vol. 19, no. 1, p. 9-18. http://dx.doi.org/10.1017/S026646740300302X.
http://dx.doi.org/10.1017/S0266467403003...
realized that generalist species are apparently indifferent to forest destruction, while those with smaller niches (experts) seem to be less likely to survive the deforestation, as observed in this study where there was predominance of generalists. However, the large proportion of specialist (sun/share epiphytes), indicates that the fragments still have conditions for their establishment, mainly by their physiognomic conditions.
The species can vary greatly in their susceptibility to fragmentation due to their different dispersion abilities, persistence and establishment in fragmented landscapes (Kolb and Diekmann, 2004Kolb, A. and Diekmann, M., 2004. Effect of life-history traists of responses of plant species to forest fragmentation. Conservation Biology, vol. 19, no. 3, p. 929-938. http://dx.doi.org/10.1111/j.1523-1739.2005.00065.x.
http://dx.doi.org/10.1111/j.1523-1739.20...
).
The fragments studied in the region of the great curve of the Xingu River revealed a rich bryoflora, which represents approximately 25% of species registered in the state of Pará (318 spp.), especially considering the fact that this study included only one type of forest formation. The results showed that predominant families were like those typically found in surveys in tropical forests, but the standard of rarity found here did not follow the pattern observed for both bryophytes and other groups of plants in tropical forests, probably due to historical fragmentation and disturbance in the area. The richness and species composition were determined mainly by the physiognomic characteristics of the studied areas.
Acknowledgements
The authors are grateful to the Programa de Pós-graduação em Ciências Biológicas – Botânica Tropical (UFRA/MPEG) for the infrastructure; to the Norte Energia S.A. for allowing access to the study area and the logistics for conducting the collections; to the Projeto Salvamento e Aproveitamento Científico da Flora da UHE Belo Monte, which is part of the Plano Básico Ambiental of Norte Energia S.A., and object of the agreement between MPEG/FADESP/Biota Projetos e Consultoria Ambiental Ltda. for the logistic support during the collections; to CAPES for the concession of the master scholarship for the first author.
-
(With 4 figures)
References
- Acebey, A., Gradstein, SR. and Krömer, T., 2003. Species richness and habitat diversification of bryophytes in submontane rain forest and fallows of Bolivia. Journal of Tropical Ecology, vol. 19, no. 1, p. 9-18. http://dx.doi.org/10.1017/S026646740300302X
» http://dx.doi.org/10.1017/S026646740300302X - Alvarenga, LDP. and Lisboa, RCL., 2009. Contribuição para o conhecimento da taxonomia, ecologia e fitogeografia de Briófitas da Amazônia Oriental. Acta Amazonica, vol. 39, no. 3, p. 495-504. http://dx.doi.org/10.1590/S0044-59672009000300003
» http://dx.doi.org/10.1590/S0044-59672009000300003 - Alvarenga, LDP. and PÔRTO, KC., 2007. Patch size and isolation effects on epiphytic and epiphyllous bryophytes in the fragmented Brazilian Atlantic forest. Biological Conservation, vol. 34, p. 415-427.
- Alvarenga, LDP., Pôrto, KC. and Oliveira, JRPM., 2010. Habitat loss effects on spatial distribution of non-vascular epiphytes in a Brazilian Atlantic forest. Biodiversity and Conservation, vol. 19, no. 3, p. 619-635. http://dx.doi.org/10.1007/s10531-009-9723-2
» http://dx.doi.org/10.1007/s10531-009-9723-2 - Ayres, M., AYRES JÚNIOR, M., Ayres, DL. and Santos, AAS., 2007. BIOESTAT: aplicações estatísticas nas áreas das ciências bio-médicas. Belém: Instituto de Desenvolvimento Sustentável Mamirauá. 364 p.
- Brito, ES. and Ilkiu-Borges, AL., 2013. Bryoflora of the municipalities of Soure and Cachoeira do Arari, on Marajó Island, in the state of Pará, Brazil. Acta Botanica Brasilica, vol. 27, no. 1, p. 124-141. http://dx.doi.org/10.1590/S0102-33062013000100013
» http://dx.doi.org/10.1590/S0102-33062013000100013 - Colwell, RK., 2009. EstimateS: Statistical estimation of species richness and shared species from samples. Storrs: University of Connecticut. Available from: <http://viceroy.eeb.uconn.edu/EstimateS>. Access in: Aug. 2012.
» http://viceroy.eeb.uconn.edu/EstimateS - COLWELL, RK., Mao, CX. and Chang, J., 2004. Interpolating, extrapolating, and comparing incidence-based species accumulation curves. Ecology, vol. 85, no. 10, p. 2717-2727. http://dx.doi.org/10.1890/03-0557
» http://dx.doi.org/10.1890/03-0557 - Cornelissen, JH. and TER STEEGE, H., 1989. Distribuition and ecology of epiphytic bryophytes and lichens in dry evergreen Forest of Guyana. Journal of Tropical Ecology, vol. 5, no. 2, p. 131-150. http://dx.doi.org/10.1017/S0266467400003400
» http://dx.doi.org/10.1017/S0266467400003400 - Costa, DP., 2014. Briófitas. In Jardim Botânico do Rio de Janeiro - JBRJ. Lista de espécies da flora do Brasil. Rio de Janeiro. Available from: <http://floradobrasil.jbrj.gov.br>. Access in: July 2014.
» http://floradobrasil.jbrj.gov.br - Crandall-Stotler, B., Stotler, RE. and Long, DG., 2009. Morphology and classification of the Marchantiophyta. In GOFFINET, B. and SHAW, AJ. (Eds.). Bryophyte biology. 2nd ed. Cambridge: University Press Cambridge. p. 1-54.
- Cunha, DA. and Ferreira, LV., 2012. Impacts of the Belo Monte hydroelectric dam construction on pioner vegetation formations along the Xingu River, Pará State, Brazil. Brazilian Journal of Botany, vol. 35, no. 2, p. 159-167. http://dx.doi.org/10.1590/S1806-99592012000200005
» http://dx.doi.org/10.1590/S1806-99592012000200005 - Florschütz-De Waard, J., 1986. Musci (Part II). In STOFFERS, AL. and LINDEMAN, JC. (Eds.). Flora of Suriname. Leiden: Brill. p. 274-361.
- Gama, JRV., Souza, AL., Martins, SV. and Souza, DR., 2005. Comparação entre florestas de várzea e de terra firme do estado do Pará. Revista Árvore, vol. 29, no. 4, p. 607-616. http://dx.doi.org/10.1590/S0100-67622005000400013
» http://dx.doi.org/10.1590/S0100-67622005000400013 - Garcia, ET., 2012. Briófitas (Bryophyta e Marchantiophyta) de Remanescentes Florestais no Reservatório de Tucuruí, Pará, Brasil. Pará: Universidade Federal Rural da Amazônia, Museu Paraense Emilio Goeldi. 87 p. Masters Dissertation.
- Garcia, ET., Ilkiu-Borges, AL. and Tavares-Martins, ACC., 2014. Brioflora de Florestas de terra firme na Área de Proteção Ambiental do Lago de Tucuruí PA, Brasil. Hoehnea, vol. 41, no. 4, p. 499-514. http://dx.doi.org/10.1590/2236-8906-10/2013
» http://dx.doi.org/10.1590/2236-8906-10/2013 - Glime, JM., 2007. Economic and ethnic uses of bryophytes. In Editorial Committee (Ed.). Flora of North America North of Mexico: bryophyta, part 1. New York: Oxford Universit Press. p. 14-41. vol. 27.
- Goffinet, B., Buck, WR. and Shaw, AJ., 2009. Morphology, anatomy, and classification of the Bryophyta. In GOFFINET, B. and SHAW, AJ. (Eds.). Bryophyte biology. 2nd ed. Cambridge: University Press Cambridge. p. 55-138.
- Gradstein, SR. and Costa, DP., 2003. The Hepaticae and Anthocerotae of Brazil. Memoirs of the New York Botanical Garden, vol. 87, p. 1-336.
- Gradstein, SR. and Ilkiu-Borges, AL., 2009. Guide to the Plants of Central French Guiana. Part 4. Liverworts and Hornworts. Memoirs of the New York Botanical Garden. vol. 76, p. 1-140.
- Gradstein, SR. and Pócs, T., 1989. Bryophytes. In LIETH, H. and WERGER, MJA. (Eds.). Tropical rain forest ecosystems. Amsterdam: Elseiver Science Publishers. p. 311-325. http://dx.doi.org/10.1016/B978-0-444-42755-7.50022-5
» http://dx.doi.org/10.1016/B978-0-444-42755-7.50022-5 - Gradstein, SR., Churchill, SP. and Salazar, NA., 2001. Guide to the Bryophytes of Tropical America. Memoirs of the New York Botanical Garden, vol. 86, p. 1-577.
- Hair JUNIOR, JF., Anderson, RE., Tathan, RL. and Black, WC., 2006. Análise multivariada de dados. Porto Alegre: Bookman. 593 p.
- Husband, BC. and Barrett, SCH., 1996. Metapopulation perspective in plant population biology. Journal of Ecology, vol. 84, no. 3, p. 461-469. http://dx.doi.org/10.2307/2261207
» http://dx.doi.org/10.2307/2261207 - Ilkiu-Borges, AL., Macêdo, LPC.,PEREIRA, MAV. and LISBOA, RCL., 2013. Briófitas em Caxiuanã: resultados do levantamento em duas parcelas da grade do PPBIO. In LISBOA, PLB. (Ed.). Caxiuanã: paraíso ainda preservado. Pará: Museu Paraense Emilio Goeldi. p. 287-295.
- Ilkiu-Borges, AL., LISBOA, RCL. and MORAES, ENR., 2009. Avanços no conhecimento da brioflora. In LISBOA, PLB. (Ed.). Caxiuanã: desafios para a conservação de uma Floresta Nacional na Amazônia. Pará: Museu Paraense Emílio Goeldi. p. 313-330.
- Ilkiu-Borges, AL., Tavares, ACC. and Lisboa, RCL., 2004. Briófitas da Ilha de Germoplasma, reservatório de Tucuruí, Pará, Brasil. Acta Botanica Brasilica, vol. 18, no. 3, p. 689-692. http://dx.doi.org/10.1590/S0102-33062004000300026
» http://dx.doi.org/10.1590/S0102-33062004000300026 - Instituto Nacional de Meteorologia - INMET, 2012. Brasília. Available from: <http://www.inmet.gov.br>. Access in: Mar 2012.
» http://www.inmet.gov.br - Kolb, A. and Diekmann, M., 2004. Effect of life-history traists of responses of plant species to forest fragmentation. Conservation Biology, vol. 19, no. 3, p. 929-938. http://dx.doi.org/10.1111/j.1523-1739.2005.00065.x
» http://dx.doi.org/10.1111/j.1523-1739.2005.00065.x - Lisboa, RCL. and ILKIU-BORGES, AL., 1995. Diversidade das briófitas de Belém (PA) e seu potencial como indicadoras de poluição. Boletim do Museu Paraense Emilio Goeldi. Série Ciências Naturais, vol. 11, p. 199-225.
- Lisboa, RCL. and ILKIU-BORGES, AL., 1996. Briófitas da Serra de Carajás e sua possível utilização como indicadoras de metais. Boletim do Museu Paraense Emilio Goeldi. Série Ciências Naturais, vol. 12, p. 161-181.
- Lisboa, RCL. and ILKIU-BORGES, AL., 2001. Briófitas de São Luís do Tapajós, Município de Itaituba, com novas adições para o Estado do Pará. Boletim do Museu Paraense Emilio Goeldi. Série Ciências Naturais, vol. 17, p. 75-91.
- Lisboa, RCL. and TAVARES, ACC., 2008. Briófitas de Santarém Novo, Pará. In JARDIM, MAG. and ZOGHBI, MGB. (Eds.). Flora Resex Chocoaré-Mato Grosso (PA): diversidades e usos. Museu Paraense Emílio Goeldi. p. 51-61.
- Lisboa, RCL., Lima, MJL. and Maciel, UN., 1999. Musgos da Ilha de Marajó - II – Anajás, Pará, Brasil. Acta Amazonica, vol. 29, no. 2, p. 201-206. http://dx.doi.org/10.1590/1809-43921999292206
» http://dx.doi.org/10.1590/1809-43921999292206 - Mausel, P., Wu, Y., Li, Y., Moran, EF. and Brondizio, ES., 1993. Spectral identification of successional stages following deforestation in the Amazon. Geocarto International, vol. 8, no. 4, p. 61-71. http://dx.doi.org/10.1080/10106049309354430
» http://dx.doi.org/10.1080/10106049309354430 - Mittermeier, RA., Werner, T., Ayres, JM. and Fonseca, GAB., 1992. O país da megadiversidade. Campinas. Available from: <http://www.bdpa.cnptia.embrapa.br>. Access in: June 2012.
» http://www.bdpa.cnptia.embrapa.br - Moraes, EN. and Lisboa, RCL., 2009. Diversidade, taxonomia e distribuição por estados brasileiros das famílias Bartramiaceae, Brachytheciaceae, Bryaceae, Calymperaceae, Fissidentaceae, Hypnaceae e Leucobryaceae (Bryophyta) da Estação Científica Ferreira Penna, Caxiuanã, Pará, Brasil. Acta Amazonica, vol. 39, no. 4, p. 773-792. http://dx.doi.org/10.1590/S0044-59672009000400006
» http://dx.doi.org/10.1590/S0044-59672009000400006 - Moraes, ENR. and LISBOA, RCL., 2006. Musgos (Bryophyta) da Serra dos Carajás, Estado do Pará, Brasil. Boletim do Museu Paraense Emilio Goeldi. Série Ciências Naturais, vol. 1, p. 39-68.
- Mota de Oliveira, S., ter Steege, H., Cornelissen, JHC. and Robbert Gradstein, S., 2009. Niche assembly of epiphytic bryophyte communities in the Guianas: a regional approach. Journal of Biogeography, vol. 36, no. 11, p. 2076-2084. http://dx.doi.org/10.1111/j.1365-2699.2009.02144.x
» http://dx.doi.org/10.1111/j.1365-2699.2009.02144.x - Moura, OS., Ilkiu-Borges, AL. and Brito, ES., 2013. Brioflora (Bryophyta e Marchantiophyta) da Ilha do Combu, Belém, PA, Brasil. Hoehnea., vol. 40, no. 1, p. 143-165. http://dx.doi.org/10.1590/S2236-89062013000100008
» http://dx.doi.org/10.1590/S2236-89062013000100008 - Oliveira, AA. and Mori, SA., 1999. A central Amazonian terra firme forest. I. High tree species richness on poor soils. Biodiversity and Conservation, vol. 8, no. 9, p. 1219-1244. http://dx.doi.org/10.1023/A:1008908615271
» http://dx.doi.org/10.1023/A:1008908615271 - Oliveira, JRPM., Pôrto, KC. and Silva, MPP., 2011. Richness preservation in a fragmented landscape: a study of epiphytic bryophytes in an Atlantic forest remnant in Northeast Brazil. Journal of Bryology, vol. 33, no. 4, p. 279-290. http://dx.doi.org/10.1179/1743282011Y.0000000017
» http://dx.doi.org/10.1179/1743282011Y.0000000017 - Pires, JM. and Prance, GT., 1985. The vegetation types of the brazilian Amazon. In PRANCE, GT. and LOVEJOY, TE. (Eds.). Key environments: Amazonia. Oxford: Pergamon Press. p. 109-145.
- Pires, JM., 1973. Tipos de vegetação da Amazônia. Belém: Museu Goeldi. p. 179-202. Publicações Avulsas, vol. 20.
- Pursell, AR., 2007. Fissidentaceae. Flora Neotropica Monograph, vol. 1, p. 278.
- Richards, PW., 1984. The ecology of tropical forest bryophytes. In SCHUSTER, RM. (Ed.). New Manual of Bryology. Nichinan: Hattori Botanical Laboratory. p. 1233-1269. vol. 2.
- Salomão, RP., Vieira, ICG., Suemitsu, C., Rosa, NA., Almeida, SS., Amaral, DD. and Menezes, MPM., 2007. As florestas de Belo Monte na grande curva do rio Xingu, Amazônia Oriental. Boletim do Museu Paraense Emilio Goeldi. Série Ciências Naturais, vol. 2, p. 57-153.
- Santos, ND., Costa, DP., Kinoshita, LS. and Shepherd, GJ., 2011. Aspectos brioflorísticos e fitogeográficos de duas formações costeiras de Floresta Atlântica da Serra do Mar, Ubatuba/SP, Brasil. Biota Neotropica, vol. 11, no. 2, p. 425-438. http://dx.doi.org/10.1590/S1676-06032011000200040
» http://dx.doi.org/10.1590/S1676-06032011000200040 - Santos, RCP. and LISBOA, R., 2008. Musgos (Bryophyta) da Microrregião do Salgado Paraense e sua utilização como possíveis indicadores de ambientes perturbados. Rodriguésia, vol. 59, no. 2, p. 361-368.
- Schilling, AC. and Batista, JLF., 2008. Curva de acumulação de espécies e suficiência amostral em florestas tropicais. Revista Brasileira de Botanica. Brazilian Journal of Botany, vol. 31, no. 1, p. 179-187. http://dx.doi.org/10.1590/S0100-84042008000100016
» http://dx.doi.org/10.1590/S0100-84042008000100016 - SILVA, MPP. and PÔRTO, KC., 2007. Composição e riqueza de briófitas epíxilas em fragmentos florestais da Estação Ecológica de Murici, Alagoas. Revista Brasileira de Biologia, vol. 5, p. 243-245.
- Silva, MPP. and Pôrto, KC., 2009. Effect of fragmentation on the community structure of epixylic bryophytes in Atlantic Forest remnants in the Northeast of Brazil. Biodiversity and Conservation, vol. 18, no. 2, p. 317-337. http://dx.doi.org/10.1007/s10531-008-9487-0
» http://dx.doi.org/10.1007/s10531-008-9487-0 - Silva, MPP. and Pôrto, KC., 2010. Spatial structure of bryophyte communities along an edge-interior gradient in an Atlantic Forest remnant in Northeast Brazil. Journal of Bryology, vol. 32, no. 2, p. 101-112. http://dx.doi.org/10.1179/037366810X12578498136110
» http://dx.doi.org/10.1179/037366810X12578498136110 - Silva, MPP. and Pôrto, KC., 2013. Bryophyte communities along horizontal and vertical gradients in a human-modified Atlantic Forest remnant. Botany, vol. 91, no. 3, p. 155-166. http://dx.doi.org/10.1139/cjb-2012-0194
» http://dx.doi.org/10.1139/cjb-2012-0194 - Söderström, L. and During, HJ., 2005. Bryophyte rarity viewed from the perspectives of life history strategy and metapopulation dynamics. Journal of Bryology, vol. 27, no. 3, p. 261-268. http://dx.doi.org/10.1179/174328205X70010
» http://dx.doi.org/10.1179/174328205X70010 - Tavares-Martins, ACC., Lisboa, RCL. and Costa, DP., 2014. Bryophyte flora in upland forests at different successional stages and in the various strata of host trees in northeastern Pará, Brazil. Acta Botanica Brasilica, vol. 28, no. 1, p. 46-58. http://dx.doi.org/10.1590/S0102-33062014000100005
» http://dx.doi.org/10.1590/S0102-33062014000100005 - Ter Steege, H., Sabatier, D., Castellanos, H., Van Andel, T., Duivenvoorden, J., Adalardo De Oliveira, A., Ek, R., Lilwah, R., Maas, P. and Mori, S., 2000An analysis of Amazonian floristic composition, including those of the Guiana Shield. Journal of Tropical Ecology, vol. 16, no. 6, p. 801-828. http://dx.doi.org/10.1017/S0266467400001735
» http://dx.doi.org/10.1017/S0266467400001735 - Vanzolini, PE., 1992. Paleoclimas e especiação em animais da américa do sul tropical. Estudos Avançados, vol. 6, no. 15, p. 1-25. http://dx.doi.org/10.1590/S0103-40141992000200003
» http://dx.doi.org/10.1590/S0103-40141992000200003 - Yano, O., 1989. Briófitas. In FIDALGO, O. and BONONI, VLR (Eds.). Técnicas de coleta, preservação e herborização de material botânico. São Paulo: Instituto de Botânica. p. 27-30.
- Yano, O., 1992. Leucobryaceae (Bryopsida) do Brasil. São Paulo: Universidade de São Paulo. 318 p. Doctoral Thesis.
- Zartman, CE. and Nascimento, H., 2006. Are patch-tracking metacommunities dispersal limited? Inferences from abundance-occupancy patterns of epiphylls in Amazonian forest fragments. Biological Conservation, vol. 127, no. 1, p. 46-54. http://dx.doi.org/10.1016/j.biocon.2005.07.012
» http://dx.doi.org/10.1016/j.biocon.2005.07.012 - Zartman, CE. and Shaw, AJ., 2006. Metapopulation extinction thresholds in rain forest remnants. American Naturalist, vol. 167, no. 2, p. 177-189. http://dx.doi.org/10.1086/499376 PMid:16670979.
» http://dx.doi.org/10.1086/499376 - Zartman, CE., 2003. Habitat fragmentation impacts on epiphyllous bryophyte communities in central Amazonia. Ecology, vol. 84, no. 4, p. 948-954. http://dx.doi.org/10.1890/0012-9658(2003)084[0948:HFIOEB]2.0.CO;2
» http://dx.doi.org/10.1890/0012-9658(2003)084[0948:HFIOEB]2.0.CO;2
Publication Dates
-
Publication in this collection
Aug 2015
History
-
Received
15 Oct 2014 -
Accepted
31 Dec 2014




