Abstract
Southern corn leaf blight (SCLB) is one of the most important corn leaf diseases. Appropriate management strategies and the use of resistant cultivars allow disease control. Therefore, knowing the aspects related to the pathogen and the response of hosts makes it possible to design efficient strategies for selecting genotypes resistant to this disease. In this sense, the objective was to carry out the Bipolaris maydis isolate characterization, evaluating the pathogenicity in different popcorn lines and the symptoms generated in the host after inoculation. The isolate characterization consisted of the macromorphological evaluation of the colonies and the micromorphological evaluation of the conidia in the PDA medium. An experiment was carried out in a greenhouse to evaluate the pathogenicity of the isolate, using 20 inbred lines of popcorn in a randomized block design with four replicates. Inoculation was carried out by spraying leaves, with a suspension containing 1.0 x 104 conidia.ml-1 of the CF/UENF 501 isolate of B. maydis. An incidence assessment and three assessments of disease symptom severity were performed, with seven days intervals between evaluations. The morphological characterization data of the isolate were analyzed using descriptive statistics, and for disease severity, the linear regression model was applied the first-degree model. The variance analysis was performed for the linear and angular coefficients obtained for each treatment. When a difference was found, the Scott-Knott clustering algorithm at 5% significance was applied. The isolate had gray-green colonies, a cottony appearance, and an irregular shape. The lines L353, L354, and L624 show more resistance at the beginning and throughout the evaluations. The high virulence of the CF/UENF 501 isolate made it possible to differentiate the lines in terms of disease intensity and the pattern of symptoms presented.
Keywords:
leaf disease; fungus; Zea mays L
Resumo
Mancha-de-Bipolaris é uma das mais importantes doenças foliares do milho. Estratégias de manejo adequadas e o uso de cultivar resistente permitem o controle da doença, mas para tanto, conhecer os aspectos associados ao patógeno e a resposta do hospedeiro é necessário para traçar estratégias eficientes para seleção de genótipos resistentes a essa doença. Neste sentido, objetivou-se realizar a caracterização do isolado de Bipolaris maydis avaliando a patogenicidade em diferentes linhagens de milho-pipoca e os sintomas gerados no hospedeiro a partir da inoculação. A caracterização do isolado consistiu na avaliação macromorfológica das colônias e micromofológica dos conídios em meio nutritivo BDA. Para avaliação da patogenicidade do isolado foi conduzido um experimento em casa de vegetação utilizando 20 linhagens endogâmicas de milho-pipoca, com delineamento em blocos casualizados, com quatro repetições. A inoculação foi realizada por meio de pulverização em folhas, com uma suspensão contendo 1,0 x 104 conídios.ml-1 do isolado CF/UENF 501 de B. maydis. Foi realizada uma avaliação de incidência e três avaliações de severidade dos sintomas da doença, com o intervalo de sete dias para cada avaliação. Os dados da caracterização morfológica do isolado foram analisados por meio da estatística descritiva e para severidade da doença foi aplicado o modelo de regressão linear de primeiro grau. Realizou-se a análise de variância para o coeficiente linear e angular obtido para cada tratamento e quando constatada diferença aplicou-se o agrupamento de médias de Scott-knott a 5% de significância. O isolado apresentou colônias com coloração cinza esverdeado, aspecto algodonoso e forma irregular. As linhagens L353 e L354 e L624 estão entre as linhagens que apresentaram maior resistência no inicio e ao longo das avaliações. A elevada virulência do isolado CF/UENF 501 possibilitou diferenciar as linhagens quanto a intensidade da doença, bem como o padrão dos sintomas apresentados.
Palavras-chave:
doença foliar; fungo; Zea mays L
1. Introduction
The species Bipolaris maydis [(Nisikado and Miyake) Shoem] (teleomorph: Cochliobolus heterostrophus) causing southern corn leaf blight disease (SCLB) is an important pathogen that affects the Zea mays species in warm, humid regions of the world (White, 1999WHITE, D.G., 1999. Compendium of corn diseases. 3rd ed. St. Paul: American Phytopathological Society, 128 p.). The yield loss occurs due to leaf surface lesion increase, compromising the photosynthetic area of plants (Agrios, 2005AGRIOS, G.N., 2005. Foliar diseases caused by Ascomycetes and Deuteromycetes (Mitosporic Fungi). In: G. AGRIOS, ed. Plant pathology. 5th ed. Burlington: Elsevier Academic Press, pp. 466-468.).
When favorable environmental conditions provide the multiplication of pathogens, the yield of susceptible maize genotypes can be reduced by more than 70% (Ali et al., 2011ALI, F., RAHMAN, H., DURRISHAHWAR, N.F., MUNIR, M. and ULLAH, H., 2011. Genetics analysis and maturity and morphological traits under maydis leaf blight epiphytotic in maize. Journal of Agricultural and Biological Science, vol. 6, no. 8, pp. 13-19.). Utilizing chemical control and resistant genotypes are the primary forms of control (Pate et al., 2017PATE, R.T., WEATHERLY, M., HEDGES, L.B., MIDEROS, S., FELLOWS, G.M. and CARDOSO, F., 2017. 363 effects of foliar fungicide on whole plant brown midrib and floury corn varieties. Journal of Animal Science, vol. 95, no. 2, suppl. 2, pp. 176-177. http://dx.doi.org/10.2527/asasmw.2017.363.
http://dx.doi.org/10.2527/asasmw.2017.36...
). However, the high cost of fungicides and the problems they can cause to the environment point out the use of resistant cultivars as the best control disease measure. It is also noteworthy that there are no registered products for the control of B. maydis.
Some steps must be considered for developing a resistant cultivar, such as knowledge about pathogen variability and the morphological, molecular, and pathogenic characterization of the species. Pal et al. (2015)PAL, I., SINGH, V., GOGOI, R., HOODA, K.S. and BEDI, N., 2015. Characterization of Bipolaris maydis isolates of different maize cropping zones of India. Indian Phytopathology, vol. 68, no. 1, pp. 63-66. indicate the existence of variability for this pathogen, which allows finding differences in aspects associated with the morphology of the species and the infection levels.
Besides the variability present in the pathogen, the response of hosts represents an important step for assessing resistance. Resistance to B. maydis race O, more prevalent in maize, is quantitatively inherited (Burnette and White, 1985BURNETTE, D.C. and WHITE, D.G., 1985. Inheritance of resistance to Bipolaris maydis race O in crosses derived from nine resistance inbred lines of maize. Phytopathology, vol. 75, no. 11, pp. 1195-1200. http://dx.doi.org/10.1094/Phyto-75-1195.
http://dx.doi.org/10.1094/Phyto-75-1195...
; Holley and Goodman, 1989HOLLEY, R.N. and GOODMAN, M.M., 1989. New sources of resistance to Southern corn leaf blight from tropical hybrid maize derivatives. Plant Disease, vol. 73, no. 7, pp. 562-564. http://dx.doi.org/10.1094/PD-73-0562.
http://dx.doi.org/10.1094/PD-73-0562...
), allowing to find distinct reactions at the level of the disease or in the morphology of symptoms when different genotypes are considered.
The characterization of an isolate regarding its morpho-pathogenicity is of great importance for future work on developing SCLB-resistant maize cultivars. For this, knowing the morphology of the isolate and the host reaction types will help recognize the pathogen and symptoms in the resistance reaction tests of cultivars in popcorn. Thus, this study aimed to characterize the CF/UENF 501 isolate of B. maydis regarding its morphology, pathogenicity in different popcorn lines, and the symptoms generated in the host after inoculation.
2. Materials and Methods
2.1. Obtaining and characterization of the B. maydis isolate
In this study, the isolate “CF/UENF 501” of the species of B. maydis, kept at the phytosanitary clinic of UENF, was used. The isolate was collected in Campos dos Goytacazes - RJ, from a typical lesion of B. maydis in popcorn leaf. The macromorphological and micromorphological characterization of B. maydis isolates was performed in Petri dishes containing Potato Dextrose Agar – PDA medium (200g of potato, 20g of dextrose, 20g of agar, and 1000 ml of distilled water) and incubated for 14 days at 25 °C. Colony color was determined by looking at the front and back of the colony. The measurement of mycelial growth was obtained with the aid of a digital caliper, through the average of colony diameter measurements in two orthogonal axes, at 14 days of growth.
The diameter of the conidia was determined from the length and width of 30 conidia on microscope slides stained with bromophenol blue. The microscope used was an optical Nikon E-400 equipped with a digital camera.
2.2. Pathogenicity assessment
2.2.1. Plant material and conduct of the experiment
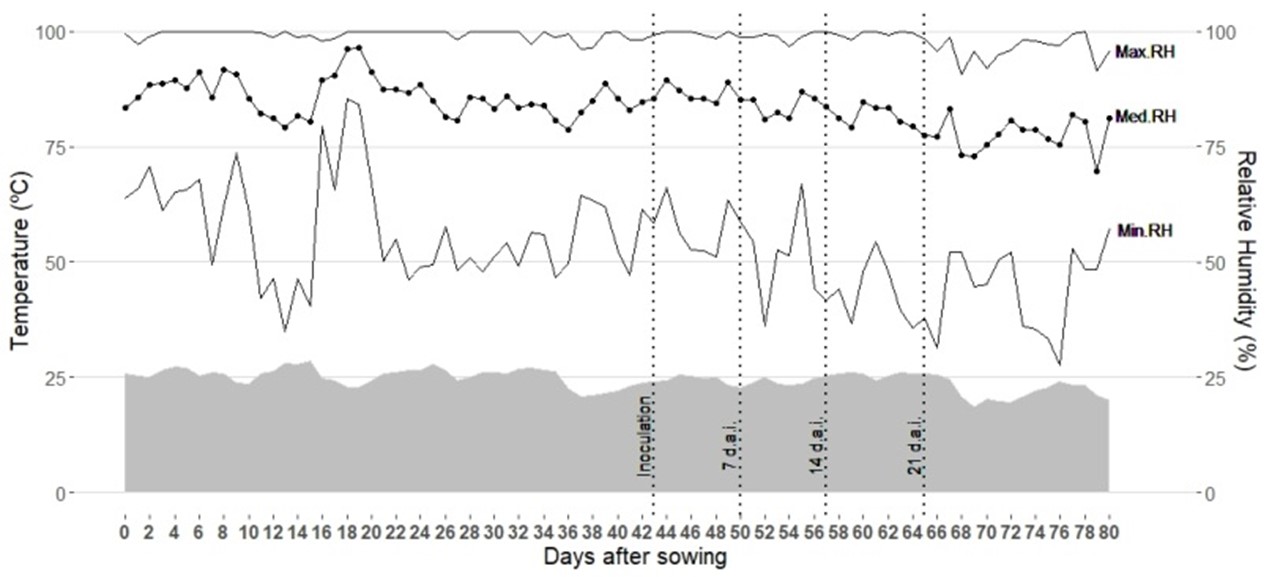
The experiment was conducted in a greenhouse at the Universidade Estadual do Norte Fluminense Darcy Ribeiro (State University of North Fluminense Darcy Ribeiro - UENF), located in Campos dos Goytacazes - RJ, Brazil. The study started with the sowing of popcorn lines on April 29, 2019, and ended on July 3, 2019, with the third and last assessment of the B. maydis severity. The average temperature (°C) and air humidity (%) during the period of the experiment were 25 °C and 85%, respectively (see Figure 1).

Variation of average daily temperature (right axis) and % relative humidity (left axis). Vertical lines indicate inoculation, severity at 7, 14, and 21 days, respectively.
The experiment was carried out in randomized blocks design with four replications and 20 treatments. The treatments studied were inbred popcorn (Zea mays L.) lines with 12 different genealogies (shown in Table 1).
The plants were cultivated in plastic pots with a volume of 5 dm3, containing a substrate based on soil: sand: manure, in a volumetric ratio of 1:1:1. Sowing was carried out with four seeds in each pot. On the twenty-seventh day after sowing, thinning was carried out, leaving only two plants per pot, constituting an experimental plot. The plants were irrigated manually, using hoses.
2.2.2. Inoculum preparation
From the pure culture of the isolate, CF/UENF 501 kept in a Petri dish containing PDA culture medium, disks of approximately 6 mm in diameter containing structures of the isolate were removed from the edge of colonies. The obtained discs were placed in the center of the Petri dish containing PDA culture medium and then incubated in a growth chamber at different temperatures with alternating light/dark for 12 h. After this period, a suspension of conidia was prepared for plant inoculation. To prepare the suspension, the surface of the culture medium, which had the fungus structure, was scraped off with a brush and placed in a container with water. The spore concentration was adjusted to 1.0 x 104 conidia.ml-1, with the aid of a hemacytometer.
2.2.3. Inoculation and symptom assessment
Inoculation was performed when the plants reached the V4 phenological stage (the fourth leaf fully expanded, with visible collar, ligule, and auricles), according to the protocol used by Santos et al. (2016)SANTOS, J.S., AMARAL JÚNIOR, A.T., VIVAS, M., VIVAS, J.M.S., KUROSAWA, R.N.F., SILVEIRA, S.F., 2016. Características culturais e patológicas de Bipolaris maydis em diferentes meios de cultura. Revista Brasileira de Milho e Sorgo, vol. 15, no. 3, pp. 461-469. http://dx.doi.org/10.18512/1980-6477/rbms.v15n3p461-469.
http://dx.doi.org/10.18512/1980-6477/rbm...
. Inoculation on the leaves was carried out by spraying the suspension containing the conidia, with total wetting of the plants to the point of runoff. 500 ml of suspension were used in each repetition, totaling 12.5 ml per plant.
Seven days after inoculation, disease symptoms were quantified, evaluating the incidence by counting leaves with signs and the severity of disease symptoms on the leaves. An incidence assessment and three severity assessments were performed at seven-day intervals. The severity was assessed on the fifth leaf, previously identified, visually and with the aid of a diagrammatic scale (James, 1971JAMES, W.C., 1971. A manual of assessment keys of plant diseases. St. Paul: American Phytopathological Society Press. Publ., no. 1458.).
2.3. Statistical analysis
Data regarding the morphological characterization of the isolate were analyzed using descriptive statistics. The mean incidence and severity, obtained by the arithmetic mean of the three assessments, were presented as a gradient of disease occurrence. For the severity data throughout the assessments, a first-degree linear regression model was applied. The linear and angular coefficients obtained for each treatment were tested for normality of residuals and homogeneity of variances by the Shapiro-Wilk (Shapiro and Wilk, 1965SHAPIRO, S.S. and WILK, M.B., 1965. An analysis of variance test for normality (complete sample). Biometrika, vol. 52, no. 3, pp. 591-611. http://dx.doi.org/10.1093/biomet/52.3-4.591.
http://dx.doi.org/10.1093/biomet/52.3-4....
), and O’Neill and Mathews (2002)O’NEILL, M.E. and MATHEWS, K.L., 2002. Levene tests of homogeneity of variance for general block and treatment designs. Biometrics, vol. 58, no. 1, pp. 216-224. http://dx.doi.org/10.1111/j.0006-341X.2002.00216.x. PMid:11890318.
http://dx.doi.org/10.1111/j.0006-341X.20...
tests at 5% significance, respectively. Given the assumptions, the variance analysis at 5% significance for the F-test was performed. When a difference was found, the means clustering was applied using the Scott-Knott method, also at 5% significance.
3. Results
3.1. Morphological characterization of the isolate
The macromorphological evaluation of isolates consisted of the description and quantification of the variables associated with the appearance of the colony. It was observed that in the PDA medium, the gray - dark greenish color was predominantly on the front of the isolate colony. Meanwhile, the black color prevailed on the back of the colony (shown in Table 2). The colony texture was cotton wool, and the presence of sectors was found in all replicates in the CF/UENF 501 isolate colony. The mycelial growth of the colony had an irregular shape with 44.25 mm in diameter after 14 days of incubation (shown in Table 2).
Macromorphological and micromorphological evaluation of the isolate “CF/UENF 501” from B. maydis.
The diameter of conidia ranged between 52.79 and 91.97 µm long and between 10.88 and 16.35 µm wide. The conidia had a fusiform shape with light brown coloration, with distoseptum and truncated hilum. The number of septa ranged between 4-10. Conidiophores usually appeared individually or in small groups, septate, straight, or sinuous, with germ tubes appearing at both ends. In this isolate, it was possible to observe the production of chlamydospores (shown in Table 2 and see Figure 2B).

(A) Symptom of isolate CF/UENF 501 in popcorn leaf; (B) Colony Front; (C) Colony Back; (D) Conidia in PDA medium.
3.2. Pathogenicity of the isolate
All lines showed lesions in different parts of the inoculated leaves, showing high pathogenicity of the isolate used in the experiment, which can be confirmed with the high disease incidence symptoms of plants and the different levels of average severity (see Figure 3).

Environmental gradient of B. maydis symptoms in the experimental units. (A) Incidence of B. maydis; (B) Average severity B. maydis.
The severity assessment over time allowed to follow the disease progress. Summarizing the results through the regression lines made it possible to study its coefficients for each genotype. In this way, the Linear Coefficient (LC) on the axis (y) is considered the starting point of the severity of leaf symptoms. The Angular Coefficient (AC) represented the slope of the estimated straight line for each genotype, as the rate of addition of symptoms that multiplies time on the (x) axis.
The variance analysis made it possible to detect significant differences in LC and AC variables between the lines (shown in Table 3). It indicates that the genotypes react differently to the B. maydis isolate initially and throughout the SCLB epidemic in popcorn plants due to their genetic resistance.
Summary of the variance analysis for the linear coefficient (LC) and angular coefficient (AC) variables obtained from the linear regression model of B. maydis severity over time.
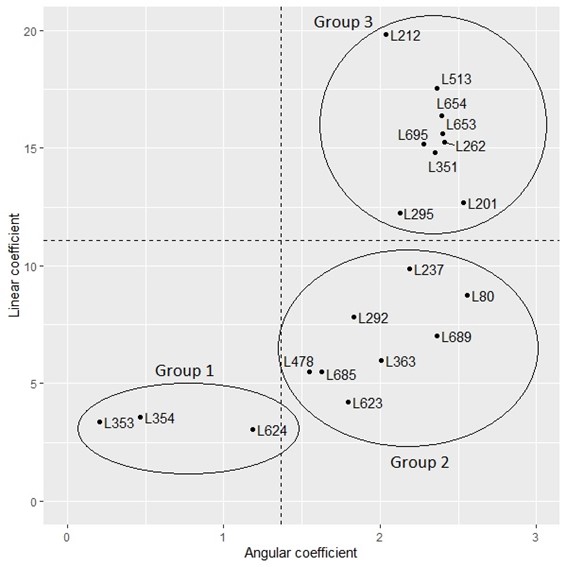
The cluster analysis of the genotypes based on the estimated coefficients for the regressions of the progress of disease symptoms allowed us to observe the separation of two similar groups for LC and AC, respectively. 55% of the lines were grouped with the lowest LC, and 15% of the total lines were gathered with the lowest AC value (see Figure 4).

Dispersion of popcorn lines in terms of linear coefficient (LC) and angular coefficient (AC) obtained from the linear regression model of B. maydis severity over time. Horizontal line delimits the limit of similar groups for LC, and vertical line indicates the limit for AC formed by the Scott-Knott test at 5%.
When comparing the grouping results for the two variables, it was observed that the lines separated with the formation of three groups. Group 1, with 15% of the lines, corresponds to the lowest LC and AC values. Group 2, with 40% of the lines, corresponds to the lowest LC and the highest AC values simultaneously, and group 3, with 45% of the lines, corresponds to the highest values for both coefficients.
As for the groups formed, there is one with the highest resistance level, comprising L353, L354, and L624. For these lines, the regression line estimates showed low values for the coefficients. The severity assessed in these treatments was initially low, and there was no significant severity increase over time, evidenced by the low values found for the addition rate.
Lines L237, L292, L363, L478, L623, L624, L685, L689, and L80, considered intermediate, showed few symptoms at the beginning of the severity assessment. However, there was an expressive increase in the severity of B. maydis over time, indicating a partial susceptibility of these genotypes. Genotypes L201, L212, L513, L654, L653, L262, L351, L295, and L695 formed a group considered susceptible for presenting similar responses, at the beginning and throughout the evaluations, with a high severity level.
3.3. Characterization of symptoms
The inoculation of the CF/UENF 501 isolate of B. maydis conferred three distinct patterns of symptoms among the evaluated genotypes (see Figure 5). The symptoms found in popcorn leaves were A) fusiform or elliptical lesions with a dark brown border; B) elongated brown lesions; C) orange punctate lesions. The appearance of type A) lesions were the most prevalent among the genotypes evaluated.

Different symptoms found in popcorn genotypes inoculated with B. maydis CF/UENF 501 isolate.
4. Discussion
4.1. Isolate characterization
The characterization of the isolate from the morphological evaluation allowed the pathogen identification. Pal et al. (2015)PAL, I., SINGH, V., GOGOI, R., HOODA, K.S. and BEDI, N., 2015. Characterization of Bipolaris maydis isolates of different maize cropping zones of India. Indian Phytopathology, vol. 68, no. 1, pp. 63-66. and Santos et al. (2016)SANTOS, J.S., AMARAL JÚNIOR, A.T., VIVAS, M., VIVAS, J.M.S., KUROSAWA, R.N.F., SILVEIRA, S.F., 2016. Características culturais e patológicas de Bipolaris maydis em diferentes meios de cultura. Revista Brasileira de Milho e Sorgo, vol. 15, no. 3, pp. 461-469. http://dx.doi.org/10.18512/1980-6477/rbms.v15n3p461-469.
http://dx.doi.org/10.18512/1980-6477/rbm...
found a similar coloration to that found in this work, based on the morphological characterization of the colonies.
Manamgoda et al. (2014)MANAMGODA, D.S., ROSSMAN, A.Y., CASTLEBURY, L.A., CROUS, P.W., MADRID, H., CHUKEATIROTE, E. and HYDE, K.D., 2014. The genus Bipolaris. Studies in Mycology, vol. 79, pp. 221-288. http://dx.doi.org/10.1016/j.simyco.2014.10.002. PMid:25492990.
http://dx.doi.org/10.1016/j.simyco.2014....
described the species B. maydis regarding colony morphology, conidia, and symptoms found in Zea mays leaves. These results are similar to those found in this work for macromorphological and micromorphological characterization, indicating that the isolate “CF/UENF” 501 belongs to the B. maydis species. In addition to agreeing with the results found by Marin-Felix et al. (2017)MARIN-FELIX, Y., SENWANNA, C., CHEEWANGKOON, R. and CROUS, P.W., 2017. New species and records of Bipolaris and Curvularia from Thailand. Mycosphere, vol. 8, no. 9, pp. 1556-1574. http://dx.doi.org/10.5943/mycosphere/8/9/11.
http://dx.doi.org/10.5943/mycosphere/8/9...
for species identification based on molecular technique. Sun et al. (2020)SUN, X., QI, X., WANG, W., LIU, X., ZHAO, H., WU, C., CHANG, X., ZHANG, M., CHEN, H. and GONG, G., 2020. Etiology and symptoms of maize leaf spot caused by Bipolaris spp. in Sichuan, China. Pathogens, vol. 9, no. 3, pp. 229. http://dx.doi.org/10.3390/pathogens9030229. PMid:32244886.
http://dx.doi.org/10.3390/pathogens90302...
summarized crop morphological and microscopy traits and the symptom characterization in corn. They were able to identify distinct species of Bipolaris by combining data of morphological, molecular, and symptomatology.
4.2. Pathogenicity of the isolate
The presence of disease symptoms in all plants of different lines inoculated with the isolate “CF/UENF 501” demonstrated the high pathogenicity of isolates in different genetic samples of the host. As for disease severity, there was a difference between the inoculated lines. This result reflects the genetic resistance of lines since genotypes are obtained from different populations with dissimilar genetic characteristics.
The resistance reaction of genotypes to a pathogen can be due to limiting the multiplication of pathogen (Clarke, 1986CLARKE, D.D., 1986. Tolerance of parasites and disease in plants and its significance in host-parasite interactions. New York: Academic Press, pp. 161-197. Advance in Plant Pathology, no. 5.) or to the delay in its development (Clarke, 1986CLARKE, D.D., 1986. Tolerance of parasites and disease in plants and its significance in host-parasite interactions. New York: Academic Press, pp. 161-197. Advance in Plant Pathology, no. 5.; Little et al., 2010LITTLE, T.J., SHUKER, D.M., COLEGRAVE, N., DAY, T. and GRAHAM, A.L., 2010. The coevolution of virulence: tolerance in perspective. PLoS Pathogens, vol. 6, no. 9, e1001006. http://dx.doi.org/10.1371/journal.ppat.1001006. PMid:20838464.
http://dx.doi.org/10.1371/journal.ppat.1...
; Raberg, 2014RÅBERG, L., 2014. How to live with the enemy: understanding tolerance to parasites. PLoS Biology, vol. 12, no. 11, pp. e1001989. http://dx.doi.org/10.1371/journal.pbio.1001989. PMid:25369060.
http://dx.doi.org/10.1371/journal.pbio.1...
). For SCLB, there is a large variability of popcorn genotypes with different disease resistance degrees (Mubeen et al., 2017MUBEEN, S., RAFIQUE, M., MUNIS, M.F.H. and CHAUDHARY, H.J., 2017. Study of southern corn leaf blight (SCLB) on maize genotypes and its effect on yield. Journal of the Saudi Society of Agricultural Sciences, vol. 16, no. 3, pp. 210-217. http://dx.doi.org/10.1016/j.jssas.2015.06.006.
http://dx.doi.org/10.1016/j.jssas.2015.0...
; Kurosawa et al., 2018KUROSAWA, R.D.N.F., VIVAS, M., AMARAL, A.T.D., RIBEIRO, R.M., MIRANDA, S.B., PENA, G.F., LEITE, J.T. and MORA, F., 2018. Popcorn germplasm resistance to fungal diseases caused by Exserohilum turcicum and Bipolaris maydis. Bragantia, vol. 77, no. 1, pp. 36-47. http://dx.doi.org/10.1590/1678-4499.2017035.
http://dx.doi.org/10.1590/1678-4499.2017...
; Santos et al., 2020SANTOS, J.S., SOUZA, Y.P., VIVAS, M., AMARAL JUNIOR, A.T., ALMEIDA FILHO, J.E., MAFRA, G.S. and FERREIRA, F.R.A., 2020. Genetic merit of popcorn lines and hybrids for multiple foliar diseases and agronomic properties. Functional Plant Breeding Journal, vol. 2, no. 2, pp. 33-47. http://dx.doi.org/10.35418/2526-4117/v2n2a2.
http://dx.doi.org/10.35418/2526-4117/v2n...
; Saluci et al., 2020SALUCI, J.C.G., VIVAS, M., DE ALMEIDA, R.N., DUTRA, I.P., CARLOS, M.C., AMARAL JÚNIOR, A.T. and SCAPIM, C.A., 2020. Potential of Popcorn S4 lines for resistance to southern corn leaf blight. Functional Plant Breeding Journal, vol. 2, no. 2, pp. 79-87. http://dx.doi.org/10.35418/2526-4117/v2n2a6.
http://dx.doi.org/10.35418/2526-4117/v2n...
). However, immune genotypes for the disease in Zea mays have not been described (Bhandari et al., 2017BHANDARI, R.R., ARYAL, L., SHARMA, S., ACHARYA, M., POKHREL, A., APAR, G.C., SALINA, K., SAHADEV, K.C., BHAGARATHI, S., KAMAL, B., ARJUN, C. and PANTHI, S., 2017. Screening of maize genotypes against southern leaf blight (Bipolaris maydis) during summer season in Nepal. World Journal of Agricultural Research, vol. 5, no. 1, pp. 31-41.; Mubeen et al., 2017MUBEEN, S., RAFIQUE, M., MUNIS, M.F.H. and CHAUDHARY, H.J., 2017. Study of southern corn leaf blight (SCLB) on maize genotypes and its effect on yield. Journal of the Saudi Society of Agricultural Sciences, vol. 16, no. 3, pp. 210-217. http://dx.doi.org/10.1016/j.jssas.2015.06.006.
http://dx.doi.org/10.1016/j.jssas.2015.0...
). Thus, when evaluating the response of popcorn lines tested in contact with the pathogen, it is expected that the genotypes with higher resistance levels have low initial severity and low rate of symptom progression due to the action of resistance genes to delay disease development. The inheritance of resistance to SCLB, naturally polygenic, can explain the different levels found for severity (Poland et al., 2009POLAND, J.A., BALINT-KURTI, P.J., WISSER, R.J., PRATT, R.C. and NELSON, R.J., 2009. Shades of gray: the world of quantitative disease resistance. Trends in Plant Science, vol. 14, no. 1, pp. 21-29. http://dx.doi.org/10.1016/j.tplants.2008.10.006. PMid:19062327.
http://dx.doi.org/10.1016/j.tplants.2008...
; Kump et al., 2011KUMP, K.L., BRADBURY, P.J., WISSER, R.J., BUCKLER, E.S., BELCHER, A.R., OROPEZA-ROSAS, M.A., ZWONITZER, J.C., KRESOVICH, S., MCMULLEN, M.D., WARE, D., BALINT-KURTI, P.J. and HOLLAND, J.B., 2011. Genome-wide association study of quantitative resistance to southern leaf blight in the maize nested association mapping population. Nature Genetics, vol. 43, no. 2, pp. 163-168. http://dx.doi.org/10.1038/ng.747. PMid:21217757.
http://dx.doi.org/10.1038/ng.747...
), conferring a quantitative effect on the disease. Thus, the reaction of genotypes occurs with different resistance levels, assessed by the severity symptoms of leaves.
According to Kraja et al. (2000)KRAJA, A., DUDLEY, J.W. and WHITE, D.G., 2000. Identification of tropical and temperate maize populations having favorable alleles for disease resistance. Crop Science, vol. 40, no. 4, pp. 948-954. http://dx.doi.org/10.2135/cropsci2000.404948x.
http://dx.doi.org/10.2135/cropsci2000.40...
, differences in genotype responses regarding disease severity occur due to genetic composition variability. The use of genotypes with different genealogies allowed the identification of differences in SCLB symptoms among the evaluated lines, as it allowed to observe a wide variation in disease severity and the symptoms presented. It is noteworthy that Saluci et al. (2020)SALUCI, J.C.G., VIVAS, M., DE ALMEIDA, R.N., DUTRA, I.P., CARLOS, M.C., AMARAL JÚNIOR, A.T. and SCAPIM, C.A., 2020. Potential of Popcorn S4 lines for resistance to southern corn leaf blight. Functional Plant Breeding Journal, vol. 2, no. 2, pp. 79-87. http://dx.doi.org/10.35418/2526-4117/v2n2a6.
http://dx.doi.org/10.35418/2526-4117/v2n...
point out the PR023 and PA091 populations, which gave rise to the lines mentioned earlier, as a potential source of resistance to B. maydis. Thus, it can be confirmed that the potential of parents for resistance to SCLB is reflected in the high resistance of the originated lines, resulting from the transfer of resistance alleles between the involved genotypes, conferring a low disease severity in those lines.
The tested lines were divided into three groups according to the pathogenicity response of isolates. Group 1 stood out with the lines showing reduced severity at the beginning of evaluations and end. Group 2 contains the lines with low severity only at the beginning of the evaluations, and group 3, consists of lines that showed high severity at the beginning of the evaluations. The SCLB severity quantification over time and the separation of the three groups based on the resistance response indicated that the SCLB resistance expression observed in the lines reflected the high virulence of the isolate. Thus, the pathogen was sufficiently aggressive to the point of provoking the activation of the defense mechanisms of plants, generating different responses in the intensity and format of symptoms expressed in the lines. The use of highly virulent isolates is essential for evaluating and selecting genotypes since low virulent ones do not allow for the generation of different responses and, consequently, group separation regarding the disease levels.
Disease intensity, color, and shape were the aspects that differed in the inoculated lines. Disease intensity is associated with severity, and this, in turn, is strongly influenced by inoculum pressure. Shah et al. (2016)SHAH, W.U., NAEEM, A., ADNAN, M., JUNAID, K., SHAH, S.R.A. and ATTAULLAH, M.I., 2016. Study on the response of different maize cultivars to various inoculum levels of Bipolaris maydis (Y. Nisik & C. Miyake) shoemaker under field conditions. Journal of Entomology and Zoology Studies, vol. 4, no. 2, pp. 533-537. observed that the higher the inoculum pressure, the greater the severity of B. maydis. It is common for the inoculum pressure to influence these aspects strongly in field experiments involving natural inoculation since the inoculum pressure may not be uniformly distributed in the cultivation area. This can result in a great possibility of finding genotypes with different levels of resistance. However, when the evaluation conditions are controlled, the inoculum pressure is homogeneous in all genotypes. Therefore, the variation found in the lines reflects the resistance response of the genotypes to the isolate (Hussain et al., 2016HUSSAIN, H., RAZIQ, F., KHAN, I., SHAH, B., ALTAF, M., ATTAULLAH, U.W., NAEEM, A., ADNAN, M., JUNAID, K., SHAH, S.R.L. and IQBAL, M., 2016. Effect of Bipolaris maydis (Y. Nisik & C. Miyake) shoemaker at various growth stages of different maize cultivars. Journal of Entomology and Zoology Studies, vol. 4, no. 2, pp. 439-444.).
The spot caused by B. maydis on Zea mays may show symptoms with brown coloration, reddish margins, elliptical at first, and elongating to a rectangular shape, which may vary according to the cultivar or according to the race of the pathogen (Sun et al., 2020SUN, X., QI, X., WANG, W., LIU, X., ZHAO, H., WU, C., CHANG, X., ZHANG, M., CHEN, H. and GONG, G., 2020. Etiology and symptoms of maize leaf spot caused by Bipolaris spp. in Sichuan, China. Pathogens, vol. 9, no. 3, pp. 229. http://dx.doi.org/10.3390/pathogens9030229. PMid:32244886.
http://dx.doi.org/10.3390/pathogens90302...
). The species B. maydis can elicit different virulence degrees based on the size and type of lesion it caused in the host (Sun et al., 2020SUN, X., QI, X., WANG, W., LIU, X., ZHAO, H., WU, C., CHANG, X., ZHANG, M., CHEN, H. and GONG, G., 2020. Etiology and symptoms of maize leaf spot caused by Bipolaris spp. in Sichuan, China. Pathogens, vol. 9, no. 3, pp. 229. http://dx.doi.org/10.3390/pathogens9030229. PMid:32244886.
http://dx.doi.org/10.3390/pathogens90302...
). Considering that the symptoms observed resulted from a single isolate, the symptom variations found in this work are exclusively the result of the lines' defense mechanisms in contact with the virulent isolate.
5. Conclusions
The morphological characterization of the CF/UENF 501 isolate and the pathogenicity test are strong indications that the fungus is truly the species B. maydis. However, further studies with molecular markers should be used for better confirmation.
The analysis of the estimated regression coefficients allowed inferences about the disease progress. It evidenced the ability of the pathogen to discriminate the lines in terms of resistance in at least three resistance classes.
Lines L353, L354, and L624 were identified as potential sources of resistance to Southern Corn Leaf Blight.
The different levels of severity and symptoms found among the lines may indicate that the high virulence of the pathogen was essential to generate different responses on resistance to B. maydis in popcorn lines.
Acknowledgements
The authors are grateful for funding from Fundação Carlos Chagas Filho de Amparo à Pesquisa do Estado do Rio de Janeiro and Conselho Nacional de Desenvolvimento Científico e Tecnológico for financial support. This study was financed in part by the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior – Brasil (CAPES) – Finance Code 001.
References
- AGRIOS, G.N., 2005. Foliar diseases caused by Ascomycetes and Deuteromycetes (Mitosporic Fungi). In: G. AGRIOS, ed. Plant pathology 5th ed. Burlington: Elsevier Academic Press, pp. 466-468.
- ALI, F., RAHMAN, H., DURRISHAHWAR, N.F., MUNIR, M. and ULLAH, H., 2011. Genetics analysis and maturity and morphological traits under maydis leaf blight epiphytotic in maize. Journal of Agricultural and Biological Science, vol. 6, no. 8, pp. 13-19.
- BHANDARI, R.R., ARYAL, L., SHARMA, S., ACHARYA, M., POKHREL, A., APAR, G.C., SALINA, K., SAHADEV, K.C., BHAGARATHI, S., KAMAL, B., ARJUN, C. and PANTHI, S., 2017. Screening of maize genotypes against southern leaf blight (Bipolaris maydis) during summer season in Nepal. World Journal of Agricultural Research, vol. 5, no. 1, pp. 31-41.
- BURNETTE, D.C. and WHITE, D.G., 1985. Inheritance of resistance to Bipolaris maydis race O in crosses derived from nine resistance inbred lines of maize. Phytopathology, vol. 75, no. 11, pp. 1195-1200. http://dx.doi.org/10.1094/Phyto-75-1195
» http://dx.doi.org/10.1094/Phyto-75-1195 - CLARKE, D.D., 1986. Tolerance of parasites and disease in plants and its significance in host-parasite interactions New York: Academic Press, pp. 161-197. Advance in Plant Pathology, no. 5.
- HOLLEY, R.N. and GOODMAN, M.M., 1989. New sources of resistance to Southern corn leaf blight from tropical hybrid maize derivatives. Plant Disease, vol. 73, no. 7, pp. 562-564. http://dx.doi.org/10.1094/PD-73-0562
» http://dx.doi.org/10.1094/PD-73-0562 - HUSSAIN, H., RAZIQ, F., KHAN, I., SHAH, B., ALTAF, M., ATTAULLAH, U.W., NAEEM, A., ADNAN, M., JUNAID, K., SHAH, S.R.L. and IQBAL, M., 2016. Effect of Bipolaris maydis (Y. Nisik & C. Miyake) shoemaker at various growth stages of different maize cultivars. Journal of Entomology and Zoology Studies, vol. 4, no. 2, pp. 439-444.
- JAMES, W.C., 1971. A manual of assessment keys of plant diseases St. Paul: American Phytopathological Society Press. Publ., no. 1458.
- KRAJA, A., DUDLEY, J.W. and WHITE, D.G., 2000. Identification of tropical and temperate maize populations having favorable alleles for disease resistance. Crop Science, vol. 40, no. 4, pp. 948-954. http://dx.doi.org/10.2135/cropsci2000.404948x
» http://dx.doi.org/10.2135/cropsci2000.404948x - KUMP, K.L., BRADBURY, P.J., WISSER, R.J., BUCKLER, E.S., BELCHER, A.R., OROPEZA-ROSAS, M.A., ZWONITZER, J.C., KRESOVICH, S., MCMULLEN, M.D., WARE, D., BALINT-KURTI, P.J. and HOLLAND, J.B., 2011. Genome-wide association study of quantitative resistance to southern leaf blight in the maize nested association mapping population. Nature Genetics, vol. 43, no. 2, pp. 163-168. http://dx.doi.org/10.1038/ng.747 PMid:21217757.
» http://dx.doi.org/10.1038/ng.747 - KUROSAWA, R.D.N.F., VIVAS, M., AMARAL, A.T.D., RIBEIRO, R.M., MIRANDA, S.B., PENA, G.F., LEITE, J.T. and MORA, F., 2018. Popcorn germplasm resistance to fungal diseases caused by Exserohilum turcicum and Bipolaris maydis. Bragantia, vol. 77, no. 1, pp. 36-47. http://dx.doi.org/10.1590/1678-4499.2017035
» http://dx.doi.org/10.1590/1678-4499.2017035 - LITTLE, T.J., SHUKER, D.M., COLEGRAVE, N., DAY, T. and GRAHAM, A.L., 2010. The coevolution of virulence: tolerance in perspective. PLoS Pathogens, vol. 6, no. 9, e1001006. http://dx.doi.org/10.1371/journal.ppat.1001006 PMid:20838464.
» http://dx.doi.org/10.1371/journal.ppat.1001006 - MANAMGODA, D.S., ROSSMAN, A.Y., CASTLEBURY, L.A., CROUS, P.W., MADRID, H., CHUKEATIROTE, E. and HYDE, K.D., 2014. The genus Bipolaris. Studies in Mycology, vol. 79, pp. 221-288. http://dx.doi.org/10.1016/j.simyco.2014.10.002 PMid:25492990.
» http://dx.doi.org/10.1016/j.simyco.2014.10.002 - MARIN-FELIX, Y., SENWANNA, C., CHEEWANGKOON, R. and CROUS, P.W., 2017. New species and records of Bipolaris and Curvularia from Thailand. Mycosphere, vol. 8, no. 9, pp. 1556-1574. http://dx.doi.org/10.5943/mycosphere/8/9/11
» http://dx.doi.org/10.5943/mycosphere/8/9/11 - MUBEEN, S., RAFIQUE, M., MUNIS, M.F.H. and CHAUDHARY, H.J., 2017. Study of southern corn leaf blight (SCLB) on maize genotypes and its effect on yield. Journal of the Saudi Society of Agricultural Sciences, vol. 16, no. 3, pp. 210-217. http://dx.doi.org/10.1016/j.jssas.2015.06.006
» http://dx.doi.org/10.1016/j.jssas.2015.06.006 - O’NEILL, M.E. and MATHEWS, K.L., 2002. Levene tests of homogeneity of variance for general block and treatment designs. Biometrics, vol. 58, no. 1, pp. 216-224. http://dx.doi.org/10.1111/j.0006-341X.2002.00216.x PMid:11890318.
» http://dx.doi.org/10.1111/j.0006-341X.2002.00216.x - PAL, I., SINGH, V., GOGOI, R., HOODA, K.S. and BEDI, N., 2015. Characterization of Bipolaris maydis isolates of different maize cropping zones of India. Indian Phytopathology, vol. 68, no. 1, pp. 63-66.
- PATE, R.T., WEATHERLY, M., HEDGES, L.B., MIDEROS, S., FELLOWS, G.M. and CARDOSO, F., 2017. 363 effects of foliar fungicide on whole plant brown midrib and floury corn varieties. Journal of Animal Science, vol. 95, no. 2, suppl. 2, pp. 176-177. http://dx.doi.org/10.2527/asasmw.2017.363
» http://dx.doi.org/10.2527/asasmw.2017.363 - POLAND, J.A., BALINT-KURTI, P.J., WISSER, R.J., PRATT, R.C. and NELSON, R.J., 2009. Shades of gray: the world of quantitative disease resistance. Trends in Plant Science, vol. 14, no. 1, pp. 21-29. http://dx.doi.org/10.1016/j.tplants.2008.10.006 PMid:19062327.
» http://dx.doi.org/10.1016/j.tplants.2008.10.006 - RÅBERG, L., 2014. How to live with the enemy: understanding tolerance to parasites. PLoS Biology, vol. 12, no. 11, pp. e1001989. http://dx.doi.org/10.1371/journal.pbio.1001989 PMid:25369060.
» http://dx.doi.org/10.1371/journal.pbio.1001989 - SALUCI, J.C.G., VIVAS, M., DE ALMEIDA, R.N., DUTRA, I.P., CARLOS, M.C., AMARAL JÚNIOR, A.T. and SCAPIM, C.A., 2020. Potential of Popcorn S4 lines for resistance to southern corn leaf blight. Functional Plant Breeding Journal, vol. 2, no. 2, pp. 79-87. http://dx.doi.org/10.35418/2526-4117/v2n2a6
» http://dx.doi.org/10.35418/2526-4117/v2n2a6 - SANTOS, J.S., AMARAL JÚNIOR, A.T., VIVAS, M., VIVAS, J.M.S., KUROSAWA, R.N.F., SILVEIRA, S.F., 2016. Características culturais e patológicas de Bipolaris maydis em diferentes meios de cultura. Revista Brasileira de Milho e Sorgo, vol. 15, no. 3, pp. 461-469. http://dx.doi.org/10.18512/1980-6477/rbms.v15n3p461-469
» http://dx.doi.org/10.18512/1980-6477/rbms.v15n3p461-469 - SANTOS, J.S., SOUZA, Y.P., VIVAS, M., AMARAL JUNIOR, A.T., ALMEIDA FILHO, J.E., MAFRA, G.S. and FERREIRA, F.R.A., 2020. Genetic merit of popcorn lines and hybrids for multiple foliar diseases and agronomic properties. Functional Plant Breeding Journal, vol. 2, no. 2, pp. 33-47. http://dx.doi.org/10.35418/2526-4117/v2n2a2
» http://dx.doi.org/10.35418/2526-4117/v2n2a2 - SHAH, W.U., NAEEM, A., ADNAN, M., JUNAID, K., SHAH, S.R.A. and ATTAULLAH, M.I., 2016. Study on the response of different maize cultivars to various inoculum levels of Bipolaris maydis (Y. Nisik & C. Miyake) shoemaker under field conditions. Journal of Entomology and Zoology Studies, vol. 4, no. 2, pp. 533-537.
- SHAPIRO, S.S. and WILK, M.B., 1965. An analysis of variance test for normality (complete sample). Biometrika, vol. 52, no. 3, pp. 591-611. http://dx.doi.org/10.1093/biomet/52.3-4.591
» http://dx.doi.org/10.1093/biomet/52.3-4.591 - SUN, X., QI, X., WANG, W., LIU, X., ZHAO, H., WU, C., CHANG, X., ZHANG, M., CHEN, H. and GONG, G., 2020. Etiology and symptoms of maize leaf spot caused by Bipolaris spp. in Sichuan, China. Pathogens, vol. 9, no. 3, pp. 229. http://dx.doi.org/10.3390/pathogens9030229 PMid:32244886.
» http://dx.doi.org/10.3390/pathogens9030229 - WHITE, D.G., 1999. Compendium of corn diseases 3rd ed. St. Paul: American Phytopathological Society, 128 p.
Publication Dates
-
Publication in this collection
02 Feb 2022 -
Date of issue
2024
History
-
Received
28 Sept 2021 -
Accepted
16 Dec 2021