SUMMARY
The objective of this study was to evaluate the macronutrient accumulations in leaf, stem, and shoot of elephant grass cv. Roxo. The treatments consisted of seven growth ages (9, 18, 27, 36, 45, 54, and 63 days) and three growing seasons (rainy, transition, and dry). A completely randomized design and a split-plot time arrangement were used, where the ages were the plots and the seasons the subplots, with three replications. Interaction between the factors age and season of cultivation for the contents and the daily accumulation rates of N, P, K, Ca, Mg, and S in leaf, stem, and shoot fractions were calculated. The margins of daily accumulation of nutrients N, P, K, Mg, and S presented positive linear behavior in all seasons studied; however, with variation in the magnitude of the response between them. The daily accumulation rate of Ca in the canopy was adjusted to a linear model for the rainy and dry seasons and to a quadratic model for the transition season, reaching a maximum of 4.60 kg ha-1 at 59.76 days of growth. The accumulation of nutrients in elephant grass cv. Roxo showed the following decreasing order: K > N > P > Mg > Ca > S in the rainy and transition seasons, while in the dry season the following decreasing order was observed: K > P > N > Ca > Mg > S.
Keywords:
absorption run; elephant grass; mineral nutrition
RESUMO
Objetivou-se avaliar a marcha de acúmulo de macronutrientes na folha, colmo e na parte aérea do capim-elefante cv. Roxo. Os tratamentos consistiram em sete idades de crescimento (9; 18; 27; 36; 45; 54 e 63 dias) em três períodos de cultivo (chuvoso, de transição e seco). Adotou-se um delineamento inteiramente casualizado e um arranjo de parcelas subdivididas no tempo, onde as idades foram alocadas nas parcelas e os períodos de cultivo nas subparcelas, com três repetições. Constatou-se interação entre os fatores idade e período de cultivo para os teores e para as taxas de acúmulo diário de N, P, K, Ca, Mg e S nas frações folha, colmo e parte aérea. As marchas de acúmulo diário dos nutrientes N, P, K, Mg e S apresentaram comportamento linear positivo para todos os períodos estudados, todavia com variação na magnitude da resposta entre estes. A taxa de acúmulo diário de Ca na parte aérea do dossel ajustou-se ao modelo linear para os períodos chuvoso e seco e ao modelo quadrático para o período de transição, atingindo máximo de 4,60 kg ha-1 aos 59,76 dias de crescimento. O acúmulo dos nutrientes no capim-elefante cv. Roxo apresentou a seguinte ordem decrescente: K > N > P > Mg > Ca > S no período chuvoso e de transição enquanto no período seco observou-se a seguinte ordem decrescente: K > P > N > Ca > Mg> S.
Palavras-chave:
marcha de absorção; capimelefante; nutrição mineral
INTRODUCTION
An efficient management of fertilization requires knowledge of nutrient accumulation (ECHER et al., 2009ECHER, F.R.; DOMINATO, J.C.; CRESTE, J.E. Absorção de nutrientes e distribuição da massa fresca e seca entre órgãos de batata-doce. Horticultura Brasileira, v.27, n.2, p.176-182, 2009.). The knowledge of the nutritional requirements in each phase of the growth cycle would make it possible to improve the synchrony between availability and nutrient requirements of the plant, making it possible to increase the agronomic and economic efficiency of the system. The accumulation and exportation of nutrients by the plant, on the other hand, varies according to the age of the plant, plant's organ, species, cultivar, type of nutrient, management, and edaphoclimatic conditions in which the canopy develops (COSTA et al., 2010COSTA, K.A.P.; OLIVEIRA, I.P.; SEVERIANO, E.C.; SAMPAIO, F.M.T.; CARRIJO, M.S.; RODRIGUES, C.R. Extração de nutrientes pela fitomassa de cultivares de brachiaria brizantha sob doses de nitrogênio. Ciência Animal Brasileira, v.11, n.2, p.307-314, 2010.; SILVA et al., 2015SILVA, M.R.; PELISSARI, A.; MORAES, A.; SANDINI, I.E.; CASSOL, L.C.; ASSMANN, T.S.; OLIVEIRA, E.B. Acumulação de nutrientes e produção forrageira de aveia e azevém em função da aplicação de calcário e gesso em superfície. Revista de Ciências Agrárias, v.38, n.3, p.346-356, 2015.).
Diffused in tropical and subtropical regions, elephant grass (Pennisetum purpureum) has been distinguished by relevant and appreciable agronomic and bromatological characteristics for the production of ruminants. It is quite responsive to management; however, very demanding in soil fertility for maintenance of its productive potential, which may vary according to the edaphoclimatic conditions and cultivar adopted.
Most studies on nutrient accumulation are carried out in intensive irrigation systems, making it difficult to extend the results to plants managed on dry lands. It is worth mentioning the scarcity of studies on the partition of nutrients in forage plants. Thus, the demand for research on nutrient accumulation in tropical regions is explicit, considering the variation of rainfall, in particular regarding the requirements of macronutrients by elephant grass cv. Roxo. It is also worth noting that studies of this nature make it possible to generate predictive models of macronutrient accumulation by elephant grass under dry conditions. Thus, the objective was to evaluate the macronutrient accumulations in the leaf, stem, and shoot of elephant grass cv. Roxo managed at different ages and seasons of growth under waterless conditions.
MATERIAL AND METHODS
The experiment was carried out at the Núcleo de Ensino e Estudos em Forragicultura of the Departamento de Zootecnia of the Centro de Ciências Agrárias of the Universidade Federal do Ceará-NEEF/DZ/CCA/UFC, located at latitude 03° 44’ 32’ S' and longitude 38° 34’ 40”W in Fortaleza-CE, with a rainy tropical Aw' climate classification according to Köppen (1936)KÖPPEN, W. Das geographisca System der Klimate. Gebr, Borntraeger, 1936. 1-44p.. The study was carried out from May to November of 2012, in an area of elephant grass (Pennisetum purpureum) cv. Roxo established five years prior to the study. The soil of the area is classified as yellow Argisol, with a sandy texture. Three successive cycles of growth in distinct seasons (rainy, transition, and dry) were evaluated according to rainfall during the experimental timeframe.
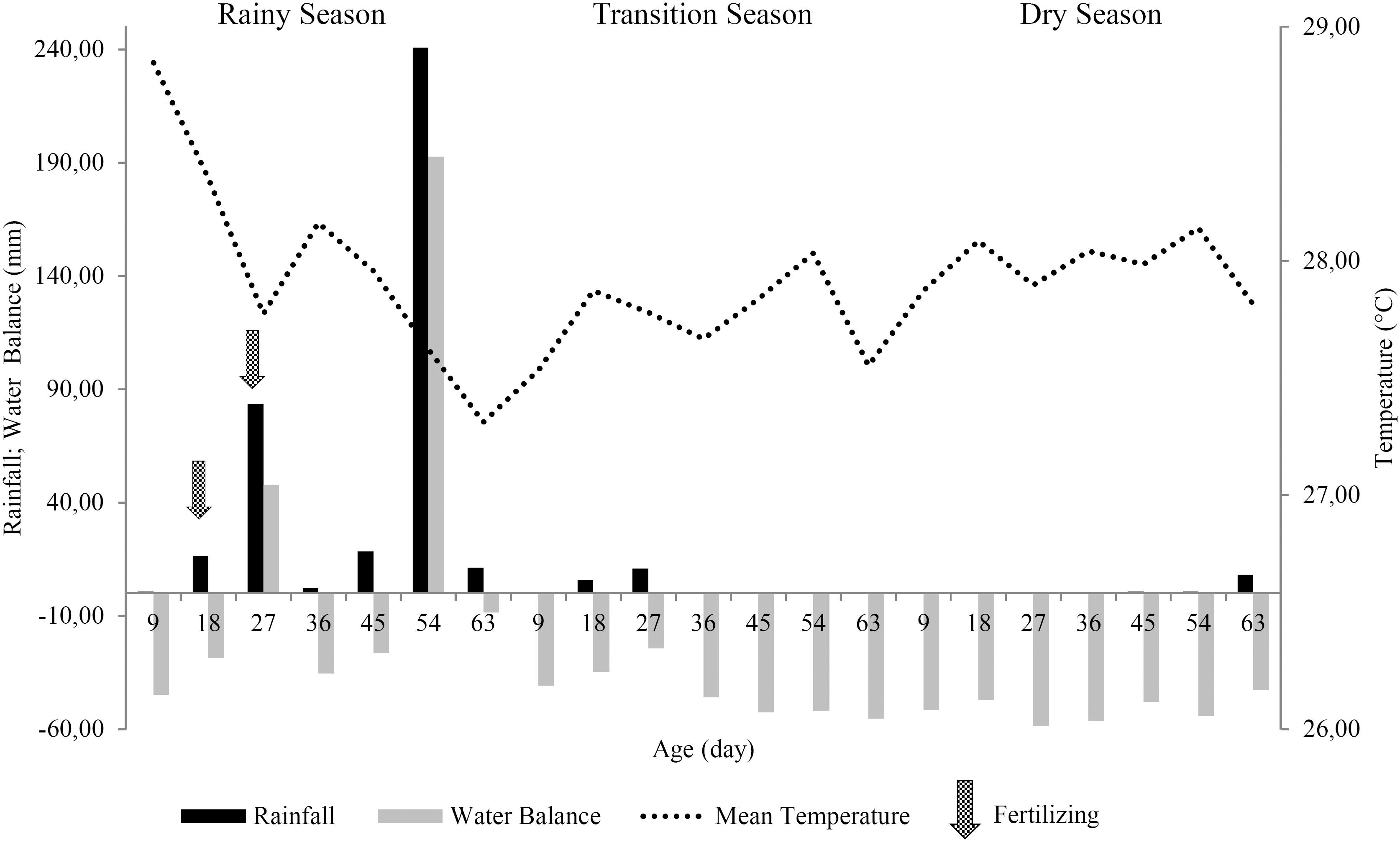
There was accumulated rainfall of 373.30, 17.30, and 9.60 mm in the rainy, transition, and dry seasons, respectively. While the potential evapotranspiration of the crop presented values of 39.43, 46.11, and 52.58 mm day-1 for rainy, transition, and dry season, respectively. The climatological variables were obtained at the Meteorological Station of the Universidade Federal do Ceará, Campus do Pici (Figure 1).
The accumulation of macronutrients was quantified at seven growth ages (9, 18, 27,36, 45, 54, and 63 days after cutting close to the soil) during three successive growing cycles. The design was completely randomized in a split-plot time arrangement, where the ages were the plots and the seasons (rainy, transition, and dry) the subplots, with three replications. The area of each experimental unit was 3.0 × 3.5m.
Prior to the experimental period, two standardization cuts were performed (January and April of 2012) aiming to homogenize the canopy. Due to the standardization cuts, phosphate fertilization was applied in January based on the soil analysis at the time, which justifies the high phosphorous contents at the later soil analysis, making cover phosphate fertilization not necessary.
Before the beginning of the experiment, soil chemical analysis of the experimental area of the layer of 0 to 20 cm was carried out, presenting the following results: pH in water 6.0; 20.89 g kg-1of O.M.; 397 and 3.91 mg dm-3 of P and K, respectively; and 2.8, 1.5, 0.15, and 0.04 cmolc dm-3 of Ca++, Mg++, Al3+, and Na+, respectively.
Maintenance fertilization was carried out with doses equivalent to 200 kg ha-1 of K2O (potassium chloride), 600 kg ha-1 year-1 of nitrogen (urea), and 50 kg ha-1 of FTE BR-12. The recommendations of potassium and FTE BR-12 were based on the Fertility Commission of the State of Minas Gerais (CFSEMG, 1999COMISSÃO DE FERTILIDADE DO SOLO DO ESTADO DE MINAS GERAIS - CFSEMG. Recomendações para uso de corretivos e fertilizantes em Minas Gerais – 5a Aproximação. 5.ed. Viçosa, MG: Universidade Federal de Viçosa, 1999. 359p.). Fertilization was fractionated into two doses, the first one (100% of FTE BR-12, 50% of nitrogen, and 50% of potassium) was administered at day 18 and the second one was administered at day 27. It should be noted that the fertilization was carried out exclusively during the rainy season, due to the high temporal variation of rainfall characteristic of the region, so that it was decided to administer the nutrients at the time of highest rainfall probability. At the end of each growing season, a soil chemical analysis (layer 0-20 cm) was performed to characterize soil fertility (Table 1).
At each growing age in the three cultivation periods, a sample of total forage biomass (TFB) was collected, using a frame of known area (1.0 × 1.0 m). TFB was fractionated into green leaf blade biomass, green stem biomass, and dead forage biomass (data not shown). All the fractions of the plant were predried in a forced ventilation oven at 65 °C until reaching constant weight. The predried biomass of the plant fractions were summed and used to estimate the TFB productivity of elephant grass cv. Roxo managed under cutting at ground level.
The pre-dried green leaf blade and stem biomasses were ground and submitted to nitroperchloric digestion to determine the nitrogen (N), phosphorus (P), potassium (K), calcium (Ca), magnesium (Mg), and sulfur (S) contents according the methodology described by Silva (2009)SILVA, F.C. Manual de análises químicas de solos, plantas e fertilizantes. 2.ed. Brasília, DF: Embrapa informação tecnológica / Embrapa solos, 2009. 627p.. The accumulated amount of each nutrient in leaves and stems of elephant grass cv. Roxo was determined by multiplication of the estimated biomass of each fraction (kg ha-1) versus the content of the respective nutrients. The accumulation of nutrients in the shoot was obtained by the sum of the accumulated amounts in the leaf and the stem, in each growth age.
Data were submitted to analysis of variance, mean comparison test, and regression analysis. The interaction age x season was deployed when significant (p<0.05) by F test for nutrient accumulation in leaf, stem, and shoot fractions. Only the main effect of the seasons was presented for nutrient content. The seasons (rainy, transition, and dry) were compared by the Scott Knott test (p<0.05). The ages were evaluated by regression analysis (T test, p<0.05). The models were selected from the significance of linear and quadratic coefficients and the coefficient of determination. As a tool for statistical analysis, the computational software System for Statistical and Genetic Analysis (SAEG) version 9.1 (UFV, 2007UNIVERSIDADE FEDERAL DE VIÇOSA - UFV. SAEG - sistema para análises estatísticas. Versão 9.1. Viçosa, MG: Fundação Arthur Bernardes, 2007.) was used.
RESULTS AND DISCUSSION
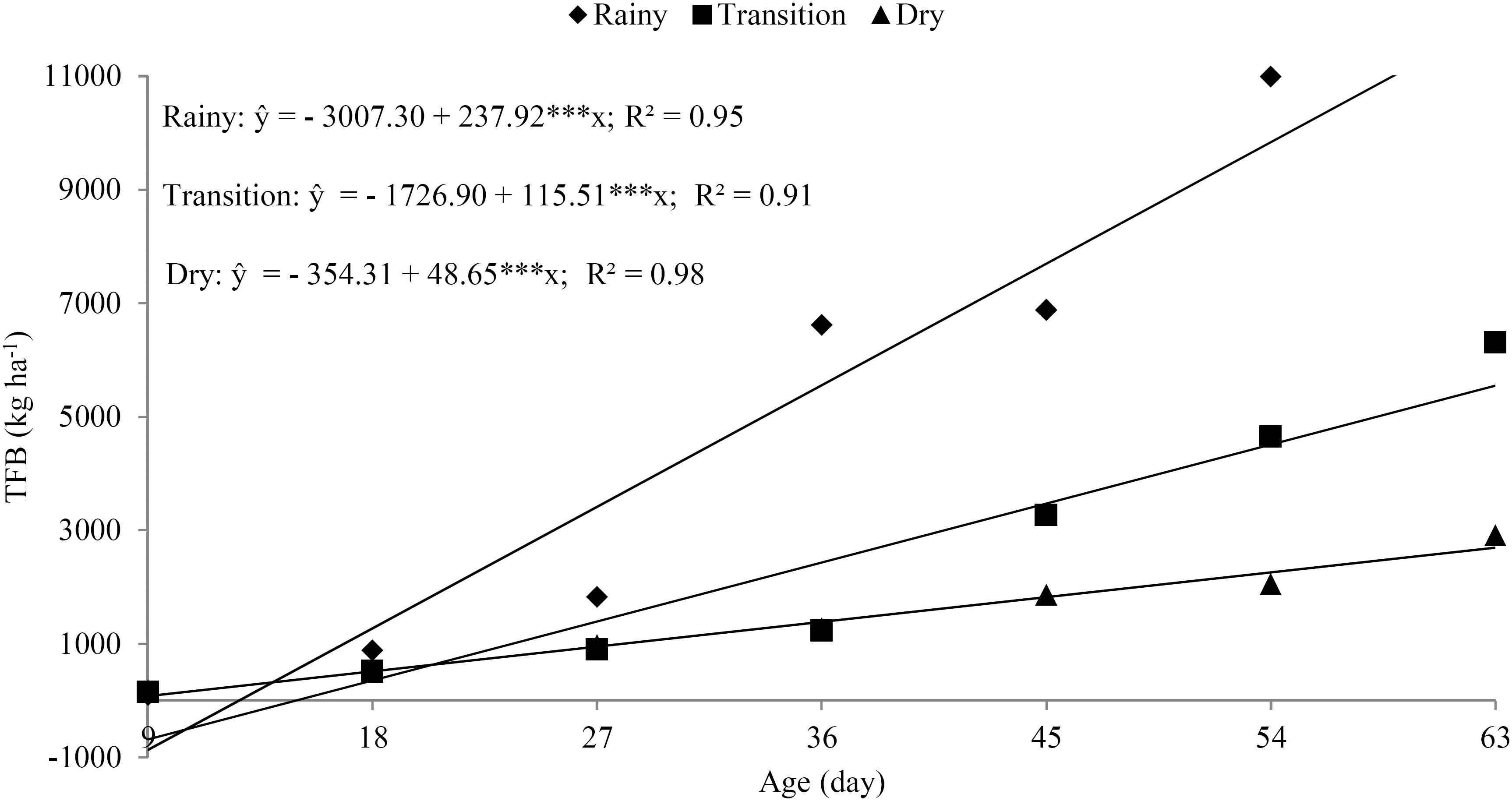
From the proposed equations, yields of 11,869.26, 5,550.23, and 2,692.68 kg ha-1 were estimated at age 63 days after cutting (DAC) in the rainy, transition, and dry seasons, respectively. The positive linear increment for total forage biomass of the elephant grass as a function of the growth ages (Figure 2) helps explain the linear response for the accumulation of most of the studied nutrients. It is worth noting that the forage was managed only up to 63 days of age in order to match the productivity and quality of the biomass produced, since harvest at later ages leads to qualitative reduction of the forage due to a lower leaf/stem ratio, an increase in the rumen undegradable fiber fraction, and a reduction in the crude protein of the forage (BHERING et al., 2008BHERING, M.; CABRAL, L.S.; ABREU, J.G.; SOUZA, A.L.; ZERVOUDAKIS, J.T.; RODRIGUES, R.C.; PEREIRA, G.A.C.; REVERDITO, R.; OLIVEIRA, Í. S. Características agronômicas do capim- elefante roxo em diferentes idades de corte na Depressão Cuiabana. Revista Brasileira de Saúde e Produção Animal [online], v.9, n.3, p.384-396, 2008.; LOPES et al., 2013aLOPES, M.N.; CÂNDIDO, M.J.D.; POMPEU, R.C.F.F.; SILVA, R.G.; LOPES, J.W.B.; FERNANDES, F.R.B.; LACERDA, C.L.; BEZERRA, F.M.L. Fluxo de biomassa em capim-massai durante o estabelecimento e rebrotação com e sem adubação nitrogenada. Revista Ceres, v.60, n.3, p.363-371, 2013a.).
Total forage biomass (BFT) of the shoot; coefficient of determination (R2), significant at the level of 0.1% (***)
The studied factors, growth age and growing season, presented an interaction (p <0.001) for nitrogen (N), phosphorus (P), potassium (K), calcium (Ca), magnesium (Mg), and sulfur (S) contents in leaf and stem fractions, except for the P and K content in the stem fraction (Tables 2 and 3). It should be noted that the macronutrient contents were not submitted to regression analysis, only to mean comparison test, comparing the contents between the growing seasons at each growth age, which were used as indicators of the nutritional status of elephant grass cv. Roxo. An interaction (age x season) was also observed for nutrient accumulation in the leaf, stem, and shoot of elephant grass cv. Roxo.
Nitrogen, phosphorus, and potassium contents in the leaf and stem fractions of elephant grass cv. Roxo at different growth ages in three growing seasons
Calcium, magnesium, and sulfur contents in elephant grass cv. Roxo at different growth ages in three growing seasons
According to Werner (1997)WERNER, J.C.; PAULINO, V.T.; CANTARELLA, H.; ANDRADE, N.O.; QUAGGIO, J.A. Forrageiras. In: RAIJ, B.; CANTARELLA, H.; QUAGGIO, J.A.; FURLANI, A.M.C. (Ed.). Recomendações de adubação e calagem para o Estado de São Paulo. 2.ed. Campinas: Instituto Agronômico, 1997. p.263-273 (Boletim Técnico, 100)., the appropriate leaf nitrogen content for elephant grass should be between 15 and 25 g kg-1. In the present study, the leaf nitrogen content (LNC) of elephant grass cv. Roxo remained within the recommended range from day 36 DAC to harvest in the rainy season. For the transition and dry seasons, LNC remained below the lower limit at all ages (Table 2) due to the fact that nitrogen fertilization took place only in the rainy season (Figure 1), resulting in low residual N availability as it is very dynamic in the soil-plant system.
From the age of 27, higher LNC and stem nitrogen content (SNC) were observed for the rainy season, intermediate contents for the transition season, and lower contents for the dry season, probably due to the combined effect of nutrient availability and negative water balance since the contact between N and root occurs predominantly through a mass flow process (SILVA et al., 2011SILVA, A.A.; COSTA, A.M.; LANA, R.M. Q.; LANA, A.M.Q. Absorção de micronutrientes em pastagem de Brachiaria decumbens, após aplicação de cama de peru e fontes minerais na fertilização. Bioscience Journal, v.27, n.1, p.41-48, 2011.; TEIXEIRA et al., 2011TEIXEIRA, L.A.J.; QUAGGIO, J.A.; MELLIS, E.V. Ganhos de eficiência fertilizante em bananeira sob irrigação e fertirrigação. Revista Brasileira de Fruticultura, v.33, p.272-278, 2011.), requiring appropriate matrix potential for this to happen (Table 2).
The leaf phosphorus content (LPC) of elephant grass cv. Roxo quantified for all ages and seasons was in accordance with the recommendations by Werner (1997)WERNER, J.C.; PAULINO, V.T.; CANTARELLA, H.; ANDRADE, N.O.; QUAGGIO, J.A. Forrageiras. In: RAIJ, B.; CANTARELLA, H.; QUAGGIO, J.A.; FURLANI, A.M.C. (Ed.). Recomendações de adubação e calagem para o Estado de São Paulo. 2.ed. Campinas: Instituto Agronômico, 1997. p.263-273 (Boletim Técnico, 100). for elephant grass (1 to 3 g kg-1), and was similar to the content observed by Santos et al. (2012)SANTOS, R.L.; AZEVEDO, V.M.; FREIRE, F.J.; ROCHA, A.T.; TAVARES, J.A.; FREIRE, M.B.G.S. Extração e eficiência de uso de nutrientes em capim-elefante na presença de gesso. Revista Brasileira de Ciência do Solo, v.36, p.497-505, 2012. (3.60 g kg-1 for elephant grass cv. Roxo), revealing the tolerance of elephant grass cv. Roxo to extended water deficit regarding the absorption of P (Table 2).
From day 54 DAC there was progressive increase in LPC of elephant grass cv. Roxo between rainy, transition, and dry seasons due to the concentration effect of P on foliar biomass in response to the reduction in plant growth (COSTA et al., 2013COSTA, N.L.; MORAES, A; CARVALHO, P.C.F.; MONTEIRO, A.L.G.; MOTTA, A.C.V.; OLIVEIRA, R.A. Composição e extração de nutrientes por Trachypogon plumosus sob adubação e idades de rebrota. Archivos de Zootecnia, v.62, n.238, p.227-238, 2013.)
The stem phosphorus content (SPC) was not influenced by the growth age, presenting mean values of 5.25, 6.28, and 6.48 g kg-1 for the rainy, transition, and dry seasons, respectively (Table 2). On average, lower SPC was observed in the rainy season in relation to the other seasons, probably due to the effect of nutrient dilution as a consequence of the greater relative growth of the stem regarding the absorption of phosphorus in the rainy season (CARMO et al., 2011CARMO, G.A.; OLIVEIRA, F.R.A.; MEDEIROS, J.F.; OLIVEIRA, F.A.; M.S. CAMPOS; FREITAS D.C. Teores foliares, acúmulo e partição de macronutrientes na cultura da abóbora irrigada com água salina. Revista Brasileira de Engenharia Agrícola e Ambiental, v.15, n.5, p.512-518, 2011.).
The lack of significance of SPC between the transition and dry seasons reinforces that the forage was able to absorb and maintain similar P contents during these two cycles of regrowth under low availability of water in the soil. Although water deficit compromises contact and nutrient absorption, absorption may occur depending on the water stress time, as observed by Leão et al. (2011)LEÃO, D.A.S.; FREIRE, A L O. MIRANDA, J.R.P. Estado nutricional de sorgo cultivado sob estresse hídrico e adubação fosfatada. Pesquisa Agropecuária Tropical, v.41, n.1, p.74-79, 2011..
It was found that the leaf potassium content (LKC) of elephant grass cv. Roxo, at all ages and seasons, remained above the upper limit of the sufficiency range (15-30 g kg-1) recommended by Werner (1997)WERNER, J.C.; PAULINO, V.T.; CANTARELLA, H.; ANDRADE, N.O.; QUAGGIO, J.A. Forrageiras. In: RAIJ, B.; CANTARELLA, H.; QUAGGIO, J.A.; FURLANI, A.M.C. (Ed.). Recomendações de adubação e calagem para o Estado de São Paulo. 2.ed. Campinas: Instituto Agronômico, 1997. p.263-273 (Boletim Técnico, 100). and above the content observed by Silva et al. (2011)SILVA, A.A.; COSTA, A.M.; LANA, R.M. Q.; LANA, A.M.Q. Absorção de micronutrientes em pastagem de Brachiaria decumbens, após aplicação de cama de peru e fontes minerais na fertilização. Bioscience Journal, v.27, n.1, p.41-48, 2011. (27.07 g kg-1) for elephant grass cv. Roxo. The response in question may be related to a possible nutritional unbalance between N and K (PRADO et al., 2011PRADO, R.M.; HOJO, R.H.; AVALHÃES, C.C.; VALE, D.W.; PIMENTEL, U.V. Desempenho do capim-tanzânia cultivado em solução nutritiva com à omissão de macronutrientes. Scientia Agraria Paranaenis, v.10, n.1, p 58-68, 2011.) (Table 2).
The elephant grass yielded the highest LKC in the dry season, at all growth ages (Table 2). This was probably due to the negative water balance that occurred during the dry season (Figure 1), which induced a strategic accumulation of K in the leaves as an adaptive response to stress by water deficit (MAHMOOD et al., 2009MAHMOOD, T.; ASHRAF, M.; SHAHBAZ, M. Does exogenous application of glycinebetaine as a presowing seed treatment improve growth and regulate some key physiological attributes in wheat plants grown under water déficit conditions? Pakistan Journal Botany, v.41, n.3, p.1291-1302, 2009.).
The stem potassium content (SKC) was not influenced by the growth age, presenting mean values of 95.03, 82.06, and 98.32 g kg-1 for the rainy, transition, and dry seasons, respectively. The mean SKC did not differ (p>0.05) between the rainy and dry seasons, both being higher than the transition season. The highest SKC being in elephant grass cv. Roxo cultivated in the rainy and dry seasons may have occurred due to the effect of excessive absorption "luxury consumption" and concentration, respectively (Table 2).
The leaf calcium content (LCaC) of the elephant grass cv. Roxo observed was below the sufficiency values (3 to 8 g kg-1; WERNER, 1997WERNER, J.C.; PAULINO, V.T.; CANTARELLA, H.; ANDRADE, N.O.; QUAGGIO, J.A. Forrageiras. In: RAIJ, B.; CANTARELLA, H.; QUAGGIO, J.A.; FURLANI, A.M.C. (Ed.). Recomendações de adubação e calagem para o Estado de São Paulo. 2.ed. Campinas: Instituto Agronômico, 1997. p.263-273 (Boletim Técnico, 100).) and those observed by Santos et al. (2012)SANTOS, R.L.; AZEVEDO, V.M.; FREIRE, F.J.; ROCHA, A.T.; TAVARES, J.A.; FREIRE, M.B.G.S. Extração e eficiência de uso de nutrientes em capim-elefante na presença de gesso. Revista Brasileira de Ciência do Solo, v.36, p.497-505, 2012. (16.11 g kg-1) for elephant grass cv. Roxo, except for at ages 27 and 36 DAC for the transition and dry seasons, respectively (Table 3). It should be noted that there was no need to correct soil fertility for Ca; however, potassium fertilization possibly compromised Ca uptake, as reported by Vale et al. (2011)VALE, D.W.; PRADO, R.M.; AVALHÃES, C.C.; HOJO, R.H. Omissão de macronutrientes na nutrição e no crescimento da cana-de-açúcar cultivada em solução nutritiva. Revista Brasileira de Ciências Agrárias, v.6, n.2, p.189-196, 2011., since both minerals compete for the same sites of absorption in the root system.
The LCaC of elephant grass cv. Roxo varied between growing seasons from the age of 18 DAC to harvest. Higher LCaC was observed in the dry season (p<0.05) from age 45 DAC to harvest. Possibly, this concentration effect in LCaC was due to the lower production of biomass in the dry season (Figure 2).
The stem calcium content (SCaC) of the elephant grass cv. Roxo differed between seasons for all evaluated ages (Table 3), this was possibly because of the low translocation of this nutrient in the plant and because the Ca absorption by the root system was conditioned to rainfall, which was quite irregular during the evaluated seasons (Figure 1).
The leaf magnesium content (LMgC) of elephant grass cv. Roxo presented values compatible with the sufficiency range of 1.5 to 5.0 g kg-1 (WERNER, 1997WERNER, J.C.; PAULINO, V.T.; CANTARELLA, H.; ANDRADE, N.O.; QUAGGIO, J.A. Forrageiras. In: RAIJ, B.; CANTARELLA, H.; QUAGGIO, J.A.; FURLANI, A.M.C. (Ed.). Recomendações de adubação e calagem para o Estado de São Paulo. 2.ed. Campinas: Instituto Agronômico, 1997. p.263-273 (Boletim Técnico, 100).) at all ages and growing seasons, and it was similar to the content reported by Santos et al. (2012)SANTOS, R.L.; AZEVEDO, V.M.; FREIRE, F.J.; ROCHA, A.T.; TAVARES, J.A.; FREIRE, M.B.G.S. Extração e eficiência de uso de nutrientes em capim-elefante na presença de gesso. Revista Brasileira de Ciência do Solo, v.36, p.497-505, 2012. (1.60 g kg-1) for elephant grass cv. Roxo. Thus, it was observed that, despite the negative water balance, the plant was able to maintain the adequate LMgC (Table 3).
The LMgC and stem magnesium content (SMgC) differed between seasons from the ages of 18 and 27 DAC to harvest, respectively. From age 45 DAC to harvest, elephant grass cv. Roxo presented the lowest LMgC and SMgC in the rainy and transition seasons, probably due to the effect of nutrient dilution in the biomass produced (BATISTA & MONTEIRO, 2010BATISTA, K.; MONTEIRO, F.A. Variações nos teores de potássio, cálcio e magnésio em capim-marandu adubado com doses de nitrogênio e de enxofre. Revista Brasileira de Ciência do Solo, v.34, p.151-161, 2010.).
The leaf sulfur content (LSC) was below the lower limit of the sufficiency range (1 to 3 g kg-1; WERNER, 1997WERNER, J.C.; PAULINO, V.T.; CANTARELLA, H.; ANDRADE, N.O.; QUAGGIO, J.A. Forrageiras. In: RAIJ, B.; CANTARELLA, H.; QUAGGIO, J.A.; FURLANI, A.M.C. (Ed.). Recomendações de adubação e calagem para o Estado de São Paulo. 2.ed. Campinas: Instituto Agronômico, 1997. p.263-273 (Boletim Técnico, 100).) and similar to the content (5.12 g kg-1) observed by Silva et al. (2011)SILVA, A.A.; COSTA, A.M.; LANA, R.M. Q.; LANA, A.M.Q. Absorção de micronutrientes em pastagem de Brachiaria decumbens, após aplicação de cama de peru e fontes minerais na fertilização. Bioscience Journal, v.27, n.1, p.41-48, 2011. for elephant grass cv. Roxo, at all ages and growing seasons. Possibly, the amounts of S supplied via phosphate fertilization (single super phosphate, 8% S) and micronutrient sorted complex (FTE BR- 12) were not sufficient to meet the demand for S of elephant grass cv. Roxo. For the transition and dry seasons, some nutritional unbalance between N and S may have occurred (AVALHÃES et al., 2009AVALHÃES, C.C.; PRADO, R.M.; ROZANE, D.E.; ROMUALDO, L.M.; CORREIA, M.A.R. Omissão de macronutrientes no crescimento e no estado nutricional de capim-elefante (cv. Mott) cultivado em solução nutritiva. Scientia Agraria, v.10, n.3, p.215-222, 2009.; HOJO et al., 2012HOJO, R.H.; AVALHÃES, C.C.; VALE, D.W.; PRADO R.M.; PIMENTEL, U.V. Desempenho do capim-tanzânia cultivado em solução nutritiva com a omissão de macronutrientes. Scientia Agraria, v.10, n.2, p.63-73, 2012.). LSC and stem sulfur content (SSC) presented oscillation between the growth ages for all the growing seasons, showing no perceptible pattern between the seasons. An increasing response (p<0.001) was observed for the accumulation of macronutrients as a function of the growth age in the leaf, stem, and shoot components of the elephant grass cv. Roxo in the three growing seasons (rainy, transition and dry), except for the accumulation of Ca in the leaf and shoot for the transition season (Tables 4 and 5). Variation (p<0.05) was observed in the magnitude of the accumulation of N, P, K, Ca, Mg, and S as a function of the season for all the components analyzed (leaf, stem, and shoot) (Tables 4 and 5).
Nitrogen, phosphorus, and potassium accumulation in elephant grass cv. Roxo at different growth ages and seasons
Calcium, magnesium, and sulfur accumulations in elephant grass cv. Roxo at different growth ages in three growing seasons
Daily leaf nitrogen accumulation rates (LNA) were 1.66, 0.42, and 0.12 kg ha-1 day-1 for rainy, transition, and dry seasons, respectively. As for stem nitrogen accumulation (SNA), daily accumulations of 1.20, 0.14, and 0.01 kg ha-1 day-1 were estimated for rainy, transition, and dry seasons, respectively. From 45 DAC to harvest, the LNA and SNA presented higher values in the rainy season, intermediate values in the transition season, and lower values in the dry season (Table 3).
Nitrogen was the second most accumulated nutrient in the biomass of the shoot of elephant grass in the rainy and transition seasons, and the third in the dry season. The daily shoot nitrogen accumulation rate (SHNA) of the canopy was estimated at 2.87, 0.56, and 0.13 kg ha-1 day-1 for the rainy, transition, and dry seasons, respectively, resulting in accumulations of 139.50, 25.80, and 6.57 kg of N ha-1 at the age 63 DAC for rainy, transition, and dry seasons, respectively (Table 3). SHNA presented higher values in the rainy season, intermediate values in the transition season, and lower values in the dry season from age 45 DAC to harvest.
Daily leaf phosphorus accumulation rates (LPA) were estimated at 0.17, 0.11, and 0.07 kg ha-1 day-1 for the rainy, transition, and dry seasons, respectively. The stem phosphorus accumulation (SPA) presented values of 0.46, 0.18, and 0.06 kg ha-1 day-1 for the rainy, transition, and dry seasons, respectively. LPA and SPA presented, from age 27 and 45 DAC to harvest, respectively, higher values in the rainy season, intermediate values in the transition season, and lower values in the dry season.
Phosphorus was the third most accumulated nutrient in the shoot of the canopy in the rainy and transition seasons, and the second in the dry season (Table 3). Daily shoot phosphorus accumulation rates (SHPA) of 0.64, 0.30, and 0.13 kg ha-1 day-1 were estimated for the rainy, transition, and dry seasons, respectively, resulting in accumulations of 33.72, 15.73, and 8.30 kg ha-1 at age 63 DAC in the rainy, transition, and dry seasons, respectively. SHPA presented higher values in the rainy season, intermediate values in the transition season, and lower values in the dry season from age 45 DAC to harvest.
Daily leaf potassium accumulation rates (LKA) were estimated at 4.02, 2.16, and 1.11 kg ha-1 day-1 for rainy, transition, and dry seasons, respectively. For daily stem potassium accumulation rates (SKA), accumulations of 9.48, 2.84, and 1.08 kg ha-1 day-1 for the rainy, transition and dry seasons were estimated, respectively (Table 3). LKA and SKA presented higher values in the rainy season, intermediate values in the transition season, and lower values in the dry season from age 45 DAC to harvest.
Potassium was the most accumulated nutrient in the shoot of the canopy in the three studied seasons. Daily shoot potassium accumulation rates (SHKA) were estimated at13.51, 5.00, and 2.19 kg ha-1 day-1 in the rainy, transition, and dry seasons, respectively, resulting in accumulations of 675.16, 251.08, and 132.20 kg ha-1 at age 63 DAC, for the rainy, transition, and dry seasons, respectively. The SHKA presented higher values in the rainy season, intermediate values in the transition season, and lower values in the dry season from age 45 DAC to harvest.
Daily leaf calcium accumulation rates (LCaA) of the elephant grass cv. Roxo for the rainy and dry seasons were estimated at 0.10 and 0.08 kg ha-1 day-1, respectively (Table 5). In the transition season, the LCaA adjusted to the quadratic model, estimated a maximum accumulation of 4.60 kg ha-1 at 55.18 DAC. Daily stem calcium accumulation rates (SCaA) were estimated at 0.049, 0.015, and 0.011 kg ha-1 day-1 for rainy, transition, and dry seasons, respectively. Calcium was the fifth most accumulated nutrient in the shoot of the canopy in the rainy and transition seasons and the fourth in the dry season. Daily shoot calcium accumulation rates (SHCaA) of the canopy were estimated at 0.15 and 0.09 kg ha-1 day-1 for the rainy and dry seasons, respectively, resulting in accumulations of 5.98 and 4.46 kg ha-1 at age 63 DAC for the rainy and dry seasons, respectively. In the transition season, the SHCaA of the elephant grass, adjusted to the quadratic model, estimated a maximum accumulation of 4.64 kg ha-1 at 59.76 DAC. Considering the growing seasons, LCaA, SCaA, and SHCaA varied according to the growth age of the plant (Table 5).
The daily leaf magnesium accumulation rates (LMgA) were estimated at 0.17, 0.08, and 0.06 kg ha-1 day-1 for the rainy, transition, and dry seasons, respectively. The daily stem magnesium accumulation rates (SMgA) were estimated at 0.21, 0.04, and 0.02 kg ha-1 day-1 for rainy, transition, and dry seasons, respectively. Magnesium was the fourth most accumulated nutrient in the shoot of the elephant grass cv. Roxo in the rainy and transition seasons, and the fifth in the dry season. Daily shoot magnesium accumulation rates (SHMgA) were quantified as 0.38, 0.13, and 0.08 kg ha-1 day-1, for the rainy, transition, and dry seasons, respectively, resulting in accumulations of 20.86, 7.47, and 5.01 kg ha-1 day-1 at 63 DAC for the rainy, transition, and dry seasons, respectively. Considering the growing seasons, it was found that LMgA, SMgA, and SHMgA presented higher values in the rainy season, intermediate values in the transition season, and lower values in the dry season from age 45 DAC to harvest.
The daily leaf sulfur accumulation rates (LSA) of elephant grass cv. Roxo were estimated at 0.038, 0.008, and 0.001 kg ha-1 day-1 for the rainy, transition, and dry seasons, respectively. The daily stem sulfur accumulation rates (SSA) were estimated at 0.063, 0.006, and 0.002 kg ha-1 day-1for the rainy, transition, and dry seasons, respectively. Sulfur was the sixth most accumulated nutrient in the shoot the canopy of elephant grass cv. Roxo in the three growing seasons. The daily shoot sulfur accumulation rates (SHSA) of the canopy were estimated at 0.102, 0.014, and 0.004 kg ha-1 day-1 for the rainy, transition, and dry seasons, respectively, resulting in accumulations of 4.82, 0.914, and 0.272 kg ha-1 at age 63 DAC in the rainy, transition, and dry seasons, respectively.
Considering the growing seasons, it was found that the LSA varied according to age, while the SSA and SHSA presented higher values in the rainy season, intermediate values in the transition season, and lower values in the dry season from age 45 DAC to harvest.
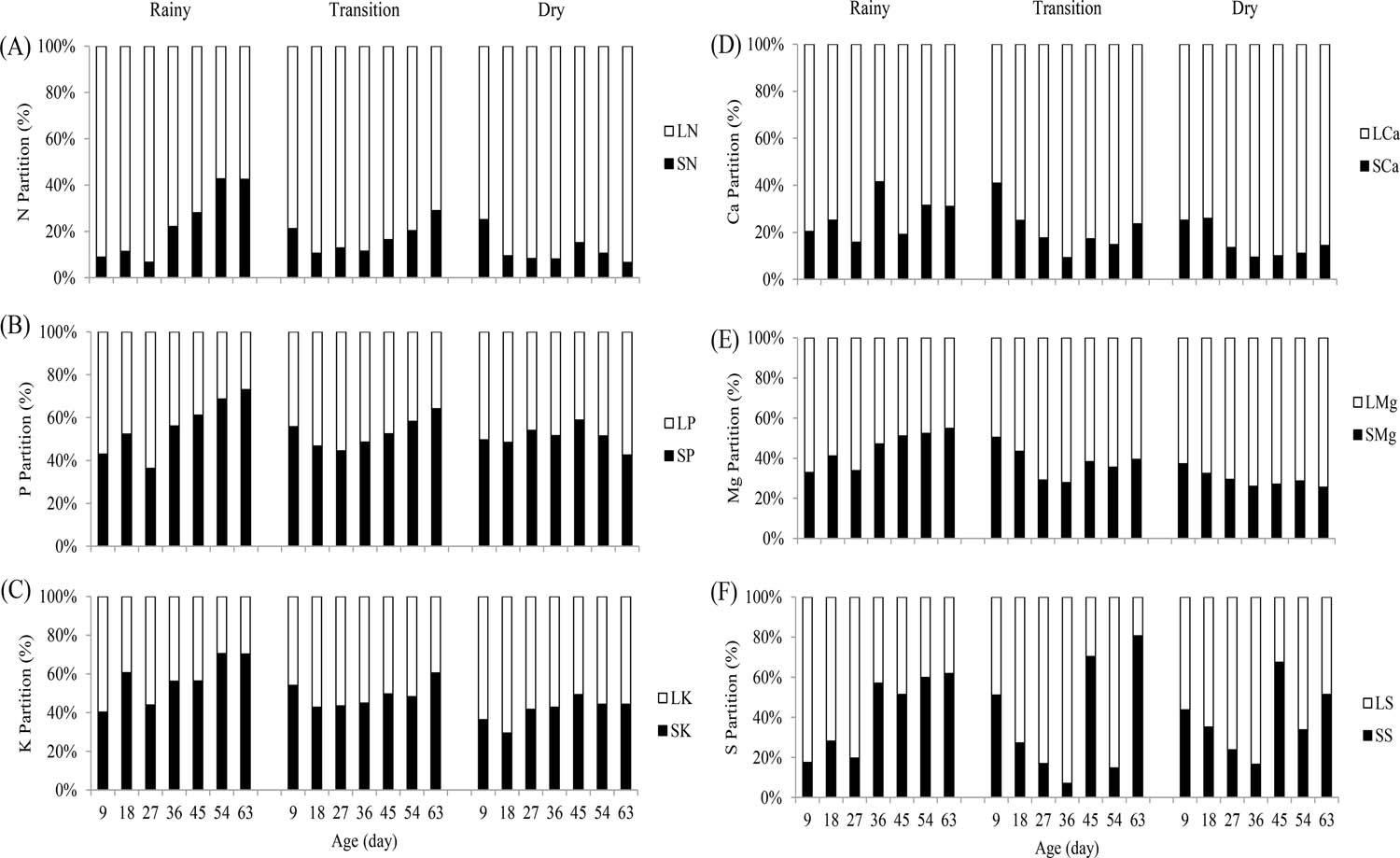
The elephant grass cv. Roxo partitioned more N to the leaf than to the stem, showing a more pronounced effect in the dry season in relation to the others (Figure 3A). The partition of nutrients in the plant is the product of the increase in biomass by the nutrient content in the plant tissue. Thus, the largest partition of N being observed in the leaf in the present study resulted from the higher LNC in relation to the SNC (Table 2), due to the function of nitrogen in plant metabolism, since it is a constituent of chlorophyll besides directly participating in photosynthesis, which occurs predominantly in the leaf (MARSCHNER, 2012MARSCHNER, P. Marchner's mineral nutrition of higher plants. 3th ed. New York: Academic Press, 2012. 651p.; LOPES et al., 2013bLOPES, M.N.; CANDIDO, M.J.D.; POMPEU, R.C.F.F.; SILVA, R.G.; CARVALHO, T.C.F.; SOMBRA, W.A.; MORAIS NETO, L.B.; PEIXOTO, M.J.A. Biomass flow in massai grass fertilized with nitrogen under intermittent stocking grazing with sheep. Revista Brasileira de Zootecnia, v.42, p.13-21, 2013b.).
Nitrogen, potassium, phosphorus, calcium, magnesium, and sulfur partitions in the shoot of elephant grass cv. Roxo
From the age 27 DAC to harvest in the rainy and transition seasons, the canopy partitioned more phosphorus to the stem (Figure 3B). The preferential allocation of P in the stem is because of the highest daily increase in stem biomass and the highest SPC in comparison to LPC (Table 2). As the plant grows there is a greater investment in stem biomass, allowing stem elongation in order to improve the distribution of photosynthetically active radiation inside the canopy (PARSONS et al., 2011PARSONS, A.; ROWARTH, J.; THORNLEY, J. AND NEWTON, P. Primary production of grasslands, herbage accumulation and use, and impacts of climate change. In: LEMAIRE, G.; HODGSON, J. E CHABBI, A. (Eds.) Grassland productivity and Ecosystem Services Wallingford, UK: CAB International, 2011. p.3-18.). With this, there is a preferential allocation of nutrients to the plant's stem, as observed in the present study.
In the rainy season, the canopy partitioned more P to the stem from age 36 DAC to harvest. For the transition and dry seasons, there was a certain isonomy in the partition of P between the canopy fractions; however, from the age 45 DAC to harvest in the dry season the plant partitioned more P to the leaf (Figure 3B). This response may be related to the increase in leaf area index from 1.90 to 2.70, observed between ages 54 and 63 DAC, and a consequent increase in leaf biomass, indicating that rainfall occurred at the end of the dry season (Figure 1), which stimulated plant growth and demanded a higher amount of P, as this is fundamental in energy metabolism and biomass production (OLIVEIRA et al., 2012OLIVEIRA, P.S.R.; DEMINICIS, B.B.; CASTAGNARA, D.D.; GOMES, F.C.N. Efeito da adubação com fósforo do capim-mombaça em solos com texturas arenosa e argilosa. Archivos de Zootecnia, v.61, n.235, p.397-406, 2012.; DIAS et al., 2015DIAS, D.G.; PEGORARO, R.F.; ALVES, D.D.; PORTO, E.M.V.; NETO, J.A.S.; ASPIAZÚ, I. Produção do capim Piatã submetido a diferentes fontes de fósforo. Revista Brasileira de Engenharia Agrícola e Ambiental, v.19, n.4, p.330-335, 2015.).
A larger partition of K was observed in the stem of elephant grass cv. Roxo in the rainy season, a result of the higher SKA in relation to LKA (Table 4), which was due to the biological role of K in the plant metabolism, such as to polarize the cells of the conducting vessels (MARSCHNER, 1995MARSCHNER, H. Mineral nutrition of higher plants. 2th ed. London: Academic Press, 1995. 889p.). For the transition season, a certain isonomy was observed for the partition of K. For the dry season, the canopy partitioned a greater amount of K to the leaf (Figure 3C), justifying the higher LKC in this season (Table 2).
A higher allocation of Ca was observed to the leaf than the shoot at all ages and seasons evaluated (Figure 3D), and that can be justified by the higher LCaC in relation to SCaC, probably due to the higher transpiratory rate of leaves. This is caused by transport of Ca in the plant occurring predominantly unidirectionally via conducting vessels of the xylem, allowing accumulation of this nutrient caused by the transpiratory flow of the organ in which it accumulates (MALAVOLTA et al., 1997MALAVOLTA, E.; VITTI, G.C.; OLIVEIRA, S.A. Avaliação do estado nutricional das plantas: princípios e aplicações. 2. Ed. Piracicaba: Associação Brasileira da Potassa e do Fosfato, 1997. 319 p.).
In the rainy season, the elephant grass cv. Roxo partitioned a larger amount of Mg to the stem from age 45 DAC to harvest (Figure 3E). At that age, a leaf/stem ratio (L/S) reduction was observed from 1.40 to 1.00, indicating a higher investment in supporting tissues present in the stem. Consequently, a higher amount of Mg was demanded as it is necessary for the formation of the middle lamella of plant tissues (PRADO, 2008PRADO, R.M. Nutrição de Plantas. 1.ed. São Paulo: UNESP/FUNESP. 2008; 408 p.). Also, from the age 27 DAC to harvest higher SMgC was observed in the rainy season (Table 4) in relation to the other seasons.
For the transition and dry seasons, the plant partitioned a larger amount of Mg to the leaf (Figure 3E). It should be noted that in these seasons the L/S ratio remained higher than 1.30 in all studied ages. Thus, the largest portion of leaves in the canopy demanded a higher amount of Mg, since it has a structural function as the central atom of the chlorophyll molecule, besides being the activator of several enzymes necessary for photosynthesis (PRADO, 2008PRADO, R.M. Nutrição de Plantas. 1.ed. São Paulo: UNESP/FUNESP. 2008; 408 p.).
Sulfur was partitioned in a greater amount to the stem from age 36 DAC to harvest (Figure 3F), accumulating on average 60.57% between the ages 36 and 63 DAC in the rainy season. For the transition and dry seasons, a partitioning of S similar to the rainy season was observed, changing only in the magnitude of the response. From age 9 to 36 DAC, a linear decrease of S was observed in the leaf (Figure 3F), which was also observed by Zobiole et al. (2010)ZOBIOLE, L.H.S.; CASTRO, C.; OLIVEIRA, F.A.; JUNIOR, A.O. Marcha de absorção de macronutrientes na cultura do girassol. Revista Brasileira de Ciência do Solo, v.34, p.425-433, 2010.. In the interval between 45 and 63 DAC, an allocation superior to 60% of S was observed in the stem of the shoot of the canopy.
Despite the accumulation rates of N, P, K, Mg, and S by elephant grass cv. Roxo showing positive linear behavior, it was noticed that the accumulation of those nutrients was greatest between the ages 27 and 36 DAC in the rainy season and between the ages 36 and 45 DAC for the transition season. The response in the present study is justified by the maximum population density of tillers being observed at 31 and 39 DAC in the rainy and transition seasons, respectively, as observed by Maranhão et al. (2018)MARANHÃO, T.D.; CÂNDIDO, M.J.D.; LOPES, M.N.; POMPEU, R.C.F.F.; CARNEIRO, M.S.S.; FURTADO, R.N.; SILVA, R.R.; SILVEIRA, F.G.A. Biomass componentes of Pennisetum purpureum cv. Roxo managed at different growth ages and seasons. Revista Brasileira de Saúde e Produção Animal [online], v.19, n.1, p.11-22, 2018.. As for the dry season, the accumulation of nutrients was enhanced between the ages of 18 and 27 DAC. Possibly, this pronounced accumulation at the beginning of the growth cycle was due to the mobilization of organic reserves presents in the rhizomes of elephant grass.
The macronutrient accumulations and contents in the biomass of the shoot of elephant grass cv. Roxo were modified according to the growing age and season. The accumulation of macronutrients in elephant grass cv. Roxo showed the following decreasing order: K > N > P > Mg > Ca > S for the rainy and transition seasons and K > P > N > Ca > Mg > S for the dry season. The age with the greatest nutrient accumulation varied according to the growing season of elephant grass cv. Roxo.
The accumulation pattern of macronutrients by elephant grass cv. Roxo along the growth cycle allows recommendation of maintenance fertilization with the nutrients N, P, K, Ca, Mg, and S at the beginning (18 days after cutting) and in the middle phase of the growth cycle, between the ages 27 and 36 days after cutting during the rainy season.
REFERENCES
- AVALHÃES, C.C.; PRADO, R.M.; ROZANE, D.E.; ROMUALDO, L.M.; CORREIA, M.A.R. Omissão de macronutrientes no crescimento e no estado nutricional de capim-elefante (cv. Mott) cultivado em solução nutritiva. Scientia Agraria, v.10, n.3, p.215-222, 2009.
- BATISTA, K.; MONTEIRO, F.A. Variações nos teores de potássio, cálcio e magnésio em capim-marandu adubado com doses de nitrogênio e de enxofre. Revista Brasileira de Ciência do Solo, v.34, p.151-161, 2010.
- BHERING, M.; CABRAL, L.S.; ABREU, J.G.; SOUZA, A.L.; ZERVOUDAKIS, J.T.; RODRIGUES, R.C.; PEREIRA, G.A.C.; REVERDITO, R.; OLIVEIRA, Í. S. Características agronômicas do capim- elefante roxo em diferentes idades de corte na Depressão Cuiabana. Revista Brasileira de Saúde e Produção Animal [online], v.9, n.3, p.384-396, 2008.
- CARMO, G.A.; OLIVEIRA, F.R.A.; MEDEIROS, J.F.; OLIVEIRA, F.A.; M.S. CAMPOS; FREITAS D.C. Teores foliares, acúmulo e partição de macronutrientes na cultura da abóbora irrigada com água salina. Revista Brasileira de Engenharia Agrícola e Ambiental, v.15, n.5, p.512-518, 2011.
- COMISSÃO DE FERTILIDADE DO SOLO DO ESTADO DE MINAS GERAIS - CFSEMG. Recomendações para uso de corretivos e fertilizantes em Minas Gerais – 5a Aproximação. 5.ed. Viçosa, MG: Universidade Federal de Viçosa, 1999. 359p.
- COSTA, K.A.P.; OLIVEIRA, I.P.; SEVERIANO, E.C.; SAMPAIO, F.M.T.; CARRIJO, M.S.; RODRIGUES, C.R. Extração de nutrientes pela fitomassa de cultivares de brachiaria brizantha sob doses de nitrogênio. Ciência Animal Brasileira, v.11, n.2, p.307-314, 2010.
- COSTA, N.L.; MORAES, A; CARVALHO, P.C.F.; MONTEIRO, A.L.G.; MOTTA, A.C.V.; OLIVEIRA, R.A. Composição e extração de nutrientes por Trachypogon plumosus sob adubação e idades de rebrota. Archivos de Zootecnia, v.62, n.238, p.227-238, 2013.
- DIAS, D.G.; PEGORARO, R.F.; ALVES, D.D.; PORTO, E.M.V.; NETO, J.A.S.; ASPIAZÚ, I. Produção do capim Piatã submetido a diferentes fontes de fósforo. Revista Brasileira de Engenharia Agrícola e Ambiental, v.19, n.4, p.330-335, 2015.
- ECHER, F.R.; DOMINATO, J.C.; CRESTE, J.E. Absorção de nutrientes e distribuição da massa fresca e seca entre órgãos de batata-doce. Horticultura Brasileira, v.27, n.2, p.176-182, 2009.
- HOJO, R.H.; AVALHÃES, C.C.; VALE, D.W.; PRADO R.M.; PIMENTEL, U.V. Desempenho do capim-tanzânia cultivado em solução nutritiva com a omissão de macronutrientes. Scientia Agraria, v.10, n.2, p.63-73, 2012.
- KÖPPEN, W. Das geographisca System der Klimate Gebr, Borntraeger, 1936. 1-44p.
- LEÃO, D.A.S.; FREIRE, A L O. MIRANDA, J.R.P. Estado nutricional de sorgo cultivado sob estresse hídrico e adubação fosfatada. Pesquisa Agropecuária Tropical, v.41, n.1, p.74-79, 2011.
- LOPES, M.N.; CÂNDIDO, M.J.D.; POMPEU, R.C.F.F.; SILVA, R.G.; LOPES, J.W.B.; FERNANDES, F.R.B.; LACERDA, C.L.; BEZERRA, F.M.L. Fluxo de biomassa em capim-massai durante o estabelecimento e rebrotação com e sem adubação nitrogenada. Revista Ceres, v.60, n.3, p.363-371, 2013a.
- LOPES, M.N.; CANDIDO, M.J.D.; POMPEU, R.C.F.F.; SILVA, R.G.; CARVALHO, T.C.F.; SOMBRA, W.A.; MORAIS NETO, L.B.; PEIXOTO, M.J.A. Biomass flow in massai grass fertilized with nitrogen under intermittent stocking grazing with sheep. Revista Brasileira de Zootecnia, v.42, p.13-21, 2013b.
- MAHMOOD, T.; ASHRAF, M.; SHAHBAZ, M. Does exogenous application of glycinebetaine as a presowing seed treatment improve growth and regulate some key physiological attributes in wheat plants grown under water déficit conditions? Pakistan Journal Botany, v.41, n.3, p.1291-1302, 2009.
- MALAVOLTA, E.; VITTI, G.C.; OLIVEIRA, S.A. Avaliação do estado nutricional das plantas: princípios e aplicações. 2. Ed. Piracicaba: Associação Brasileira da Potassa e do Fosfato, 1997. 319 p.
- MARANHÃO, T.D.; CÂNDIDO, M.J.D.; LOPES, M.N.; POMPEU, R.C.F.F.; CARNEIRO, M.S.S.; FURTADO, R.N.; SILVA, R.R.; SILVEIRA, F.G.A. Biomass componentes of Pennisetum purpureum cv. Roxo managed at different growth ages and seasons. Revista Brasileira de Saúde e Produção Animal [online], v.19, n.1, p.11-22, 2018.
- MARSCHNER, H. Mineral nutrition of higher plants 2th ed. London: Academic Press, 1995. 889p.
- MARSCHNER, P. Marchner's mineral nutrition of higher plants 3th ed. New York: Academic Press, 2012. 651p.
- OLIVEIRA, P.S.R.; DEMINICIS, B.B.; CASTAGNARA, D.D.; GOMES, F.C.N. Efeito da adubação com fósforo do capim-mombaça em solos com texturas arenosa e argilosa. Archivos de Zootecnia, v.61, n.235, p.397-406, 2012.
- PARSONS, A.; ROWARTH, J.; THORNLEY, J. AND NEWTON, P. Primary production of grasslands, herbage accumulation and use, and impacts of climate change. In: LEMAIRE, G.; HODGSON, J. E CHABBI, A. (Eds.) Grassland productivity and Ecosystem Services Wallingford, UK: CAB International, 2011. p.3-18.
- PRADO, R.M. Nutrição de Plantas 1.ed. São Paulo: UNESP/FUNESP. 2008; 408 p.
- PRADO, R.M.; HOJO, R.H.; AVALHÃES, C.C.; VALE, D.W.; PIMENTEL, U.V. Desempenho do capim-tanzânia cultivado em solução nutritiva com à omissão de macronutrientes. Scientia Agraria Paranaenis, v.10, n.1, p 58-68, 2011.
- SANTOS, R.L.; AZEVEDO, V.M.; FREIRE, F.J.; ROCHA, A.T.; TAVARES, J.A.; FREIRE, M.B.G.S. Extração e eficiência de uso de nutrientes em capim-elefante na presença de gesso. Revista Brasileira de Ciência do Solo, v.36, p.497-505, 2012.
- SILVA, A.A.; COSTA, A.M.; LANA, R.M. Q.; LANA, A.M.Q. Absorção de micronutrientes em pastagem de Brachiaria decumbens, após aplicação de cama de peru e fontes minerais na fertilização. Bioscience Journal, v.27, n.1, p.41-48, 2011.
- SILVA, F.C. Manual de análises químicas de solos, plantas e fertilizantes 2.ed. Brasília, DF: Embrapa informação tecnológica / Embrapa solos, 2009. 627p.
- SILVA, M.R.; PELISSARI, A.; MORAES, A.; SANDINI, I.E.; CASSOL, L.C.; ASSMANN, T.S.; OLIVEIRA, E.B. Acumulação de nutrientes e produção forrageira de aveia e azevém em função da aplicação de calcário e gesso em superfície. Revista de Ciências Agrárias, v.38, n.3, p.346-356, 2015.
- TEIXEIRA, L.A.J.; QUAGGIO, J.A.; MELLIS, E.V. Ganhos de eficiência fertilizante em bananeira sob irrigação e fertirrigação. Revista Brasileira de Fruticultura, v.33, p.272-278, 2011.
- UNIVERSIDADE FEDERAL DE VIÇOSA - UFV. SAEG - sistema para análises estatísticas Versão 9.1. Viçosa, MG: Fundação Arthur Bernardes, 2007.
- VALE, D.W.; PRADO, R.M.; AVALHÃES, C.C.; HOJO, R.H. Omissão de macronutrientes na nutrição e no crescimento da cana-de-açúcar cultivada em solução nutritiva. Revista Brasileira de Ciências Agrárias, v.6, n.2, p.189-196, 2011.
- WERNER, J.C.; PAULINO, V.T.; CANTARELLA, H.; ANDRADE, N.O.; QUAGGIO, J.A. Forrageiras. In: RAIJ, B.; CANTARELLA, H.; QUAGGIO, J.A.; FURLANI, A.M.C. (Ed.). Recomendações de adubação e calagem para o Estado de São Paulo 2.ed. Campinas: Instituto Agronômico, 1997. p.263-273 (Boletim Técnico, 100).
- ZOBIOLE, L.H.S.; CASTRO, C.; OLIVEIRA, F.A.; JUNIOR, A.O. Marcha de absorção de macronutrientes na cultura do girassol. Revista Brasileira de Ciência do Solo, v.34, p.425-433, 2010.
ERRATA
-
In the article "Accumulation and partition of macronutrients in Pennisetum purpureum cv. Roxo managed under different growth ages and seasons", with the number DOI: 10.1590/S1519-99402018000300001, published in the journal: Brazilian Journal of Animal Health and Production, volume 19; number 3; pg. 223-240, page 235, include the captions "LCa, SCa; LMg, SMg; LS, SS", as shown in the figure below:
Publication Dates
-
Publication in this collection
Jul-Sep 2018
History
-
Received
23 Jan 2018 -
Accepted
16 July 2018