Resumos
De abril a maio de 2000 a 2002 oito localidades com remanescentes de cerrados sensu stricto no estado de São Paulo foram amostradas para o levantamento das espécies de asteráceas, uma das famílias mais representativas da flora herbáceo-arbustiva nestas formações. Foram feitas 23 amostragens e cada área de estudo foi amostrada em média uma vez por ano durante o pico de floração das plantas. Ao todo foram obtidas 399 amostras, nas quais foram reconhecidas 89 morfoespécies (74 foram identificadas como espécies conhecidas). Quarenta por cento das espécies foram registradas uma única vez (unicatas), indicando um grande número de espécies raras. Apenas 10% das espécies que ocorreram em mais de uma amostra foram obtidas de uma mesma área (sobreposição espacial) ou de um mesmo ano de estudo (sobreposição temporal). A riqueza de espécies em cada área foi estimada por meio de transeções e depois comparada à riqueza total observada em cada área de estudo, sendo esta na maioria das vezes mais alta que a estimada com base nas transeções. A lista de espécies obtida para os cerrados amostrados foi comparada a outras 24 listas publicadas para cerrados no Brasil. Embora a maioria das espécies mais comuns tenha coincidido, oito espécies (11% das espécies identificadas) não constam das listas publicadas. Concluímos que as áreas de cerrado sensu stricto estudadas no estado de São Paulo encontram-se isoladas, com uma grande parte da flora herbáceo-arbustiva composta por várias espécies raras e exclusivas. Diante deste quadro, sugerimos que a manutenção da biodiversidade de Asteraceae depende da conservação de todo o conjunto de remanescentes de cerrado do estado de São Paulo.
Asteraceae; cerrado; biodiversidade; São Paulo; Brasil
Eight "cerrado" (savanna) fragments in the State of São Paulo (Brazil) were surveyed from 2000 to 2002 to produce an inventory of Asteraceae species. This family is one of the most important in the herbaceous and shrubby components at cerrados. Each fragment was sampled on average once a year during the plants' peak flowering period, using a fixed number of randomly placed transects. We obtained 399 samples and recognized 89 morphospecies (of which 74 were identified to species). Forty percent of the species were encountered only once (unicates). Only ten percent of the species that occurred in more than a sample were recorded again in the same fragment or in the same year. Species richness was estimated from frequencies and abundances within transects, and then compared to the observed total richness in the area, which was most of the times higher than the estimates based on transects. Our species list was then compared to other 24 cerrado species lists in the literature. Although most of our commoner species were recorded in published lists, eight species were not found in any. We conclude that the studied cerrado fragments in São Paulo State are isolated and fragile areas composed of many rare and exclusive herbaceous and shrubby species. We suggest that the maintenance of current diversity of Asteraceae species depends on maintaining and preserving all cerrado fragments in the State.
Asteraceae; cerrado; savanna; biodiversity; São Paulo; Brazil
ARTIGOS
Diversidade e ocorrência de Asteraceae em cerrados de São Paulo
Adriana Monteiro de AlmeidaI,1 1 Autor Correspondente: amdalmeida@yahoo.com ; Carlos Roberto FonsecaII; Paulo Inácio PradoI,III; Mário Almeida-NetoI; Soraia DinizIV; Umberto KubotaI; Marina Reiter BraunI; Rafael L. Galdini RaimundoI; Luciano Alves dos AnjosI; Tehra Gomes MendonçaI; Silvia de Melo FutadaI; Thomas Michael LewinsohnI
ILaboratório de Interações Insetos-Plantas, Depto. Zoologia, IB, Unicamp. CP 6109, CEP 13083-970, Campinas, SP, Brasil. http://www.ib.unicamp.br/pesquisa/projetos/biota/inseto_planta/index.html
IIEndereço Atual: Laboratório de Interação Animal-Planta, UNISINOS, CEP 93022-000, São Leopoldo, RS, Brazil, www.saude.unisinos.br/laboratorios/interacao-animal-planta
IIINúcleo de Estudos e Pesquisas Ambientais (Nepam), Unicamp, Campinas, SP, Brasil
IVEndereço Atual: Depto. Botânica e Ecologia, IB, UFMT
RESUMO
De abril a maio de 2000 a 2002 oito localidades com remanescentes de cerrados sensu stricto no estado de São Paulo foram amostradas para o levantamento das espécies de asteráceas, uma das famílias mais representativas da flora herbáceo-arbustiva nestas formações. Foram feitas 23 amostragens e cada área de estudo foi amostrada em média uma vez por ano durante o pico de floração das plantas. Ao todo foram obtidas 399 amostras, nas quais foram reconhecidas 89 morfoespécies (74 foram identificadas como espécies conhecidas). Quarenta por cento das espécies foram registradas uma única vez (unicatas), indicando um grande número de espécies raras. Apenas 10% das espécies que ocorreram em mais de uma amostra foram obtidas de uma mesma área (sobreposição espacial) ou de um mesmo ano de estudo (sobreposição temporal). A riqueza de espécies em cada área foi estimada por meio de transeções e depois comparada à riqueza total observada em cada área de estudo, sendo esta na maioria das vezes mais alta que a estimada com base nas transeções. A lista de espécies obtida para os cerrados amostrados foi comparada a outras 24 listas publicadas para cerrados no Brasil. Embora a maioria das espécies mais comuns tenha coincidido, oito espécies (11% das espécies identificadas) não constam das listas publicadas. Concluímos que as áreas de cerrado sensu stricto estudadas no estado de São Paulo encontram-se isoladas, com uma grande parte da flora herbáceo-arbustiva composta por várias espécies raras e exclusivas. Diante deste quadro, sugerimos que a manutenção da biodiversidade de Asteraceae depende da conservação de todo o conjunto de remanescentes de cerrado do estado de São Paulo.
Palavras-chave: Asteraceae, cerrado, biodiversidade, São Paulo, Brasil.
ABSTRACT
Eight "cerrado" (savanna) fragments in the State of São Paulo (Brazil) were surveyed from 2000 to 2002 to produce an inventory of Asteraceae species. This family is one of the most important in the herbaceous and shrubby components at cerrados. Each fragment was sampled on average once a year during the plants' peak flowering period, using a fixed number of randomly placed transects. We obtained 399 samples and recognized 89 morphospecies (of which 74 were identified to species). Forty percent of the species were encountered only once (unicates). Only ten percent of the species that occurred in more than a sample were recorded again in the same fragment or in the same year. Species richness was estimated from frequencies and abundances within transects, and then compared to the observed total richness in the area, which was most of the times higher than the estimates based on transects. Our species list was then compared to other 24 cerrado species lists in the literature. Although most of our commoner species were recorded in published lists, eight species were not found in any. We conclude that the studied cerrado fragments in São Paulo State are isolated and fragile areas composed of many rare and exclusive herbaceous and shrubby species. We suggest that the maintenance of current diversity of Asteraceae species depends on maintaining and preserving all cerrado fragments in the State.
Key words: Asteraceae, "cerrado", savanna, biodiversity, São Paulo, Brazil.
1. Introdução
A área coberta por cerrados no Brasil representava originalmente cerca de 23% do seu território, estendendo-se por 11 estados e pelo Distrito Federal, sendo a segunda maior formação vegetal brasileira, após a Floresta Amazônica (Pinto 1994; Eiten 1994; Alho & Martins 1995; Ratter et al. 1997). Por sua localização central, está em contato com quatro outros biomas brasileiros, além de conter trechos das três maiores bacias hidrográficas da América do Sul (Pinto 1994; Alho & Martins 1995; Ratter et al. 1997; Silva & Bates 2002). Sua vasta extensão, aliada ao contato e conseqüente troca de espécies com outros biomas, propiciam ao Cerrado uma alta biodiversidade, estimada atualmente em 1/3 da biota brasileira e 5% da fauna e flora mundiais (Alho & Martins 1995).
O bioma Cerrado é um ambiente em mosaico, formado por diferentes fitofisionomias, com composição florística distintas: cerradão, cerrado sensu stricto e campos cerrados (Eiten 1971). Também estão incluídas no bioma matas de galeria, matas ciliares, carrascos e campos rupestres (Alho 1994; Eiten 1994; Ribeiro & Walter 1998; Batalha et al. 2001; Silva & Bates 2002). No cerrado sensu stricto, a fisionomia mais freqüente do Cerrado, o estrato arbóreo tem aparência característica, com troncos contorcidos com grossas cascas, e com folhas esclerófilas.
Principalmente a partir de 1970, o Cerrado foi progressivamente ocupado pela expansão agropecuária. As áreas de Cerrado se restringiram desde então cada vez mais a pequenos remanescentes, inseridos em uma matriz de culturas e pastos, muitas vezes vizinhos a cidades, aumentando cada vez mais a sua fragmentação (MMA/SBF 2002). Estima-se que em São Paulo atualmente o cerrado ocupe menos de 7% de sua extensão original e menos de 1% da área do estado (Kronka et al. 1998; Durigan 2003).
As espécies que compõem a flora e a fauna do cerrado não são igualmente estudadas, e alguns grupos são melhor conhecidos que outros. A flora arbórea é bem melhor conhecida que a flora arbustiva (por exemplo Mendonça et al. 1998) e herbácea (por exemplo Batalha 2001). Estudos florísticos extensivos, assim como compilações de vários trabalhos mostram que a cada novo inventário espécies ainda são adicionadas à lista de angiospermas do Cerrado, indicando o conhecimento ainda incipiente para este bioma brasileiro (por exemplo Ratter & Dargie 1992; Ratter et al. 1996; Mendonça et al. 1998; Batalha & Mantovani 2001, Batalha 2001; Ratter et al. 2003).
A família Asteraceae é a maior dentre as angiospermas (Bremer 1994), com grande importância no estrato herbáceo e arbustivo de Cerrados (Ratter et al. 1997; Batalha & Mantovani 2001). O presente estudo teve como objetivos inventariar a flora de Asteraceae em remanescentes de cerrados sensu stricto do estado de São Paulo; estimar a riqueza de espécies de Asteraceae em cada área; assim como listar as espécies obtidas e comparar esta lista com outros levantamentos florísticos de cerrados brasileiros. Este estudo se distingue de outros levantamentos semelhantes por vizar apenas espécies de Asteraceae e, desta forma, apresentar uma descrição mais detalhada para um grupo diversificado porém menos conhecido, por ser formado principalmente por espécies herbáceas e arbustivas. Mostramos que os remanescentes de cerrados estudados apresentam em geral uma alta riqueza de espécies de asteráceas, em grande parte únicas e exclusivas a estes fragmentos, reforçando a necessidade de mais estudos da flora herbáceo-subarbustiva, assim como da urgência na conservação destas áreas.
2. Material e Métodos
2.1. Amostragem
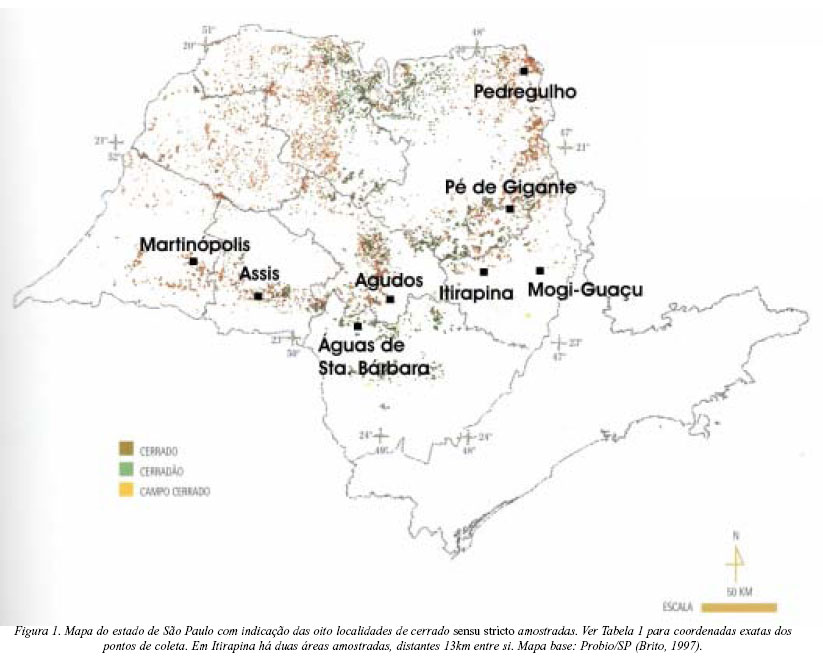
Ao todo foram feitas 23 amostragens em oito localidades de cerrados sensu stricto do estado de São Paulo (Tabela 1; Figura 1), de março a junho nos anos de 2000 a 2002, com uma maior concentração de coletas principalmente durante os meses de abril e maio, pico de floração de grande parte das espécies de asteráceas do cerrado. As áreas de cerrado amostradas foram selecionadas através de mapas (Brito 1997), assim como através da discussão com vários especialistas (ver agradecimentos). Em 2002 houve duas coletas no município de Itirapina: uma amostragem inicial em janeiro na mesma área de coleta dos anos anteriores e uma segunda amostragem em abril, efetuada em uma outra área próxima (13 km), pertencente à Estação Experimental de Itirapina (IEF SP). Esta mudança de área foi necessária pois a área de amostragem dos anos anteriores se mostrou bastante degradada, com presença de gado e perda de várias espécies devido a pisoteio excessivo. Como as análises do presente estudo são em escala geográfica, as duas áreas dentro do mesmo município foram consideradas uma mesma localidade.
Em cada área foi demarcada uma parcela de 300 x 30m de cerrado sensu stricto relativamente homogêneo, excluindo bordas e alterações fisionômicas evidentes. Em cada parcela foram sorteadas as posições de 15 transeções de 30 x 5m (150m2) e, em cada transeção registradas a presença e a abundância (até 10 indivíduos) de todos os morfotipos de Asteraceae em floração ou frutificação. As amostragens foram repetidas nas mesmas parcelas todos os anos, localizadas com a ajuda de um aparelho georeferenciador (GPS). Também foram registradas através de busca ativa, espécies de asteráceas em floração ou frutificação, fora das transeções, que foram observadas dentro e fora da parcela. Exsicatas de cada morfoespécie foram feitas como testemunho, assim como fotografias para documentar o hábito e detalhes de ramos floridos. O material botânico está depositado no herbário UEC (Unicamp).
2.2. Identificação das Espécies
A identificação das espécies coletadas foi baseada na literatura taxonômica e na comparação com exsicatas da coleção de referência já organizada pelo Projeto Temático "Diversidade de Espécies e de Interações em Plantas e Insetos Fitófagos". Vários especialistas (ver Agradecimentos) confirmaram e corrigiram as identificações de espécies. Grande parte das morfoespécies de Asteraceae coletadas (73 espécies, 82%) foi identificada até espécie.
Dentre os problemas de identificação das espécies, destacam-se alguns: (1) Vernonanthura membranacea (= Vernonia ruficoma Schltdl. ex Baker.) e Vernonia rubriramea Mart. ex DC. (= Vernonanthura) são muito semelhantes e de difícil distinção, e há dúvidas sobre sua classificação em espécies distintas (Harold Robinson, com. pess.). Neste caso, foram considerados como V. membranacea os indivíduos coletados em nosso estudo, e na comparação com listas publicadas consideramos Vernonia rubriramea e Vernonanthura membranacea como sinônimos. (2) Chromolaena odorata e Chromolaena maximilianii também são muito similares (Esteves, 2001; Harold Robinson, com. pess.). No presente estudo, designamos como C. odorata todos os indivíduos coletados e na comparação com outras listas mantivemos os nomes originais das publicações. (3) Chromolaena squalida e C. pedunculosa também são espécies muito similares e de distinção sutil embora consistente. Com o auxílio dos especialistas pudemos reconhecer as duas espécies, mas é possível que nas listas publicadas elas tenham sido confundidas. (4) Chromolaena subvelutina é uma nova combinação criada por Esteves (2001) reabilitando a espécie Eupatorium subvelutinum DC., que havia sido sinonimizada por King e Robinson (1987) em Chromolaena squalida. A distinção entre C. subvelutina e as espécies C. squalida e C. pedunculosa no material analisado é extremamente difícil, e apenas os espécimes identificados pelo especialista, Dr. Roberto L. Esteves como C. subvelutina foram mantidos. (5) Pterocaulon alopecuroides e P. virgatum são espécies semelhantes que também podem ter sido confundidas nas listas publicadas.
2.3. Análise dos Dados
A riqueza total esperada de espécies em cada área foi calculada por meio dos estimadores Chao1, Chao2 (por meio das fórmulas simplificadas), Jacknife1 e Jacknife2 (Figura 2) (Colwell & Coddington 1994) com o programa EstimateS v.7 (Colwell, 2004), onde F é a freqüência de "singletons" (F1), ou "doubletons" (F2), Q indica o número de espécies presentes em uma (Q1) e duas amostras (Q2), e m é o número total de amostras.
O estimador Chao1 usa o número de espécies representadas por um ("singleton") e dois ("doubleton") indivíduos no conjunto de amostras. Jackknife1 usa o número de espécies presentes em uma amostra. Chao2 e Jackknife2 usam o número de espécies presentes em uma (Q1) e duas amostras (Q2).
Quando as amostras não possuem "doubletons" (F2 em Chao1) ou duplicatas (Q2 em Chao2) a fórmula é indefinida. Nestes casos usamos as fórmulas completas (Figura 3) (Colwell 2004).
Os valores estimados foram então comparados ao número de espécies registradas na área, dentro e fora das transeções.
A relação entre o número de áreas em que cada espécie ocorreu e o número de estudos em que foi listada foi avaliada por meio do índice de correlação de Spearman (rS), e os resíduos avaliados para a identificação de espécies que desviam muito da tendência geral.
3. Resultados e Discussão
3.1 Distribuição Espacial e Temporal
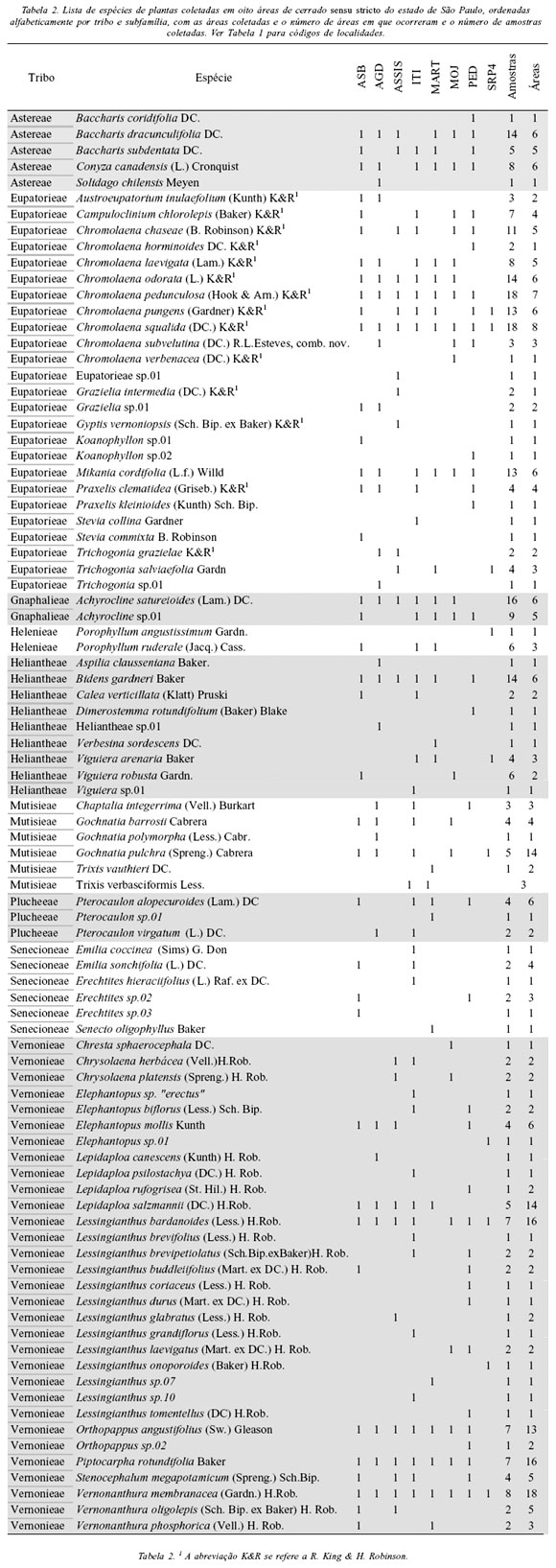
Foram amostradas 399 populações de Asteraceae nos cerrados de São Paulo, das quais foram identificadas 89 espécies / morfoespécies pertencentes a nove tribos (ou subfamília, no caso de Helenieae) (Tabela 2). As tribos com maior número de espécies foram Vernonieae, Eupatorieae e Heliantheae, que juntas somaram 73% das espécies obtidas (Figura 4). Estas proporções estão fortemente correlacionadas (r = 0,92, p = 0,0002, n = 9) com o percentual de espécies descritas para o Brasil em cada tribo (Barroso 1986). Tal correlação também ocorre para o conjunto de espécies que amostramos no sul e sudeste do Brasil (Prado et al. 2002).
Das 89 morfoespécies de Asteraceae, 74 foram identificadas até espécie (83%). Dentre as espécies mais freqüentes, destacam-se Chromolaena squalida e Vernonanthura membranacea, com os maiores números de amostras e ocorrentes em todas as localidades, seguidas por Chromolaena pedunculosa, Lessingianthus bardanoides, Orthopappus angustifolius e Piptocarpha rotundifolia, que só não ocorreram em uma localidade (Tabela 2). Uma análise das áreas amostradas em relação à sua flora de Asteraceae será publicada em separado.
Apesar de terem sido coletadas 89 espécies de Asteraceae, a maior riqueza obtida em uma só área foi 42, em Itirapina (Tabela 3). Um quinto das espécies foi registrado em cinco ou mais áreas (19 espécies ou 21%) e metade das espécies foi registrada em apenas uma das áreas amostradas (42 espécies ou 47%; Tabela 2 e Figura 5). Estes dados sugerem um alto revezamento de espécies (turnover) entre as áreas estudadas. Dentre as espécies amostradas em apenas uma área, 86% (36 espécies) ocorreram em somente uma amostra, ou seja, foram encontradas em apenas uma ocasião.
O fato de uma grande proporção das espécies ser encontrada em apenas uma localidade já foi descrito para asteráceas em áreas de campos rupestres. Na Serra da Mantiqueira um levantamento considerando apenas a tribo Eupatorieae detectou que 39% das espécies estavam restritas a apenas uma localidade, das quais 36% ocorreram em apenas uma amostra (Almeida, Prado & Lewinsohn 2004). Em levantamento da tribo Vernonieae na Serra do Espinhaço mineiro, usando a mesma metodologia do estudo anterior, cerca de 70% das espécies foram encontradas em apenas uma localidade (Prado & Lewinsohn 2000). Nesses inventários utilizou-se apenas a busca ativa de espécies em flor ou fruto, um dos métodos utilizados no presente trabalho.
A grande proporção de espécies geograficamente restritas, aqui observada para a flora de asteráceas em cerrados, parece ser um padrão comum também para o componente arbóreo deste bioma. Ratter e colaboradores (2003) observaram que dentre as 914 espécies presentes em 315 listas para cerrados "core", 309 foram unicatas e 305 foram muito raras. Da mesma forma, um segundo estudo usando o mesmo conjunto de dados (Bridgewater et al. 2004) dividiu as áreas de cerrados em seis províncias fitogeográficas e verificou que 52% das espécies foram restritas a uma província e apenas 3,9% das espécies ocorreram nas seis províncias. Durigan e colaboradores (2003) inventariaram 86 áreas de cerrados no estado de São Paulo e identificaram 554 espécies, na maioria lenhosas. Destas, 19% das espécies ocorreram em uma só área, e uma grande parte das espécies encontradas em mais de uma área tiveram distribuição restrita, enquanto apenas 10% das espécies ocorreram em mais de 50% das áreas (Durigan et al. 2003).
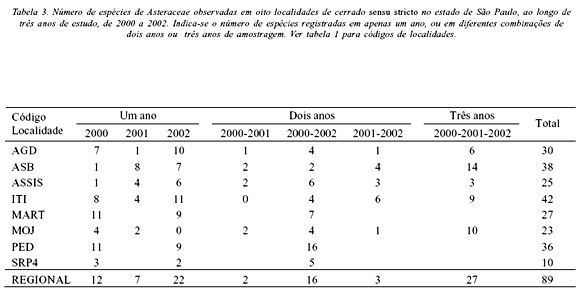
Também foi observada baixa similaridade de espécies entre anos de estudo. Cerca de metade das espécies foi registrada em apenas um ano, enquanto menos de um quarto das espécies foi registrado em apenas dois anos (Tabela 3). O mesmo padrão se repetiu localmente, levando-se em conta as cinco localidades com três anos de coletas. A única exceção foi a Reserva Biológica de Moji-Guaçu (MOJ), onde 43% das espécies foram registradas nos três anos de estudo (Tabela 3). Espécies que ocorreram em dois anos não foram observadas necessariamente em anos consecutivos. Dezesseis espécies ocorreram apenas nos 1º e 3º anos, enquanto duas espécies ocorreram apenas nos 1º e 2º anos e três espécies ocorreram apenas nos 2º e 3º anos, padrão semelhante ao observado em três dentre as cinco áreas de coletas (Tabela 3).
Para evitar diferenças devido a deslocamento da área de amostragem em diferentes anos, exatamente a mesma parcela foi amostrada a cada ano com a ajuda de um GPS. Embora as amostragens tenham sido feitas aproximadamente no mesmo período do ano, diferentes condições de temperatura e umidade podem ter deslocado a floração das espécies estudadas nos diferentes anos de amostragem. Com exceção de Assis e Itirapina em 2002, cada área foi visitada apenas uma vez em cada ano, e é possível que várias espécies estivessem presentes nas áreas, mas não em flor. Para definir mais precisamente a distribuição temporal das espécies estudadas, mais amostragens devem ser feitas ao longo do ano.
3.2. Riqueza de Asteraceae nos Cerrados Paulistas
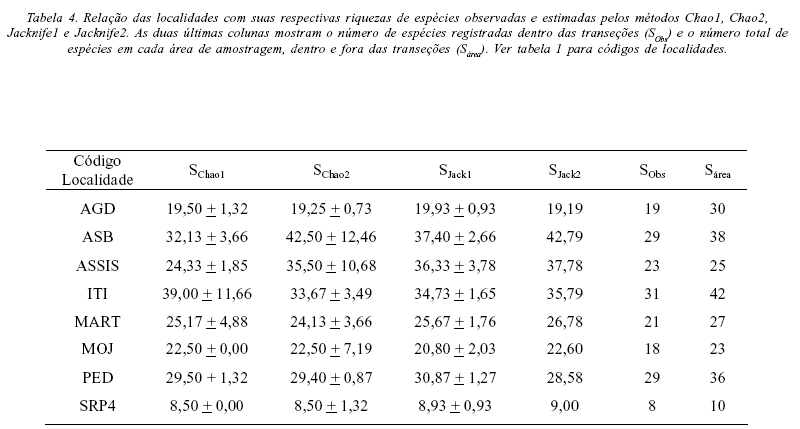
A riqueza de espécies registrada na área (ou seja, incluindo as espécies encontradas por busca ativa fora das transeções) excedeu a riqueza observada nas transeções em todos os remanescentes amostrados (Tabela 4). Ao considerarmos a riqueza total em cada área, Itirapina foi a localidade com maior riqueza (42 espécies) seguida por Águas de Santa Bárbara com 38 espécies, enquanto a gleba Pé de Gigante no Parque Estadual de Vassununga apresentou a menor riqueza, com apenas 10 espécies (Tabela 4). Batalha e Mantovani (2000, 2001) a partir de coletas em toda a área, listaram 42 espécies de Asteraceae para a gleba Pé de Gigante, das quais a metade (21 espécies) estaria florescendo no mês de abril (Batalha & Mantovani 2000). A área limitada de cerrado que amostramos nesta localidade apresentou um forte sombreamento, devido à densidade de árvores e ao grau de fechamento do dossel (Fonseca et al. 2005). A área também não foi amostrada em 2001 (Tabela 3), o que pode ter contribuído para a baixa riqueza observada no presente estudo.
Em todos os remanescentes de cerrados estudados, a ocorrência das espécies nas transeções foi usada para estimar a riqueza nas áreas amostradas. Os valores calculados pelos estimadores foram então comparados ao número de espécies registradas dentro e fora das transeções. Em seis remanescentes os estimadores chegaram a valores de riqueza compatíveis com os registrados na área. Podemos então sugerir que, cumpridas as premissas do estimador, nossa amostra tenha chegado perto da riqueza total, ao menos na parcela onde foram feitas as transeções. Em Martinópolis e em Mogi-Guaçu as transeções estimaram melhor o número de espécies, já que todos os índices tiveram valores compatíveis com o total de espécies que registramos. Nas demais quatro áreas, diferentes estimadores apresentaram valores compatíveis com o número de espécies registradas. Em Itirapina apenas Chao1 foi compatível (com uma alta variância); em Assis os dois índices de Chao foram compatíveis (Chao2 também com alta variância), enquanto em Águas de Santa Bárbara e na gleba Pé de Gigante Chao2 e Jackknife1 foram os melhores estimadores. Os dois estimadores aplicados são baseados no número de espécies raras para estimar a riqueza total de uma área e podem indicar diferentes características das áreas de estudo. O estimador Chao1 será mais alto quanto maior for o número de singletons na amostra (espécies representadas por um único indivíduo, em relação às espécies representadas por dois) (Magurran 2004), e foi o melhor estimador para Itirapina. Os estimadores Chao fornecem estimativas mínimas de riqueza e presumem homogeneidade no elenco de espécies entre amostras (Magurran, 2004). Se esta premissa for verdadeira, Itirapina, Assis, e talvez Martinópolis e Mogi-Guaçu sejam remanescentes mais homogêneos.
Em duas áreas, o número estimado pelos índices ficou abaixo do número de espécies registradas (dentro e fora das transeções): Agudos e Pedregulho (Tabela 4). Nestes casos, a distribuição das espécies nas transeções provavelmente é diferente da encontrada no restante da área amostrada. Este efeito pode ter sido causado pela agregação de indivíduos, o que é comum em asteráceas.
3.3. Comparação entre Floras de Cerrados Brasileiros
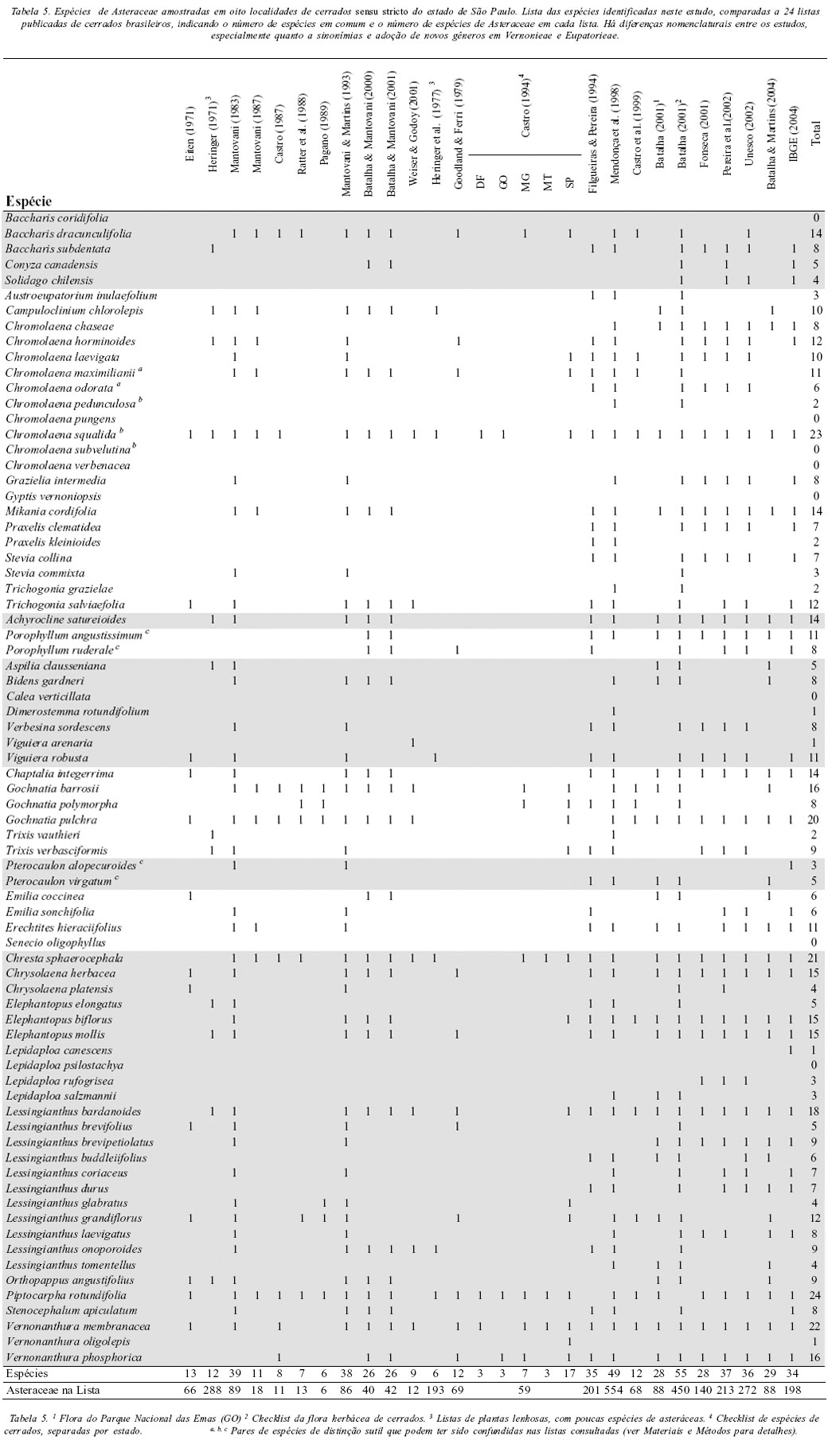
A lista de espécies de Asteraceae identificadas no presente estudo foi comparada a outras 24 listas de estudos realizados em cerrados sensu stricto, das quais 11 referentes apenas a cerrados no estado de São Paulo (Eiten 1971; Heringer 1971; Mantovani 1983, 1987; Castro 1987; Ratter et al. 1988; Pagano et al. 1989; Mantovani & Martins 1993; Batalha & Mantovani 2000, 2001; Weiser & Godoy 2001) e 13 referentes a cerrados em outros estados, podendo conter também dados de São Paulo (Heringer et al. 1977; Goodland & Ferri 1979; Castro 1994; Filgueiras & Pereira 1994; Mendonça et al. 1998; Castro et al. 1999; Batalha 2001 (duas listas: cerrado de Emas e checklist de cerrados brasileiros); Fonseca 2001; Pereira et al. 2002; Unesco 2002; Batalha & Martins 2004; IBGE 2004) (Tabela 5). As áreas mais inventariadas no estado de São Paulo, segundo estas publicações, são a Gleba Pé de Gigante do Parque Estadual de Vassununga, e a Reserva Biológica de Moji-Guaçu,com quatro listas publicadas para cada localidade.
A comparação das listas de espécies mostra claramente que, das espécies que amostramos, Chromolaena squalida é a mais freqüente, tanto dentro quanto fora do estado de São Paulo (21 listas, 87,5%; Tabela 5). Outras espécies amostradas que foram freqüentes nas listas foram Piptocarpha rotundifolia e Gochnatia pulchra (20 listas, 83%), Vernonanthura membranacea e Chresta sphaerocephala (= Eremanthus sphaerocephalus (DC.) Baker) (19 listas, 79%) e Lessingianthus bardanoides (18 listas, 75%) (Tabela 5). Com exceção do gênero Gochnatia, pertencente à tribo Mutiseae, os outros gêneros pertencem à tribo Vernonieae, que, juntamente com a tribo Eupatorieae e Heliantheae, são as mais representativas (em número de espécies) da família nos cerrados paulistas (Figura 4). Estas são também as tribos com maior número de espécies no Brasil (Prado et al. 2002). Lessingianthus bardanoides, V. membranacea, C. sphaerocephala e C. squalida foram mais freqüentemente citadas em estudos fora do estado, enquanto G. pulchra e P. rotundifolia foram citadas em um número semelhante de cerrados dentro e fora do estado.
A freqüência de cada espécie no conjunto de levantamentos publicados está diretamente relacionada com sua freqüência nos remanescentes de cerrados amostrados aqui. Há correlação seja com o conjunto de listas (rS= 0,410; p < 0,001), quanto com as listas somente do estado de São Paulo (rS = 0,386; p < 0,001) ou as de fora do estado (rS= 0,346; p < 0,05).
Os resíduos destas correlações indicam espécies discrepantes em relação à tendência geral. Duas espécies (Chromolaena pedunculosa e C. pungens) foram freqüentes em nossas amostras (7-6 áreas), e ocorreram pouco ou não ocorreram nas listas publicadas. Chromolaena pedunculosa é uma espécie muito semelhante morfologicamente a C. squalida, o que pode ter levado vários autores a identificá-la como esta última (ver métodos para problemas na identificação). Da mesma forma, Lepidaploa salzmannii e Chromolaena chaseae foram freqüentes em nossas amostras e pouco listadas para o estado de São Paulo, enquanto que para as listas de fora do estado de São Paulo, Viguiera arenaria foi freqüente em nosso levantamento e pouco listada.
Ao contrário das espécies acima destacadas, outras espécies freqüentes nas listas publicadas ocorreram pouco em nossas amostras. Chresta sphaerocephala foi freqüente em inventários tanto dentro quanto fora de São Paulo, e somente encontrada em uma área no presente levantamento. Da mesma forma, Gochnatia pulchra e Gochnatia barrosii foram muito citadas para São Paulo, enquanto Elephantopus biflorus e Vernonanthura phosphorica (=Vernonia polyanthes Less.) foram muito listadas fora de São Paulo e pouco observadas no presente levantamento. O presente estudo concentrou amostras em um intervalo de poucos meses no pico de floração da família, assim espécies que florescem exclusivamente em outros períodos podem não ter sido observadas. Esse fato pode ter sido determinante para as espécies C. sphaerocephala, G. pulchra e G. barrosii, já que apresentam floração principalmente no período de junho a início de setembro (ver Mantovani e Martins, 1993). Em nosso estudo optamos por utilizar o mesmo protocolo e intensidade de amostragem, e num curto intervalo, para oito das mais importantes localidades de cerrado do estado, o que permite a avaliação e comparação de sua riqueza de espécies.
Dentre as 74 espécies identificadas de nossas amostras de cerrados, oito (11%) não estão listadas em nenhum dos estudos consultados: Chromolaena pungens, C. subvelutina, C. verbenacea, Calea verticillata, Baccharis coridifolia, Gyptis vernoniopsis, Senecio oligophyllus e Lepidaploa psilostachya (Tabela 5). Com exceção de Chromolaena subvelutina (uma nova combinação ainda não publicada, ver métodos) as outras espécies não foram descritas nem revalidadas recentemente, o que poderia justificar sua ausência nas listas publicadas.
Asteraceae é um grupo de taxonomia difícil e com relativamente poucos especialistas no Brasil para o tamanho da família. Algumas espécies podem facilmente ser confundidas com outras mais comuns, e assim passarem desapercebidas em um estudo de florística geral de cerrados, em que o esforço de identificação abrange muitas famílias. O fato de termos registrado espécies que não se encontram listadas em nenhum dos estudos publicados reflete em parte este problema. A espécie Chromolaena pungens ilustra a possibilidade de erro de identificação de uma espécie comum; sua alta freqüência (seis áreas e 13 amostras) no presente estudo, nos leva a supor que tenha sido anteriormente coletada, porém confundida com outra espécie.
Esta não é, entretanto, a única razão para os novos registros de espécies, mesmo em áreas bastante conhecidas e apesar do número e qualidade de levantamentos florísticos precedentes em cerrados de várias regiões. Como o presente levantamento concentrou-se nas asteráceas, é possível que tenhamos coletado espécies que teriam passado desapercebidas em parte dos estudos florísticos gerais, especialmente as mais raras e inconspícuas.
O presente estudo mostrou uma grande proporção de unicatas (espécies com uma única amostra) nos cerrados do estado de São Paulo. Assim é possível que existam espécies ainda não detectadas em nenhum estudo de flora feito até o momento. Além disso, a pouca repetição de espécies entre localidades sugere que os remanescentes de cerrados paulistas são áreas isoladas, frágeis e com uma alta proporção de espécies exclusivas. O tamanho reduzido de muitos destes remanescentes sugere também que muitas das espécies raras tenham populações muito pequenas, de baixa viabilidade em longo prazo, e com reduzida oportunidade de intercâmbio genético ou recolonização a partir de outras áreas.
Enfatizamos que, apesar do maior interesse histórico na flora árborea/lenhosa dos cerrados, o estudo da flora herbácea e sub-arbustiva é igualmente importante. A manutenção e preservação destes componentes merecem interesse e preocupação, embora sejam menos conhecidos e inventariados que as árvores deste bioma.
4. Agradecimentos
O presente estudo foi financiado pela Fundação de Amparo à Pesquisa do Estado de São Paulo no projeto temático "Diversidade de espécies e de interações em plantas e insetos fitófagos" (98/05085-2), integrante do Programa Biota/Fapesp. O auxílio de vários especialistas foi imprescindível: Mara Magenta e Claudio Augusto Mondin identificaram as espécies da tribo Heliantheae, Roberto L. Esteves identificou as espécies da tribo Eupatorieae e João Semir ajudou na identificação das espécies da tribo Vernonieae. Harold Robinson, John Pruski e Nelson Ivo Matzenbacher identificaram espécimes de estudos anteriores e esclareceram dúvidas. Marisa D. Bitencourt, Humberto N. Mesquita Jr., Geraldo Kuntschik, Giselda Durigan e Marinez F. de Siqueira compartilharam imagens de satélite e listas de espécies arbóreas de áreas de cerrado, discutindo e ajudando na escolha das áreas de estudo. A.M. Almeida e C.R. Fonseca receberam bolsa de pós-doutorado FAPESP. U. Kubota recebeu bolsa de mestrado FAPESP. M.R. Braun, T.G. Mendonça, S.M. Futada, R.L.G. Raimundo, L.A. Anjos e M. Almeida-Neto receberam bolsas de treinamento técnico FAPESP. Agradecemos a toda equipe do projeto, que participou do trabalho de campo, triagem do material e das discussões no laboratório: Ricardo Fabiano, E.P. Anseloni, L.B. Mendes, A.E. Fraez, , M. Portella, A.M. Rosa; às administrações e funcionários do Instituto Florestal de São Paulo, pelo acesso às áreas de estudo e às instalações e à AMBEV, pelo acesso à área de cerrado em Agudos. Dois revisores anônimos fizeram sugestões importantes que foram incorporadas ao manuscrito.
5. Referências Bibliográficas
Recebido em 06/10/20004
Revisada em 04/03/2005
Publicado em 01/07/2005
ISSN 1676-0603
- ALHO, C.J.R. 1994. Distribuição da fauna num gradiente de recursos em mosaico. In Cerrado: caracterização, ocupação e perspectivas. 2a. Edição (M.N. PINTO, org.), Ed. Universidade de Brasília. p. 213-264.
- ALHO, C.J.R. & MARTINS, E.S. 1995. De Grão em Grão, o Cerrado Perde Espaço (Cerrado - Impactos do Processo de Ocupação). Brasília, DF, Brasil: WWF.
- ALMEIDA, A.M., PRADO, P.I. & LEWINSOHN, T.M. 2004. Geographical distribution of Eupatorieae (Asteraceae) in South-eastern and South Brazilian mountain ranges. Plant Ecology. 174: 163-181.
- BATALHA, M. A. 2001. Florística, espectro biológico e padrões fenológicos do cerrado senso lato no Parque Nacional das Emas (GO) e o componente herbáceo- subarbustivo da flora do cerrado senso lato Tese de Doutorado, Universidade Estadual de Campinas, Campinas, SP.
- BATALHA, M.A. & MANTOVANI, W. 2000. Reproductive phenological patterns of cerrado plant species at the Pé-de-Gigante reserve (Santa Rita do Passa Quatro, SP, Brazil): a comparison between the herbaceous and woody floras. Rev. Bras. Biol. 60: 129-145.
- BATALHA, M.A. & MANTOVANI, W. 2001. Floristic composition of the Cerrado in the Pé-de-Gigante reserve (Santa Rita do Passa Quatro, Southeastern Brazil). Acta bot. Bras. 15: 289-302.
- BATALHA, M.A., MANTOVANI, W. & MESQUITA JÚNIOR, H.N. 2001. Vegetation structure in cerrado physiognomies in south-eastern Brazil. Brazil. J. Biol. 61: 475-483.
- BATALHA, M.A. & MARTINS, F.R. 2004. Reproductive phenology of the cerrado plant community in Emas National Park (central Brazil). Aust. J. Bot. 52: 149-161.
- BARROSO, G. M. 1986. Sistemática de Angiospermas do Brasil. Imprensa da Universidade Federal de Viçosa, Viçosa.
- BREMER, K. 1994. Asteraceae: cladistics and classification. Timber Press, Portland.
- BRIDGEWATER, S., RATTER, J.A. & RIBEIRO, J.F. 2004. Biogeographic patterns, b-diversity and dominance in the cerrado biome of Brazil. Biodiv. Cons. 13: 2295-2318.
- BRITO, M.C.W. (coord.) 1997. Workshop: Bases para Conservação e Uso Sustentável das Áreas de Cerrado do Estado de São Paulo. Secretaria de Meio Ambiente do Estado de São Paulo. PROBIO/SP.
- CASTRO, A.A.J.F. 1987. Florística e fitossociologia de um cerrado marginal brasileiro, Parque Estadual de Vassununga, Santa Rita do Passa Quatro, São Paulo. Tese de Mestrado. Universidade Estadual de Campinas, Campinas, SP.
- CASTRO, A.A.J.F. 1994. Comparação florística geográfica (Brasil) e fitossociológica (Piauí São Paulo) de amostras de cerrado. Tese de Doutorado. Universidade Estadual de Campinas, Campinas, SP.
- CASTRO, A.A.J.F.; MARTINS, F.R.; TAMASHIRO, J.Y. & SHEPHERD, G. 1999. How rich is the flora of brazilian cerrados? Ann. Missouri Bot. Garden. 86:192- 224.
- COLWELL, R. K. 2004. EstimateS: Statistical estimation of species richness and shared species from samples. Version 7. User's Guide and application published at: http://purl.oclc.org/estimates
- COLWELL, R.K. & CODDINGTON, J.A. 1994. Estimating terrestrial biodiversity through extrapolation. Phil. Trans. R. Soc. Lond. B 345:101-118.
- DURIGAN, G., SIQUEIRA, M.F., FRANCO, G.A.D.C., BRIDGEWATER, S. & RATTER, J.A. 2003. The vegetation of priority areas for cerrado conservation in São Paulo State, Brazil. Edinb. J. Bot. 60: 217-241.
- EITEN, G. 1971. Habitat of Fazenda Campininha. São Paulo, Brazil. In Simpósio sobre o Cerrado. (M.G. FERRI, coord.). Edusp. Ed. Edgard Blücher LTDA. p.155-202.
- EITEN, G. 1994. Vegetação. In: Cerrado: caracterização, ocupação e perspectivas. 2a. Edição (M.N. PINTO, org.). Ed. Universidade de Brasília. p. 17-74
- ESTEVES, R.L. 2001. O gênero Eupatorium s.l. (Compositae Eupatorieae) no estado de São Paulo Brasil. Tese de Doutorado. Universidade Estadual de Campinas, Campinas, SP.
- FILGUEIRAS, T.S. & PEREIRA, B.A.S. 1994. Cerrado: caracterização, ocupação e perspectivas. (M.N. PINTO, org.). 2º ed. Editora da Universidade de Brasília. p. 345-404.
- FONSECA, C.R; PRADO, P.I; ALMEIDA-NETO, M; KUBOTA, U. & LEWINSOHN, T.M. 2005. Flower-heads and their insects: food web structure along a fertility gradient of Cerrado. Ecol. Ent. 30: 36-46.
- FONSECA, F.O. (org.) 2001. Olhares sobre o lago Paranoá .Secretaria de Meio Ambiente e Recursos Hídricos Semarh: //http:www.semarh.df.gov.br/site/
- GOODLAND, R. & FERRI, M.G. 1979. Ecologia do Cerrado. Editora da Universidade de São Paulo e Livraria Itatiaia Ed.
- HERINGER, E.P. 1971. Propagação e sucessão de espécies arbóreas do cerrado em função do fogo, do cupim, da capina e do aldrim (inseticida). In III Simpósio sobre cerrado. (M. G. FERRI, coord.), Editora da Universidade de São Paulo e Editora Edgard Blüncher LTDA. p.167-169.
- HERINGER, E.P., BARROSO, G.M., RIZZO, J.A. & RIZZINI, C.T. 1977. A flora do Cerrado. In: IV Simpósio sobre cerrado: bases para utilização agropecuária. (M.G. FERRI, coord.) Editora da Universidade de São Paulo e Editora Edgard Blüncher LTDA. p. 211-232.
- IBGE. 2004. Reserva Ecológica do IBGE: Ambiente e Plantas Vasculares. Rio de Janeiro: Estudos e Pesquisas. Informação Geográfica no. 3. Rio de Janeiro. IBGE-Divisão de Geociências.
- KING, R.M. & ROBINSON, H. 1987. The genera of the Eupatorieae (Asteraceae). Monographs in Systematic Botany. Missouri Botanical Garden, St. Louis, MO, 22.
- KRONKA, F.J.N., NALON, M.A., MATSUKUMA, C.K., PAVÃO, M., GUILLAUMON, J.R., CAVALLI, A.C., GIANNOTTI, E. IWANE, M.S.S., LIMA, L.M.P.R., MONTES, J. DEL CALI, I.H. & HAACK, P.G. 1998. Áreas de domínio do cerrado no estado de São Paulo. São Paulo: Secretaria de Estado do Meio Ambiente, Instituto Florestal.
- MAGURRAN, A.E. 2004. Measuring biological diversity. Blackwell Publishing, Oxford UK.
- MANTOVANI, W. 1983. Composição e similaridade florística, fenologia e espectro biológico do cerrado da reserva biológica de Moji Guaçu, Estado de São Paulo. Tese de Mestrado. Universidade Estadual de Campinas, Campinas, SP.
- MANTOVANI, W. 1987. Análise florística e fitossociológica do estado herbáceo- subarbustivo do cerrado na reserva biológica de Moji Guaçu e em Itirapina. Tese de Doutorado. Universidade Estadual de Campinas, Campinas, SP.
- MANTOVANI, W. & MARTINS, F.R. 1993. Florística do cerrado na Reserva Biológica de Moji Guaçu, SP. Acta bot. Bras. 7: 33-60.
- MENDONÇA, R.C., FELFILI, J.M., WALTER, B.M.T., JÚNIOR, M.C.S., REZENDE, A.V., FILGUEIRAS, T.S. & NOGUEIRA, P.E. 1998. Flora vascular do cerrado. In Cerrado: ambiente e flora (S.M. SANO & S.P. ALMEIDA, eds.). EMBRAPA-CPAC. p. 289-556.
- MMA/SBF. 2002. Biodiversidade Brasileira: Avaliação de áreas e ações prioritárias para a conservação, utilização sustentável e repartição dos benefícios da biodiversidade nos biomas brasileiros. Brasília.
- PAGANO, S.N.; CESAR, O.; LEITÃO-FILHO, H.F. 1989. Composição florística do estrato arbustivo- arbóreo da vegetação de cerrado da área de proteção ambiental (APA) de Corumbataí estado de São Paulo. Rev. Brasil. Biol., 49(1): 49-59.
- PEREIRA, B.A.S., SILVA, M.A., MENDONÇA, R.C. 2002. Reserva ecológica do IBGE, Brasília-DF: lista das plantas vasculares. Rio de Janeiro: IBGE-Divisão de Geociências do Distrito Federal.
- PINTO, M.N. 1994. Cerrado: caracterização, ocupação e perspectivas. 2a. Edição. Ed. Universidade de Brasília.
- PRADO, P.I. & LEWINSOHN, T.M. 2000. Associações inseto-planta no nível local e regional: Tephritidae e Vernonieae na Serra do Espinhaço. In Ecologia e comportamento de insetos(Série Oecologia Brasiliensis vol. VIII) ( R.P. Martins, T.M. Lewinsohn & M.S. Barbeitos, orgs.). Rio de Janeiro, 2000, p. 405-422.
- PRADO, P.I., LEWINSOHN, T.M., ALMEIDA, A.M., NORRBOM, A.L., BUYS, B.D., MACEDO, A.C. & LOPES, M.B. 2002. The fauna of Tephritidae (Diptera) from capitula of Asteraceae in Brazil. Proc. Entomol. Soc. Wash. 104: 1007-1028.
- RATTER, J.A., BRIDGEWATER, S., ATKINSON, R. & RIBEIRO, J.F. 1996. Analysis of the floristic composition of the Brazilian cerrado vegetation II: comparison of the woody vegetation of 98 areas. Edinb. J. Bot. 53: 153-180.
- RATTER, J.A., BRIDGEWATER, S. & RIBEIRO, J.F. 2003. Analysis of the floristic composition of the Brazilian cerrado vegetation III: Comparison of the woody vegetation of 376 areas. Edinb. J. Bot. 60: 57-109.
- RATTER, J.A. & DARGIE, T.C.D. 1992. An analysis of the floristic composition of 26 cerrado areas in Brazil. Edinb. J. Bot. 49: 235-250.
- RATTER, J.A., LEITÃO-FILHO, H.F, ARGENT, G., GIBBS, P.E., SEMIR, J., SHEPHERD, G. & TAMASHIRO, J.Y. 1988. Floristic and estructure of a brazilian cerrado. Notes R. B. G. Edinburgh. 45(1):137- 151.
- RATTER, J.A., RIBEIRO, J.F. & BRIDGWATER, S. 1997. The Brazilian Cerrado vegetation and threats to its biodiversity. Ann. Bot. 80: 223-230.
- RIBEIRO, J.F. & WALTER, B.M.T. 1998. Fitofisionomias do bioma cerrado. In Cerrado: ambiente e flora (S.M. SANO, & S.P. ALMEIDA, eds.). EMBRAPA-CPAC. p. 89-168.
- SILVA, J.M.C. & BATES, J.M. 2002. Biogeographic patterns and conservation in the south american cerrado: a tropical savanna hotspot. Bioscience 52: 225-233.
- UNESCO. 2002. Vegetação do Distrito Federal: tempo e espaço. 2º. Edição. Brasília, DF.
- WEISER, V.L. & GODOY, S.A.P. 2001. Florística em um hectare de cerrado stricto sensu na arie-cerrado pé-de-gigante, Santa Rita do Passa Quatro, SP. Acta bot. Bras. 15 (2) :201-212.
Datas de Publicação
-
Publicação nesta coleção
23 Out 2006 -
Data do Fascículo
2005
Histórico
-
Aceito
01 Jul 2005 -
Revisado
04 Mar 2005 -
Recebido
06 Out 2004