Abstracts
In the present study, four species of Nephtyidae, Aglaophamus juvenalis (Kinberg 1866), Nephtys acrochaeta Hartman 1950, Nephtys californiensis Hartman 1938 and Nephtys squamosa Ehlers 1887, were found from the intertidal zone to the shallow sublittoral (<50 m) off São Paulo, Brazil, during the program BIOTA/FAPESP Marine Benthos. Descriptions and notes on each of them are provided. Nephtys californiensis is a new record for the Brazilian coast. Keys to genera and species of Nephtyidae recorded from Brazil are given.
Polychaeta; Nephtyidae; Aglaophamus; Nephtys; key to identification; new record
Neste estudo, quatro espécies de Nephtyidae: Aglaophamus juvenalis (Kinberg 1866), Nephtys acrochaeta Hartman 1950, Nephtys californiensis Hartman 1938 e Nephtys squamosa Ehlers 1887 foram coletadas desde a região entremarés até o sublitoral raso (<50 m) da costa de São Paulo, Brasil, durante o Programa BIOTA/FAPESP Bentos Marinho. Descrições e comentários são fornecidos para cada uma delas. Nephtys californiensis é um novo registro para a costa brasileira. Chaves para gêneros e espécies de Nephtyidae registrados no Brasil são fornecidas.
Polychaeta; Nephtyidae; Aglaophamus; Nephtys; chave de identificação; novo registro
ARTICLES
Nephtyidae (Annelida: Polychaeta) from São Paulo State, Brazil, including a new record for the Brazilian coast
Nephtyidae (Annelida: Polychaeta) do Estado de São Paulo, Brasil, incluindo um novo registro para a costa brasileira
Alexandra Elaine Rizzo1 1 Corresponding author: Alexandra Elaine Rizzo, e-mail: aerizzo@hotmail.com, http://www.ib.unicamp.br/ ; Antonia Cecília Zacagnini Amaral
Departamento de Zoologia, Instituto de Biologia, Universidade Estadual de Campinas, Unicamp, CP 6109, CEP 13083-970, Campinas, SP, Brazil
ABSTRACT
In the present study, four species of Nephtyidae, Aglaophamus juvenalis (Kinberg 1866), Nephtys acrochaeta Hartman 1950, Nephtys californiensis Hartman 1938 and Nephtys squamosa Ehlers 1887, were found from the intertidal zone to the shallow sublittoral (<50 m) off São Paulo, Brazil, during the program BIOTA/FAPESP Marine Benthos. Descriptions and notes on each of them are provided. Nephtys californiensis is a new record for the Brazilian coast. Keys to genera and species of Nephtyidae recorded from Brazil are given.
Keywords: Polychaeta, Nephtyidae, Aglaophamus, Nephtys, key to identification, new record.
RESUMO
Neste estudo, quatro espécies de Nephtyidae: Aglaophamus juvenalis (Kinberg 1866), Nephtys acrochaeta Hartman 1950, Nephtys californiensis Hartman 1938 e Nephtys squamosa Ehlers 1887 foram coletadas desde a região entremarés até o sublitoral raso (<50 m) da costa de São Paulo, Brasil, durante o Programa BIOTA/FAPESP Bentos Marinho. Descrições e comentários são fornecidos para cada uma delas. Nephtys californiensis é um novo registro para a costa brasileira. Chaves para gêneros e espécies de Nephtyidae registrados no Brasil são fornecidas.
Palavras-chave: Polychaeta, Nephtyidae, Aglaophamus, Nephtys, chave de identificação, novo registro.
Introduction
Currently, five genera of Nephtyidae are known (Aglaophamus, Dentinephtys, Inermonephtys, Micronephtys, and Nephtys) with 186 nominal species, about 30% of these synonyms or nomina nuda (Rizzo & Lana, in prep.). They are distinguishable from other polychaetes by the clear presence of interramal branchiae on the ventral notopodial edge (Fauchald & Rouse 1997, Wilson 2000). Interramal branchiae have also been called "recurved cirrus" (Hartman 1938, 1940) or "interramal cirrus" (Hartman 1950, Paxton 1974, Fauchald 1976, Rainer & Hutchings 1977, Garwood & Olive 1981, Imajima & Takeda 1985, 1987, Rainer 1989, Hilbig 1997, Lovell 1997); they are located on the lower part of the notopodia, except on the anteriormost parapodia. When present (the majority of the Micronephtys species have no interramal branchiae), they are coiled toward the interior (inward) or exterior (outward). The rectangular to rounded prostomium usually ends in a dorsal pair of antennae and a ventral pair of palps, both non-articulated; the dorsal pair is absent in Inermonephtys. Subdermal eyes are usually absent; if present, they migrate from the prostomium to the anterior segments during ontogeny. A pair of ciliated nuchal organs is located dorsolaterally on the posterior margin of the prostomium. Tentacular cirri are absent. The peristomium is not visible dorsally; ventrally it is limited to the lips. The pharynx is eversible, and contains rows of soft subdistal papillae, in addition to a distal ring of papillae (absent in Inermonephtys). A pair of triangular to fusiform lateral jaws (paragnaths) is present; a pair of denticulate plates with 8-9 teeth is present in Dentinephtys (Imajima & Takeda 1987). The segments are similar in shape (the first segment with chaetae is smaller than the following ones), and all parapodia are biramous. Parapodial lobes are triangular to round, and have prechaetal and postchaetal lamellae besides dorsal and ventral cirri; each is borne by the aciculum. A dorsal cirrus is present on the interramal branchia; it is smaller than the ventral cirri, which are on the base of the neuropodium. Noto- and neurochaetae are similar to each other, and may be smooth or serrate, capillary, simple and sometimes chambered (or barred). Furcate chaetae are present in Aglaophamus, Inermonephtys and Micronephthys. Long natatory chaetae may be present in some taxa during the reproductive period. The pygidium has a dorsal anus with only one pygidial cirrus, which is characteristic among the Phyllodocida (Pleijel & Dahlgren 1998).
Cladistical relationships within the group are uncertain, and there is no evidence of apomorphies for Nephtys, Aglaophamus or Micronephtys (Rouse & Pleijel 2001). Thus, among the main diagnostic characteristics of a nephtyid are: shape and origin of the interramal branchia, morphology of the acicular lobes and parapodial lamellae, arrangement and number of the pharyngeal papillae, and type and arrangement of the chaetae. This last has usually been overlooked.
References to nephtyids previously reported from the Brazilian coast are found in Lana (1986), who mentioned the occurrence of 13 species: Nephtys laciniosa Grube 1881, Nephtys acrochaeta Hartman 1950, Nephtys caeca (Fabricius 1780), Nephtys fluviatilis Monro 1937, Nephtys hombergi Savigny 1818, Nephtys magellanica Augener 1912, Nephtys simoni Perkins 1980, Nephtys squamosa Ehlers 1887, Aglaophamus macroura (Schmarda 1961), Aglaophamus uruguayi Hartman 1953, Aglaophamus juvenalis (Kinberg 1866), Aglaophamus dibranchis (Grube 1877), and Inermonephtys palpata Paxton 1974. This last record has been revised and is now considered an erroneous identification; as a consequence, a new species of Inermonephtys from Brazil is being published (Martin, Gil & Lana, in press). In addition to these, Nephtys paradoxa Malmgren 1874 has been recorded from the sublittoral at Ubatuba on the northern coast of São Paulo (Morgado 1988). In addition to I. palpata, some of these species may have been misidentified. Nephtys caeca and N. paradoxa, for example, are cold-water species that have been recorded in the Pacific and Arctic oceans (Hartman 1950, Paxton 1974, Garwood & Olive 1981, Imajima & Takeda 1987). Nephtys laciniosa is known only from the original description, and was considered "indeterminable" by Hartman (1950: 91).
In this study, four species of Nephtyidae: A. juvenalis, N. acrochaeta, Nephtys californiensis Hartman, 1938 and N. squamosa were found from the intertidal zone to the shallow sublittoral (<50 m) off São Paulo state, Brazil, during the program BIOTA/FAPESP Marine Benthos. Nephtys californiensis is recorded for the first time from the Brazilian coast. Keys to genera and species recorded in Brazil are given.
Material and Methods
Specimens of Nephtyidae were collected during the program "BIOTA/FAPESP Bentos Marinho" (Benthic Marine Biodiversity in the State of São Paulo), sampling from intertidal zone to non-consolidated sublittoral bottoms (<50 m depth), from June 2001 to December 2002. The Northern coast of São Paulo State (23-24° S and 44-46° W), includes the municipalities of Ubatuba, Caraguatatuba, São Sebastião and Ilhabela. The polychaetes were sorted out from the sediment, fixed with 4% formalin, stored in 70% ethanol, and identified. The specific methods can be found in Amaral et al. (2006). Measurements and line drawings were made using ZEISS optical microscopy and stereomicroscopy. The examined material was deposited in the Museu de Historia Natural of the Universidade Estadual de Campinas (MHN-UNICAMP), Polychaeta Collection (under the abbreviation ZUEC-BPO AR).
Results and Discussion
1. Systematics
Nephtyidae Grube 1850
Type genus:Nephtys Cuvier 1817
Key to Genera of Nephtyidae Recorded in Brazil:
1. Interramal branchiae recurved ( coils outward)......................................Nephtys
- Interramal branchiae involute (coils inward).....................................................2
2. Dorsal pair of antennae present; pharynx with terminal papillose ring............................................................................................Aglaophamus
- Dorsal pair of antennae absent; pharynx without terminal papillose ring..........................................................................................Inermonephtys
Key to Species of Aglaophamus Recorded in Brazil:
1. Interramal branchiae starting from parapodium 4 or 4-5; proboscis with fewer than 20 rows of subdistal papillae...........................................................................2
- Interramal branchiae starting from parapodium 3 or 3-4; proboscis with 20 or more rows of subdistal papillae................................................................................3
2. Interramal branchiae starting from parapodium 4; proboscis with 20 paired distal papillae,16 subdistal papillae in rows of 4-6 and a long unpaired middorsal papilla ...................................................................................................A. juvenalis
- Interramal branchiae starting from parapodium 4-5; proboscis with 22 paired distal papillae, 14 subdistal papillae in rows of 7-8 and a ong unpaired middorsal papilla .................................................................................................A. dibranchis
3. Interramal branchiae starting from parapodium 4; proboscis with 20 paired distal papillae, 20 subdistal papillae in rows of 6-9 and an unpaired middorsal papilla...........................................................................................A. uruguayi
- Interramal branchiae starting from parapodium 3-4; proboscis with 20 paired distal papillae, 22 subdistal papillae in rows of 2-3 and a short unpaired middorsal and midventral papilla...........................................................................A. macroura
1. Aglaophamus Kinberg 1866
Type species:Aglaophamus lyratus Kinberg 1866
Description: Prostomium with a dorsal pair of antennae and a ventral pair of palps. Proboscis eversible, with longitudinal rows of subdistal papillae with a variable number of papillae in each row; distally with paired distal papillae; an unpaired middorsal papilla may be present. Internal pair of triangular paragnaths. Tentacular segment with enlarged neuropodial lobes. Notopodial and neuropodial parapodia with preacicular and postacicular lobes; dorsal and ventral cirrus. Interramal branchiae involute (curls inward). Barred, capillary and furcate chaetae may be present.
2. Aglaophamus juvenalis (Kinberg 1866)
(Figure 1)
Aglaopheme juvenalis Kinberg 1866:240
Aglaophamus juvenalis Hartman 1948:51; Fauchald 1976:16-18, Figures 1a-c; Lana 1986: 143-144, Figures 16-17.
Material examined: 1 specimen: ZUEC-BPO - AR799 (23° 50' 70.5'' S and 45° 31' 95.2'' W, Sta. 9i, 15.ii.2001, 14.7 m).
Description: Based on incomplete specimen, measuring 25 mm long and 2 mm wide (including parapodia, excluding chaetae), with 61 chaetigers. Color yellowish, with a longitudinal dorsal orange spot, more evident on anterior region of body. Body robust and cylindrical, becoming slender and squared in posterior region. Prostomium trapezoidal, small, translucent, wider than long, with a small middorsal slit, not prominent (Figure 1a). A dorsal pair of antennae and a ventral pair of palps; both inserted latero-frontally, conical, as long as half of prostomium. A pair of nuchal organs latero-dorsally on posterior margin of prostomium (Figure 1a). Proboscis partially everted, with 20 paired distal papillae (outer papillae larger than inner), lateral papillae larger than dorsal and ventral papillae. Sixteen rows of subdistal papillae arranged longitudinally along proboscis; each row with about six conical papillae; ventral papillae rows smaller than lateral and dorsal; basal papillae becoming larger towards distal end; and one, unpaired papilla, three times larger than others. Basal region of proboscis smooth. Parapodia biramous laterally on the body; except first parapodium positioned frontally. Notopodia and neuropodia (Figure 1b) less developed on anterior parapodia. Pre-acicular and postacicular lobes rounded; postacicular lobe larger than pre-acicular lobe. Dorsal cirrus conical and short on anterior parapodia (Figure 1c), elongated and filiform on median parapodia, and rudimentary on posterior parapodia (Figure 1d); dorsal cirrus not seen on first parapodium. Interramal branchiae involute from fourth parapodium, laterally ciliated (Figure 1c-d); on first parapodia occupying one-tenth of interramal space; on middle parapodia (when the larger size begins) occupying one-third of interramal space, reducing from parapodia 44 and disappearing on posterior parapodia (Figure 1e). Conical neuropodial prechaetal lobe and more-developed, rounded postacicular lobe, mainly on median parapodia (Figure 1c-d). Posterior parapodia with notopodial and neuropodial pre-acicular lobes rounded; postacicular lobes conical and distally slender, without developed expansion. An inflated lamella on neuropodial lobe of parapodia 10-25 (Figure 1c). Ventral cirrus as long as neuropodial lobe, becoming conical on anterior parapodia (Figure 1b); short and slender on posterior parapodia (Figure 1e). Dark chaetae, mainly on posterior parapodia. Barred pre-acicular chaetae (Figure 1f-g) distally slender and coiled. Furcate chaetae, present on postacicular notopodium and neuropodium, accompanying capillary chaetae with slender, marginally serrulate shaft, fewer in number than other types (Figure 1h). Post-acicular capillary chaetae apparently smooth, marginally rough, without visible denticles (Figure 1i); slender and elongated on posterior parapodia. Number of barred chaetae larger than capillary chaetae on anterior parapodia, reducing gradually towards posterior, where capillary chaetae increase. Specimen with oocytes, each measuring about 50 µm in diameter.
Remarks: Hartman (1948) mentioned longitudinal rows of subdistal proboscideal papillae with about 4 papillae, Fauchald (1976) with 5-6 papillae, and Lana (1986) with 4-5 papillae, whereas the specimens examined here have about 6 papillae per row. A dorsal cirrus was not observed on the first parapodium, although Fauchald (1976) has reported its presence. We observed a pouch-like lamella, sometimes inflated, on the median parapodia. This same structure was mentioned as a digitate lobe by Hartman (1948) and by Lana (1986). Aglaophamus juvenalis is close to A. uruguayi. However, it can be distinguished from A. uruguayi by the number of the proboscideal papillae (only from the subdistal, because both have 20 distal papillae; see key to species above), by the size of the branchiae and the development of the median parapodia. Aglaophamus uruguayi was described from material that was presumably Uruguayan (Hartman 1950), but the geographical coordinates refer to Rio Grande do Sul state, Brazil (Lana 1986). Only a few specimens of A. juvenalis have been recorded in southeastern Brazil, usually in shallow bottoms. Specimens previously identified as A. dibranchis by Temperini (1981) were referred to A. juvenalis by Lana (1986).
Occurrence: This species is recorded in southeastern Brazil (from the states of Rio de Janeiro to Paraná). It was originally described from Rio de Janeiro, collected in the shallow sublittoral (2-3 m depth), during the Eugenies Expedition to South America. In São Paulo, it was recorded in the intertidal zone, in fine sand of Dura Beach, in Ubatuba (Fauchald 1976). In Paraná, it was found in Paranaguá Bay and Laranjeiras and on the continental slope, from 6-71 m depth, in bottoms with sand and mud, sand with silt and mud, and silt and mud with shell fragments (Lana 1986). In this study, A. juvenalis was collected at Caraguatatuba, at 14.7 m depth, in bottoms with very fine sand.
Key to Species of Nephtys Recorded on Brazil:
1. Notopodia with well-developed expansions and imbricated arrangement, mainly on mid-body chaetigers.........................................................................................2
- Notopodia without well-developed expansions and imbricated arrangement...............8
2. Interramal branchiae starting before parapodium 6..............................................3
- Interramal branchiae starting from parapodium 9 or 10-11; spur-like chaetae present.........................................................................................N. acrochaeta
3. Prostomial central region with characteristic pigmentation pattern (present even in specimens conserved in alcohol for a long time).....................................................4
- Prostomial central region without characteristic pigmentation pattern......................5
4.Interramal branchiae starting from parapodium 3; proboscis with long middorsal and midventral subdistal papilla; proboscis with 23 conical distal papillae...............N. simoni.
- Interramal branchiae starting from parapodium 3-4; proboscis without long middorsal or midventral subdistal papilla; proboscis with 21 conical distal papillae.......................................................................................N. californiensis
5. Proboscis with long middorsal subdistal papilla....................................................6
- Proboscis without long middorsal subdistal papilla; interramal branchiae starting from parapodium 4-6, usually 4........................................................................N. caeca
6. Proboscis without long midventral subdistal papilla; spur-like chaetae absent........................................................................................................7
- Proboscis with long midventral subdistal papilla; spur-like chaetae present; interramal branchiae starting from parapodium 1-4, usually 3...................................N. squamosa
7. Interramal branchiae starting from parapodium 4-6, usually 5 (nearly disappearing at the end of the body); acicular lobes with prominent medial bulb or papilla..............................................................................................N. hombergi
- Interramal branchiae starting from parapodium 5 (disappearing after 36th); acicular lobes without prominent medial bulb or papilla; dorsally yellowish to brownish...........................................................................................N. fluviatilis
8. Interramal branchiae starting from parapodium 3; proboscis with long middorsal subdistal papilla..............................................................................N. magellanica
- Interramal branchiae starting from parapodium 8-12; proboscis without long middorsal subdistal papilla.................................................................................N. paradoxa
3. Nephtys Cuvier 1817
Type species:Nephtys hombergi Savigny 1818
Description: Prostomium quadrangular, pentagonal or rounded, with a dorsal pair of antennae and a ventral pair of palps, both inserted anterolaterally. A pair of nuchal organs on posterior edge of the prostomium. Proboscis eversible, cylindrical, with three distinct regions: proximal region smooth or covered with small wart-like processes; subdistal region with rows of papillae, and a long unpaired middorsal, and sometimes also a midventral, papilla may be present; distal region with arc of paired papillae. Paired distal papillae and rows of subdistal papillae numbers are invariant for each species. Parapodia with lobes widely separated; conical acicular lobes rounded or incised; postacicular and preacicular lobes short and rounded to well developed and foliaceous. Interramal branchiae recurved (curls outward) or straight. Dorsal and ventral cirrus present, conical to digitiform. Preacicular chaetae short, barred; postacicular chaetae elongated, smooth, spinulose or serrulate; furcated chaetae absent. One aciculum per parapodial ramus, distally slender and slightly curved, or straight and blunt. Pygidium small, narrow, with long, filiform, ventral anal cirrus.
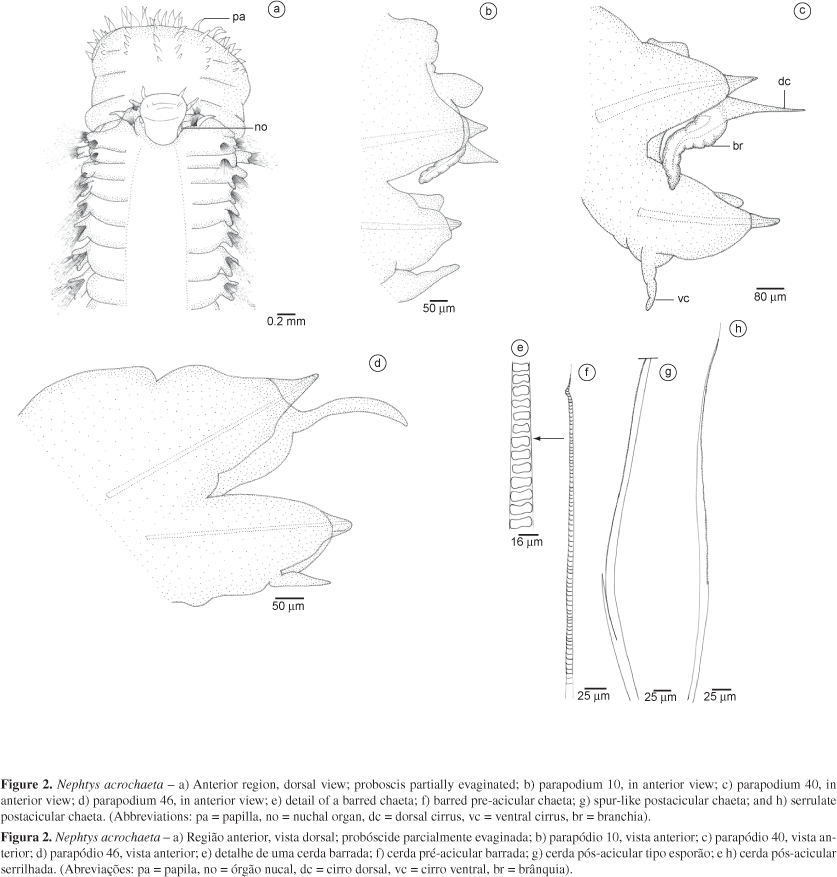
4. Nephtys acrochaeta Hartman 1950
(Figure 2)
Nephtys acrochaeta Hartman 1950: 114-116, pl. 16, Figures 1-6; Fauchald 1976: 18, Figures 1d-e; Lana 1986: 138-139, Figures 1-3.
Material examined: 6 specimens: ZUEC-BPO - AR785 (3 spec., 23° 32' 24'' S and 44° 47'19'' W, Sta. 11i, 17.iii.2001, 45 m), AR801 (1 spec., 23° 44' 16'' S and 45° 03' 02'' W, Sta. 6i, 15.ii.2001, 45 m), AR803 (1 spec., 23° 45' 28'' S and 45° 13' 43'' W, Sta. 23i, 22.iv.2001, 20 m), AR831 (1 spec., 23° 32' 24'' S and 44° 47' 19'' W, Sta. 86i, 15.xi.2001, 45 m).
Description: Complete specimen measuring 14.5 mm long and 1.5 mm wide (including parapodia, excluding chaetae), with 69 chaetigers (AR785). Largest specimen measuring 17.3 mm long and 2.5 mm wide, with 46 chaetigers. Smaller specimens yellowish with purple inner dorsal pigmentation, visible from parapodia 15 to 40; larger specimens with brownish transverse pigment stripes on dorsum and lobes. Prostomium longer than wide, with translucent edge; frontally rectilineal, laterally rounded and posteriorly elongated, fused dorsally at peristomium, near second chaetiger (Figure 2a). A dorsal pair of antennae, and a ventral pair of palps, both conical, frontal and about same length; palps at least two times larger than antennae. Mouth ventral, located between first and third parapodia. Body robust in anterior region, tapering towards posterior region. One pair of inflated bud-like nuchal organs, located on prostomium, near notopodial bases of first parapodia (Figure 2a). Proboscis translucent, partially everted in one specimen (AR831, Figure 2a), with 14 rows of longitudinal subdistal papillae; each row with at least six conical papillae; basal papillae smaller, enlarging towards distal part of proboscis; basal part of proboscis rough. All parapodia biramous. Anterior and median parapodia more developed than posterior parapodia. First chaetiger frontally, with well-developed, ventral cirrus laterally on prostomium and resembling a pair of lateral antennae; dorsal cirrus not seen. Neuropodial chaetae of first parapodia under prostomial frontal edge. Following parapodia lateral. Triangular preacicular notopodia; postacicular notopodia about one-third longer and more slender than preacicular. Notopodium with short foliaceous expansion, projecting dorsolaterally (Figure 2b-c). Conical dorsal cirrus longer than notopodial lobe (Figure 2b-c); filiform on posterior parapodia (Figure 2d). Interramal branchiae from parapodia 9 to 10 recurved, marginally ciliated, less developed in first parapodia (Figure 2b), occupying from one-third to one-half of interramal space on median parapodia (Figure 2c). Conical pre-acicular neuropodia, postacicular neuropodia enlarged and rounded; postacicular lobe more developed than pre-acicular lobe. Conical ventral cirrus, slightly larger than neuropodial lobe, decreasing in size towards posterior region. Notopodial and neuropodial lobes each bearing an aciculum, distally slender and curved. Pre-acicular chaetae barred (Figure 2e-f), shorter, and fewer in number than capillary postacicular chaetae; some capillary chaetae in superior and inferior positions in the bundle. Some spur-like (with a basal incision) marginally serrulate chaetae in median position in the bundle (Figure 2g); capillary postacicular chaetae marginally serrulate (Figure 2h). Number of barred chaetae decreasing towards posterior parapodia, and capillary chaetae increasing towards posterior parapodia. On posterior parapodia, notopodial and neuropodial expansion decreasing gradually in size.
Remarks: Hartman (1950) described N. acrochaeta based on an incomplete specimen measuring 50 mm long and 3 mm wide (including parapodia) from Rio Grande do Sul, Brazil, not from Uruguay, as correctly pointed out by Orensanz & Gianuca (1974) and Lana (1986). The characters observed in the present study agree with the original description. Hartman (1950) reported the presence of a vestigial dorsal cirrus on the first parapodium, but no dorsal cirrus was found in the specimens examined. Hartman (1950) also mentioned for this species 21 terminal paired papillae. However, species of Nephtys have 14, 20 or 22 pairs.
The main features to recognize this species are: a) 20 paired terminal papillae and an unpaired median papilla; 14 rows of longitudinal subdistal papillae, each row with 7-9 papillae, decreasing in size towards the basal part of the proboscis; b) branchiae starting at parapodium 9 or 10; c) dorsal and ventral lateral edge of a parapodium expanding over following parapodium, appearing overlapping or imbricated. Although it is not as evident as in N. acrochaeta, this feature is observed also in N. squamosa; d) absence of an inflated pouch (or digitate lobe) on the neuropodial superior margin; and e) barred preacicular chaetae and some capillary chaetae, not serrulate; postacicular chaetae serrulate and some chaetae spur-like, positioned inferiorly in the bundle.
Occurrence: This species, from material collected in Rio Grande do Sul, Brazil, during the Swedish Antarctic Expedition (1901-1903), was also recorded off São Paulo, from 60 to 88 m depth, in sandy-muddy bottoms with shell fragments, and in shallow bottoms (4-10 m) at Anchieta Island and Cananéia, in muddy bottoms rich in organic detritus. In this study, the specimens were collected in the shallow sublittoral, from 18.5 to 45 m depth, in bottoms of very fine to fine sand.
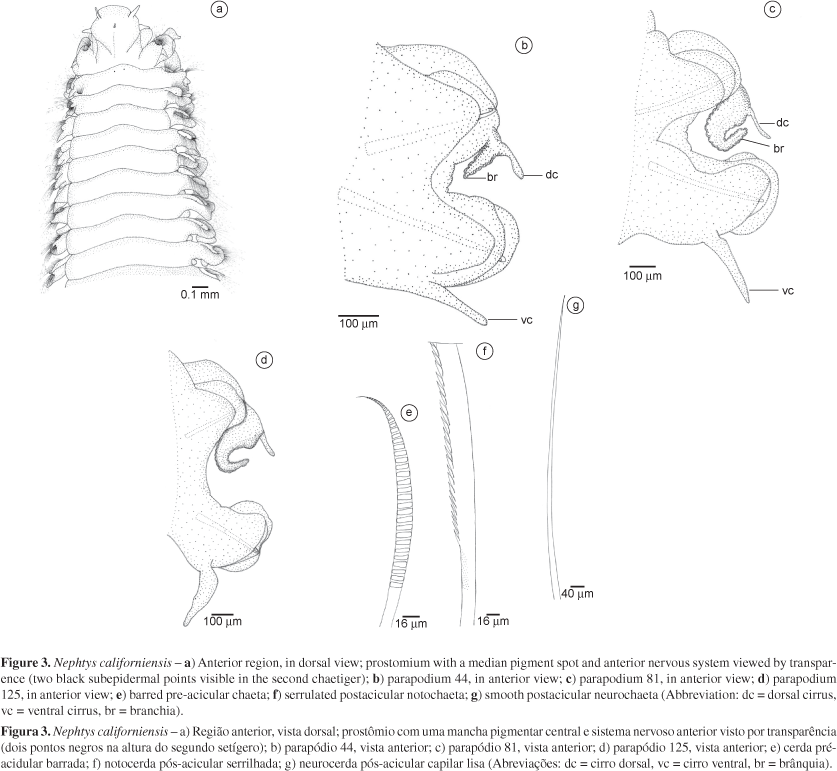
5. Nephtys californiensis Hartman 1938
(Figure 3)
Nephtys californiensis Hartman 1938: 150-151, Fig. 64; Hartman 1940: 240; Hartman 1950: 103; Imajima & Takeda 1987: 68-70, Figs 13-14; Lovell 1997: 354-356, Figure 3.
? Nephtys simoni - Lana 1986: 141, Figures 8-9 [non Perkins 1980].
Material examined: 103 specimens: ZUEC-BPO - AR297 (3 spec., 23° 41' 37.4'' S and 45° 25' 43.2''W, Sta. 199p, 18.ix.2001, intertidal), AR298 (1 spec., 23° 21'31.4''S and 44° 51' 23.2'' W, Sta. 381i, 13.xi.2001, intertidal), AR766 (2 spec., 23° 46' 29.4'' S and 45° 41' 41.4'' W, Sta. 59p, 20.viii.2001, intertidal), AR767 (1 spec., 23° 21' 23.1'' S and 44° 51' 23.7'' W, Sta. 160i, 09.v.2001, intertidal), AR768 (2 spec., 23° 22' 03.3'' S and 44° 50' 21.5'' W, Sta. 260p, 13.xi.2001, intertidal), AR769 (2 spec., 23° 21' 41.6'' S and 44° 50' 52.0'' W, Sta. 251p, 12.xi.2001, intertidal), AR770 (2 spec., 23° 38' 15.0'' S and 45° 25' 11.4'' W, Sta. 158p, 18.ix2001, intertidal), AR771 (1 spec., 23° 46' 45.1'' S and 45° 39' 51.4'' W, Sta. 33p, 20.viii.2001, intertidal), AR772 (1 spec., 23° 46' 45.1'' S and 45° 39' 51.4'' W, Sta. 47p, 20.viii.2001, intertidal), AR773 (1 spec., 23° 37' 23.9'' S and 45° 24' 21.1'' W, Sta. 218p, 19.ix.2001, intertidal), AR774 (1 spec., 23° 29' 32''S and 45° 04' 05'' W, Sta. 186i, 25.ix.2002, 5 m), AR775 (1 spec., 23° 22' 18.6'' S and 44° 50' 12.8'' W, Sta. 109p, 21.viii.2001, intertidal), AR776 (1 spec., 23° 22' 03.3'' S and 44° 50' 21.5'' W, Sta. 220p, 13.xi.2001, intertidal), AR777 (1 spec., 23° 43' 26'' S and 45° 06' 58'' W, Sta. 17i, 22.iv.2001, 35 m), AR779 (2 spec., 23° 54' 07'' S and 45° 30' 48'' W, Sta. 28i, 17.viii.2001, 25 m), AR780 (1 spec., 23° 41' 37.4'' S and 45° 25' 43.2'' W, Sta. 200p, 18.ix.2001, intertidal), AR781 (1 spec., 23° 58' 10'' S and 45° 29' 44'' W, Sta. 1i, 13.ii.2001, 45 m), AR782 (1 spec., 23° 38' 15.0'' S and 45° 25' 11.4'' W, Sta. 214p, 15.xi.2001, intertidal), AR783 (1 spec., 23° 50' 52'' S and 45° 31' 07'' W, Sta. 5i, 13.ii.2001, 5 m), AR786 (1 spec., 23° 38' 36.7'' S and 45° 25' 23.1'' W, Sta. 209p, 18.ix.2001, intertidal), AR787 (1 spec., 23° 38' 36.7'' S and 45° 25' 23.1'' W, Sta. 250p, 18.ix.2001, intertidal), AR788 (1 spec., 23° 21' 31.1'' S and 45° 51' 21.6'' W, Sta. 22p, 09.v.2001, intertidal), AR789 (2 spec., 23° 43' 26'' S and 45° 06' 58'' W, Sta. 7i, 15.ii.2001, 35 m), AR790 (3 spec., 23° 21' 31.1'' S and 45° 51' 21.6'' W, Sta. 103p, 09.v.2001, intertidal), AR791 (1 spec., 23° 46' 30.1'' S and 45° 41' 40.4'' W, Sta. 63p, 20.v.2001, intertidal), AR792 (1 spec., 23° 22' 06.2'' S and 44° 50' 17.0'' W, Sta. 92p, 21.v.2001, intertidal), AR793 (1 spec., 23° 21' 31.1'' S and 45° 51' 21.6'' W, Sta. 67p, 09.v.2001, intertidal), AR794 (1 spec., 23° 22' 18.6'' S and 44° 50' 12.8'' W, Sta. 89p, 21.viii.2001, intertidal), AR795 (3 spec., 23° 43' 26'' S and 45° 06' 58'' W, Sta. 17i, 22.iv.2001, 35 m), AR796 (1 spec., 23° 21' 23.1'' S and 44° 51' 23.7'' W, Sta. 166p, 09.v.2001, intertidal), AR798 (2 spec., 23° 41' 35.0'' S and 45° 25' 43.6'' W, Sta. 202p, 18.ix.2001, intertidal), AR800 (1 spec., 23° 41' 35.0'' S and 45° 25' 43.6'' W, Sta. 247p, 18.ix.2001, intertidal), AR802 (1 spec., 23° 46' 30.1'' S and 45° 41' 40.4'' W, Sta. 61p, 20.viii.2001, intertidal), AR804 (1 spec., 23° 22' 18.6'' S and 44° 50' 12.8'' W, Sta. 90p, 21.viii.2001, intertidal), AR806 (1 spec., 23° 22' 01.1'' S and 44° 50' 19.7'' W, Sta. 100p, 09.v.2001, intertidal), AR807 (1 spec., 23° 22' 01.1'' S and 44° 50' 19.7'' W, Sta. 93p, 09.v.2001, intertidal), AR808 (1 spec., 23° 37' 30.1'' S and 45° 23' 57.1'' W, Sta. 229p, 17.x.2001, intertidal), AR809 (1 spec., 23° 41' 27'' S and 44° 11' 42'' W, Sta. 8i, 15.ii.2001, 25 m), AR810 (1 spec., 23° 50' 52'' S and 45° 31' 07'' W, Sta. 5i, 13.ii.2001, 5 m), AR811 (3 spec., 23° 55' 26'' S and 45° 34' 35'' W, Sta. 162i, 28.vi.2002, 30 m), AR812 (4 spec., 23° 25' 31'' S and 45° 01'09''W, Sta. 119i, 26.ii.2002, 10 m), AR813 (2 spec., 23° 35' 46'' S and 45° 13' 37'' W, Sta. 154i, 24.v.2002, 5 m), AR814 (4 spec., 23° 24' 29'' S and 44° 59' 30'' W, Sta. 118i, 26.ii.2002, 10 m), AR816 (1 spec., 23° 23' 39'' S and 44° 58' 12'' W, Sta. 117i, 26.ii.2002, 15 m), AR817 (4 spec., 23° 24' 02'' S and 44° 50' 55'' W, Sta. 177i, 19.viii.2002, 17 m), AR818 (1 spec., 23° 38' 00'' S 45° 05' 58'' W, Sta. 147i, 20.v.2002, 35 m), AR819 (2 spec., 23° 29' 59'' S and 45° 09' 28'' W, Sta. 199i, 18.x.2002, 6 m), AR820 (1 spec., 23° 50' 13'' S and 45° 32' 06''W, Sta. 164i, 27.vi.2002, 15m), AR821 (3 spec., 23° 41' 57'' S and 45° 20' 25'' W, Sta. 152i, 23.v.2002, 15 m), AR822 (1 spec., 23° 55' 25'' S and 45° 29' 20'' W, Sta. 97i, 15.xii.2001, 35 m), AR824 (1 spec., 23° 32' 56'' S and 45° 00' 54'' W, Sta. 126i, 21.iii.2002, 20 m), AR825 (1 spec., 23° 30' 45'' S and 45° 04' 29'' W, Sta. 128i, 21.iii.2002, 10 m), AR827 (5 spec., 23° 26' 35'' S and 45° 00' 15'' W, Sta. 124i, 28.ii.2002, 15 m), AR828 (3 spec., 23° 54' 07'' S and 45° 30' 48'' W, Sta. 98i, 15.xii.2001, 25 m), AR829 (4 spec., 23° 31' 57'' S and 45° 08' 45'' W, Sta. 137i, 14.iv.2002, 10 m), AR830 (2 spec., 23° 33' 26'' S and 45° 08' 17'' W, Sta. 136i, 14.iv.2002, 15 m), AR832 (1 spec., 23° 50' 28'' S and 45° 29' 46'' W, Sta. 104i, 15.xii.2001, 15 m), AR834 (2 spec., 23° 29' 32'' S and 45° 04' 05'' W, Sta. 186i, 25.ix.2002, 5 m), AR836 (1 spec., 23° 25' 00'' S and 44° 01' 33'' W, Sta. 171i, 23.vii.2002, 7 m), AR837 (1 spec., 23° 22' 01.0'' S and 44° 50' 19.7'' W, Sta. 259p, 13.xi.2001, intertidal), AR838 (1 spec., 23° 50' 58'' S and 45° 14' 53'' W, Sta. 157i, 25.vi.2002, 20 m), AR839 (1 spec., 23° 55' 25'' S and 45° 29' 20'' W, Sta. 67i, 10.ix.2001, 35 m), AR840 (1 spec., 23° 32' 24'' S and 44° 47'19''W, Sta. 56i, 23.viii.2001, 45 m), AR843 (3 spec., 23° 54' 07''S 45° 30' 48'' W, Sta. 68i, 10.ix.2001, 25 m).
Description: Based on complete specimen (AR837) measuring 62.2 mm long and 2.1 mm wide (including parapodia, excluding chaetae), with 133 chaetigers. Color yellowish to whitish, with a spot of reddish pigmentation on prostomial central region and some sparse pigmentation elsewhere. Body cylindrical, uniform throughout its length, tapering near last parapodia. Prostomium longer than wide, with frontal and lateral edge rounded; frontal edge translucent (Figure 3a). Prostomial posterior edge with narrow, middorsal projection extending to second parapodium. Anterior nervous system marked by pair of dark, punctiform subdermal pigment spots on second parapodium (Figure 3a). A dorsal pair of antennae, and a ventral pair of palps, both of same length; palps with bases two times wider than bases of antennae (Figure 3a). A pair of nuchal organs dorsolaterally, on first parapodium. Parapodia biramous. First parapodium located frontally, following parapodia laterally. Proboscis totally everted in some specimens (AR792, AR802, AR807, AR836), with 22 rows of conical subdistal papillae; each row with 6-7 papillae, basally small, increasing in size towards distal end of proboscis. Twelve distal paired papillae and two unpaired papillae, one dorsal and one ventral, all conical and about the same size; long subdistal median papilla absent. Interramal branchiae from fourth parapodium, recurved, occupying almost all interramal space (Figure 3b-d). Notopodium with rounded, pre-acicular and postacicular lobes; postacicular lobe larger than pre-acicular lobe and with translucent edge on anterior to median segments (Figure 3b-d). Dorsal cirrus slender, slightly longer than branchiae (Figure 3b-d). Neuropodium with one pre-acicular lobe, smaller than postacicular lobe, with translucent edge (Figure 3b-d). Ventral cirrus foliaceous, translucent, with broader base and digitiform distal end, surpassing the neuropodial lobe in length (Figure 3b-d). Both parapodial lobes bearing one prominent aciculum. Three types of chaetae: short, barred pre-acicular (Figure 3e); flattened, long, capillary postacicular in median position of the bundle (Figure 3f); and several smooth, capillary chaetae in superior and inferior positions in the bundle (Figure 3g). Number of barred chaetae decreasing towards posterior parapodia, and number of serrulated capillary chaetae increasing. One long, slender pygidial cirrus, as long as last five parapodia together.
Remarks:Nephtys californiensis may be easily mistaken for Nephtys simoni Perkins 1980, because both have a reddish pigment spot on the prostomium, and branchiae from the third parapodium. However, several characteristics distinguish these species. Nephtys californiensis has 20 paired distal papillae and one unpaired middorsal papilla, totaling 21 conical distal papillae, all about the same size, while N. simoni has 22 paired distal papillae, all paired, and one long, unpaired middorsal papilla, totaling 23 conical distal papillae. Hartman (1938), as in the present study, did not find an unpaired middorsal papilla for N. californiensis. However, Lovell (1997) examined type-material of this species and verified the presence of one short, middorsal papilla.
In respect to the types of chaetae, N. californiensis has short barred pre-acicular chaetae, and long capillary postacicular chaetae, serrulated and flattened; whereas N. simoni has no barred chaetae, and has short capillary pre-acicular chaetae, and long capillary postacicular chaetae basally with transverse denticles and distally with larger teeth. Both N. californiensis and N. simoni have long smooth capillary chaetae. Specimens identified as N. simoni by Lana (1986) may have been mistaken for N. californiensis, since the author mentioned the presence of short, barred preacicular chaetae. The other features described by Lana (1986) agree with the specimens examined here and identified as N. californiensis.
Occurrence: This species was described from specimens collected off California (U.S.A.) in sandy bottoms, in shallow depths (<16 m). Later, it was recorded from both coasts of Mexico and from Japan, China, and Korea. The bathymetric distribution for this species is from the intertidal zone to 330 m. This is the first record of N. californiensis from the Brazilian coast. In this study, the species was collected in sandy beaches and the shallow sublittoral (<45 m), in bottoms with very fine to medium sand.
6. Nephtys squamosa Ehlers 1887
(Figure 4)
Nephtys squamosa Ehlers 1887: 128-131, pl. 37, Figs. 7-10; -Hartman 1940: 237-238, pl. 41, Figs. 98-99; Nonato & Luna 1970: 71, Figs 27-31; Fauchald 1976: 18; Taylor 1984: 35-11, Figs. 35-7-35-8; Lana 1986: 139, Figs. 4-5.
Material examined: 13 specimens: AR778 (1 spec., 23° 50' 52'' S and 45° 31' 07'' W, Sta. 30i, 15.v.2001, 5 m), AR784 (1 spec., 23° 54' 43'' S and 45° 18' 27'' W, Sta. 216i, 18.xii.2002, 16 m), AR797 (2 spec., 23° 27' 19'' S and 44° 48' 52'' W, Sta. 12i, 17.iii.2001, 35 m), AR815 (1 spec., 23° 24' 29'' S and 44° 59' 30'' W, Sta. 118i, 26.ii.2002, 10 m), AR823 (1 spec., 23° 32' 56'' S and 45° 00' 54'' W, Sta. 126i, 21.iii.2002, 20 m), AR826 (1 spec., 23° 50' 52'' S and 45° 31' 07'' W, Sta. 100i, 14.xii.2001, 5 m), AR833 (1 spec., 23° 50' 28'' S and 45° 29' 46'' W, Sta. 104i, 15.xii.2001, 15 m), AR835 (1 spec., 23° 39' 21'' S and 45° 16' 54'' W, Sta. 49i, 30.vii.2001, 15 m), AR841 (1 spec., 23° 45' 28'' S and 45° 13' 43'' W, Sta. 83i, 16.x.2001, 20 m), AR 842 (2 spec., 23° 50' 52'' S and 45° 31' 07'' W, Sta. 70i, 12.ix.2001, 5 m), AR844 (1 spec., 23° 22' 03'' S and 44° 52' 39'' W, Sta. 65i, 25.viii.2001, 10 m).
Description: Based on complete specimen (AR835) with 127 chaetigers, measuring 60 mm long and 25 mm wide (including parapodia, excluding chaetae). Body cylindrical anteriorly and rectangular posteriorly, gradually tapering towards posterior. Color whitish, with a purplish ventral longitudinal ridge. Pentagonal prostomium orangish, with long posterior V-shaped, purplish projection extending to end of second chaetiger; flattened dorsoventrally with a translucent edge, some anterior longitudinal furrows and posterior transverse ones (Figure 4a). A dorsal pair of antennae and a ventral pair of palps, antennae conical and slightly shorter than palps; palps basally enlarged (Figure 4a). Nuchal organ small, rounded, located latero-dorsally on notopodial base of first chaetiger. Mouth, opening ventrally, located between first and third chaetigers. Proboscis not evaginated. First parapodium positioned frontally, following parapodia laterally. Neuropodium of first parapodium located under prostomial lateral edge; ventral cirrus well developed, positioned laterally on prostomium, similar to second pair of lateral antennae (Figure 4a); dorsal cirrus not observed. Interramal branchiae usually recurved, or straight, laterally not ciliated; starting from third or fourth parapodium and present up to the end, occupying less than one-third of interramal space, becoming smaller on posterior parapodia (Figure 5b-c). Notopodial lobe with laterodorsal expansion well developed, translucent, imbricate on following parapodium, becoming broader in median to posterior parapodia, almost completely covering parapodial lobe. Prechaetal notopodial lobe rounded, short; acicular notopodial lobe conical, slightly longer than prechaetal lobe; postchaetal notopodial lobe well developed, with foliaceous expansion longer than acicular notopodial lobe, located posteriorly. Neuropodial expansion latero-ventrally developed, imbricate on following parapodium. Dorsal cirrus foliaceous to digitiform, slightly smaller than notopodial acicular lobe (Figure 4b-c). Postacicular neuropodial lobe, translucent, two times longer than acicular and prechaetal lobes. Ventral cirrus foliaceous, translucent, distally slender (Figure 4b-c). Parapodial lobes bearing an aciculum, distally slender and curved, protruding over anterior parapodia. Anterior parapodia with pre-acicular notochaetae and neurochaetae barred and short, as long as notopodial acicular lobe (Figure 4e), and some chaetae flattened and marginally serrulate (Figure 4d); postacicular chaetae elongated, about two times as long as pre-acicular chaetae, some cylindrical and coarse, many flattened and marginally serrulate (Figure 4g-h). Median to posterior parapodia with notochaetae and neurochaetae short and barred (Figure 4e), and short and flattened, marginally serrulate (Figure 4d); postacicular chaetae elongated and flattened, serrulate and basally cleft on one margin and slightly segmented on other margin (Figure 4f); some chaetae not cleft. Pygidium small, rounded, with dorsal anal opening and pygidial cirrus as long as last five chaetigers together.
Remarks:Nephtys squamosa is easily distinguished from its congeners in having a characteristic color pattern of purplish V-shaped patches, on the posterior edge of the prostomium, not in the center of the prostomium as do N. simoni and N. californiensis. This pattern is called "spread-eagle". Furthermore, the notopodial and neuropodial expansions are well developed and have an imbricated arrangement, similar to scales. Interramal branchiae were observed beginning on the first parapodium by Lana (1986), on the second parapodium by Nonato & Luna (1970) and on the fourth parapodium by Hartman (1950). Taylor (1984) reported that in his specimens, the branchiae arise on the third or fourth parapodia, as in the specimens examined here. In regard to the chaetae, in this species two types are mentioned: short, barred pre-acicular, or long, flattened postacicular which are marginally serrated (Hartman 1940, Nonato & Luna 1970, Taylor 1984, Lana 1986). However, in the specimens examined here, we observed other types of notochaetae and neurochaetae from the anterior to the posterior parapodia. These chaetae are short, flattened pre-acicular, marginally serrulate; or long, flattened postacicular, basally cleft (like the 'spur' chaetae in Nephtys acrochaeta Hartman 1950), but finely serrulate along one edge and slightly segmented on the other. These chaetae may not have been observed in previous studies.
Occurrence: This species has a disjunct distribution: California (Gulf of California and Catalina Island), Florida, Ecuador (Gorgona Island), Colombia, Brazil, and West Africa (Morocco). In sand (fine sand), mud, and sand-mud bottoms. From shallow bottoms to 65 m depth. In Brazil, the species has been recorded from the northeast (states of Alagoas and Sergipe, <50 m depth) southwards to Rio de Janeiro (from 12 to 197 m), São Paulo (in very fine, fine and medium sand bottoms of Dura Beach, Ubatuba, Caraguatatuba and São Sebastião; from intertidal zone to 45 m), Paraná (from 10 to 53 m) and Rio Grande do Sul.
Acknowledgments
Our thanks to the anonymous referee, to all participants of the UNICAMP (Universidade Estadual de Campinas, Instituto de Biologia, Departamento de Zoologia), and also at the CEBIMAR/USP (Centro de Biologia Marinha, Universidade de São Paulo). This work was supported by the FAPESP (Fundação de Amparo à Pesquisa, 98/07090-3, 02/04104-0) within the BIOTA/FAPESP - The Biodiversity Virtual Institute Program (www.biotasp.org.br), by the FAEPEX/UNICAMP (Fundo de Apoio ao Ensino, à Pesquisa e à Extensão), and by CNPq (Conselho Nacional de Desenvolvimento Científico e Tecnológico, 308072/2006-5). Dr. Janet Reid revised the English text.
References
FAUCHALD, K. 1976. Some Nephtyidae (Polychaeta) from Ubatuba, Brasil. Bulletin of the Southern California Academy of Sciences, 75(1):16-19.
FAUCHALD, K. & ROUSE, G. 1997. Polychaete systematics: Past and present. Zool. Scr. 26(2):71-138.
GARWOOD, P.R. & OLIVE, P.J.W. 1981. The significance of the long chaetal form of Nephtys caeca (Polychaeta: Nephtyidae) and its relationships to N. longosetosa. Sarsia, 66:195-202.
HARTMAN, O. 1938. Review of the annelid worms of the family Nephtyidae from the northeast Pacific, with descriptions of five new species. Proc. U. S. Natl. Mus., 85:143-158.
HARTMAN, O. 1940. Polychaetous Annelids. Part II. Chrysopetalidae to Goniadidae. University of Southern California Press, Los Angeles, California.
HARTMAN, O. 1948. The marine Annelids erected by Kinberg. With notes on some other types in the Swedish State Museum. Ark. Zool., 42A:1137.
HARTMAN, O. 1950. Goniadidae, Glyceridae and Nephtyidae. The University of Southern California Press, Los Angeles, California.
HILBIG, B. 1997. Family Nephtyidae Grube, 1850. pp. 317-350 in Blake, J.A., B. Hilbig, and P.H. Scott (eds). Taxonomic Atlas of the Benthic Fauna of the Santa Maria Basin and Western Santa Barbara Channel. Volume 4. The Annelida Part 1. Oligochaeta and Polychaeta: Phyllodocida (Phyllodocidae to Paralacydoniidae). Santa Barbara, Santa Barbara Museum of Natural History. Y
IMAJIMA, M. & TAKEDA, Y. 1985. Nephtyidae (Polychaeta) from Japan. I. The Genera Inermonephtys, Micronephtys and Aglaophamus. Bull. Natn. Sci. Mus., Tokyo, Ser. A, 11(2):57-90.
IMAJIMA, M. & TAKEDA, Y. 1987. Nephtyidae (Polychaeta) from Japan. II. The Genera Dentinephtys and Nephtys. Bull. Natn. Sci. Mus., Tokyo, Ser. A, 13(2):41-77.
LANA, P.C. 1986. Nephtyidae (Annelida: Polychaeta) do litoral do Estado do Paraná (Brasil). Neritica, 1(1):135-154.
LOVELL, L. 1997. A review of six species of Nephtys (Cuvier, 1817) (Nephtyidae: Polychaeta) described from the eastern Pacific. Bull. Mar. Sci., 60(2):350-363.
MARTIN, D., GIL, J. & LANA, P.C. (in press). Inermonephtys brasiliensis sp. nov. (Polychaeta: Nephtyidae) from SE Brazil, with comments on the known species of the genus. Zoosystema.
MORGADO, E.H. 1988. Anelídeos poliquetos do sublitoral da região de Ubatuba - SP, compreendida entre as ilhas Anchieta e Vitória. Tese de Doutorado, Instituto de Biologia, Universidade Estadual de Campinas.
NONATO, E. & LUNA, J. 1970. Anelídeos Poliquetas do nordeste do Brasil. I Poliquetas bentônicos da costa de Alagoas e Sergipe. Bolm. Inst. Oceanogr. S. Paulo, 19:57-130.
ORENSANZ, J.M. & GIANUCA, N.M. 1974. Contribuição ao conhecimento dos anelídeos poliquetas do Rio Grande do Sul, Brasil. I. Lista sistemática preliminar e descrição de três novas espécies. Com. Mus. Ci. PUCRGS, 4:1-37.
PAXTON, H. 1974. Contribution to the study of Australian Nephtyidae (Polychaeta). Rec. Aust. Mus., 28(7):197-208.
PERKINS, T.H. 1980. Review of species previously referred to Ceratonereis mirabilis, and descriptions of new species of Ceratonereis, Nephtys, and Goniada (Polychaeta). Proc. Biol. Soc. Wash., 93(1):1-49.
PLEIJEL, F. & DAHLGREN, T. 1998. Position and delineation of Chrysopetalidae and Hesionidae (Annelida, Polychaeta, Phyllodocida). Cladistics, 14:129-150.
RAINER, S.F. 1989. Redescription of Nephtys assimilis and N. kersivalensis (Polychaeta: Phyllodocida) and a key to Nephtys from Northern Europe. J. Mar. Biol. Ass. U.K., 69:875-889.
RAINER. S.F. & HUTCHINGS, P. 1977. Nephtyidae (Polychaeta: Erratia) from Australia. Rec. Aust. Mus., 31(8):307-347.
ROUSE, G.W. & PLEIJEL, F. 2001. Polychaetes. Oxford University Press, Oxford, U.K. 354p.
TAYLOR, J.L. 1984. Family Nephtyidae Grube, 1850. In J.M. Uebelacker & P.J. Johnson, eds. Taxonomic Guide to the Polychaetes of the Northern Gulf of Mexico. Mobile, Alabama: Barry A. Vittor, 35.1-35.20.
TEMPERINI, M.T. 1981. Sistemática e distribuição dos poliquetos errantes da plataforma continental entre as latitudes de 23°05'S e 30°00'S. MSc Thesis, Instituto Oceanográfico, Universidade de São Paulo.
WILSON, R.S. 2000. Family Nephtyidae. pp. 136-137 in Beesley, P.L., Ross, G.J.B. & Glasby, C.J. (eds.). Polychaetes & allies: The Southern synthesis. Fauna of Australia. Vol. 4A Polychaeta, Myzostomida, Pogonophora, Echiura, Sipuncula. CSIRO Publ. Melbourne xii, 465p. Y
Data Received 07/02/07
Revised 19/10/07
Published 23/11/07
ISSN 1676-0603.
- FAUCHALD, K. 1976. Some Nephtyidae (Polychaeta) from Ubatuba, Brasil. Bulletin of the Southern California Academy of Sciences, 75(1):16-19.
- FAUCHALD, K. & ROUSE, G. 1997. Polychaete systematics: Past and present. Zool. Scr. 26(2):71-138.
- GARWOOD, P.R. & OLIVE, P.J.W. 1981. The significance of the long chaetal form of Nephtys caeca (Polychaeta: Nephtyidae) and its relationships to N. longosetosa Sarsia, 66:195-202.
- HARTMAN, O. 1938. Review of the annelid worms of the family Nephtyidae from the northeast Pacific, with descriptions of five new species. Proc. U. S. Natl. Mus., 85:143-158.
- HARTMAN, O. 1940. Polychaetous Annelids. Part II. Chrysopetalidae to Goniadidae. University of Southern California Press, Los Angeles, California.
- HARTMAN, O. 1948. The marine Annelids erected by Kinberg. With notes on some other types in the Swedish State Museum. Ark. Zool., 42A:1137.
- HARTMAN, O. 1950. Goniadidae, Glyceridae and Nephtyidae. The University of Southern California Press, Los Angeles, California.
- HILBIG, B. 1997. Family Nephtyidae Grube, 1850. pp. 317-350 in Blake, J.A., B. Hilbig, and P.H. Scott (eds). Taxonomic Atlas of the Benthic Fauna of the Santa Maria Basin and Western Santa Barbara Channel. Volume 4. The Annelida Part 1. Oligochaeta and Polychaeta: Phyllodocida (Phyllodocidae to Paralacydoniidae). Santa Barbara, Santa Barbara Museum of Natural History.
- IMAJIMA, M. & TAKEDA, Y. 1985. Nephtyidae (Polychaeta) from Japan. I. The Genera Inermonephtys, Micronephtys and Aglaophamus Bull. Natn. Sci. Mus., Tokyo, Ser. A, 11(2):57-90.
- IMAJIMA, M. & TAKEDA, Y. 1987. Nephtyidae (Polychaeta) from Japan. II. The Genera Dentinephtys and Nephtys Bull. Natn. Sci. Mus., Tokyo, Ser. A, 13(2):41-77.
- LANA, P.C. 1986. Nephtyidae (Annelida: Polychaeta) do litoral do Estado do Paraná (Brasil). Neritica, 1(1):135-154.
- LOVELL, L. 1997. A review of six species of Nephtys (Cuvier, 1817) (Nephtyidae: Polychaeta) described from the eastern Pacific. Bull. Mar. Sci., 60(2):350-363.
- MARTIN, D., GIL, J. & LANA, P.C. (in press). Inermonephtys brasiliensis sp. nov. (Polychaeta: Nephtyidae) from SE Brazil, with comments on the known species of the genus. Zoosystema.
- MORGADO, E.H. 1988. Anelídeos poliquetos do sublitoral da região de Ubatuba - SP, compreendida entre as ilhas Anchieta e Vitória. Tese de Doutorado, Instituto de Biologia, Universidade Estadual de Campinas.
- NONATO, E. & LUNA, J. 1970. Anelídeos Poliquetas do nordeste do Brasil. I Poliquetas bentônicos da costa de Alagoas e Sergipe. Bolm. Inst. Oceanogr. S. Paulo, 19:57-130.
- ORENSANZ, J.M. & GIANUCA, N.M. 1974. Contribuição ao conhecimento dos anelídeos poliquetas do Rio Grande do Sul, Brasil. I. Lista sistemática preliminar e descrição de três novas espécies. Com. Mus. Ci. PUCRGS, 4:1-37.
- PAXTON, H. 1974. Contribution to the study of Australian Nephtyidae (Polychaeta). Rec. Aust. Mus., 28(7):197-208.
- PERKINS, T.H. 1980. Review of species previously referred to Ceratonereis mirabilis, and descriptions of new species of Ceratonereis, Nephtys, and Goniada (Polychaeta). Proc. Biol. Soc. Wash., 93(1):1-49.
- PLEIJEL, F. & DAHLGREN, T. 1998. Position and delineation of Chrysopetalidae and Hesionidae (Annelida, Polychaeta, Phyllodocida). Cladistics, 14:129-150.
- RAINER, S.F. 1989. Redescription of Nephtys assimilis and N. kersivalensis (Polychaeta: Phyllodocida) and a key to Nephtys from Northern Europe. J. Mar. Biol. Ass. U.K., 69:875-889.
- RAINER. S.F. & HUTCHINGS, P. 1977. Nephtyidae (Polychaeta: Erratia) from Australia. Rec. Aust. Mus., 31(8):307-347.
- ROUSE, G.W. & PLEIJEL, F. 2001. Polychaetes. Oxford University Press, Oxford, U.K. 354p.
- TAYLOR, J.L. 1984. Family Nephtyidae Grube, 1850. In J.M. Uebelacker & P.J. Johnson, eds. Taxonomic Guide to the Polychaetes of the Northern Gulf of Mexico. Mobile, Alabama: Barry A. Vittor, 35.1-35.20.
- TEMPERINI, M.T. 1981. Sistemática e distribuição dos poliquetos errantes da plataforma continental entre as latitudes de 23°05'S e 30°00'S. MSc Thesis, Instituto Oceanográfico, Universidade de São Paulo.
- WILSON, R.S. 2000. Family Nephtyidae. pp. 136-137 in Beesley, P.L., Ross, G.J.B. & Glasby, C.J. (eds.). Polychaetes & allies: The Southern synthesis. Fauna of Australia. Vol. 4A Polychaeta, Myzostomida, Pogonophora, Echiura, Sipuncula. CSIRO Publ. Melbourne xii, 465p.
Publication Dates
-
Publication in this collection
14 Mar 2008 -
Date of issue
2007
History
-
Received
07 Feb 2007 -
Reviewed
19 Oct 2007 -
Accepted
23 Nov 2007