Resumos
No presente trabalho abordam-se vários aspectos relacionados à natureza molecular da anemia falciforme (AF), desordem hematológica de caráter hereditário. A descoberta do polimorfismo do DNA no grupamento do gene betaS, originando diferentes haplótipos da doença, permitiu ampliar o conhecimento em torno da heterogeneidade clínica observada nos pacientes falcêmicos nas mais diversas regiões do mundo. Analisaram-se os diferentes haplótipos e seus parâmetros hematológicos, presentes em um grupo de 22 pacientes naturais e procedentes do estado do Ceará. A distribuição das freqüências dos haplótipos encontrados foi de 55,9% para Benin; 41,2% para República Centro-Africana (CAR); e de 2,9% para o haplótipo Senegal. Esses dados, em comparação com os demais estudos realizados no Brasil, mostram associação entre os seus valores para um alfa de 5% (p < 0,05). Os resultados obtidos possibilitam um entendimento mais completo da fisiopatologia desta doença e da sua complexidade clínica em nosso meio, bem como permitem um conhecimento mais amplo da AF em nosso país.
Anemia falciforme; Hemoglobinopatia; Globina betaS, Haplótipos
The present work deals with numerous aspects related to the molecular nature of sickle cell anemia. The discovery of the DNA polymorphism in the betas-globin gene cluster, gave origin to different haplotypes of the disease, making possible to enlarge the knowledge about the clinical heterogenity observed on the patients with sickle cell disease, in the various regions of the world. The different haplotypes and its hematological parameters were analysed in a group of 22 patients born in the State of Ceará, northeast of Brazil. The distribution found in the haplotypes frequency was of 55.9% for the Benin, of 41.2% for the CAR, and of 2.9% for Senegal haplotype. The data, compared to the others works done in Brazil, show relations among their values to alpha of 5% (p < 0,05). The results make possible a full understanding of the pathophisiology of the illness and of its clinical complexity in our State, as well as it allows a better knowledge of the sickle cell disease in our country.
Sickle cell anemia; Hemoglobinophatie; Haplotypes, betaS-globin
ARTIGO ORIGINAL ORIGINAL PAPER
Análise dos haplótipos do gene da bS-globina no Ceará
Analysis of bS-globin gene haplotypes in Ceará, Brazil
Gentil Claudino de Galiza NetoI; Maria da Silva PitombeiraII; Hélio Forta VieiraIII; Maria Laíse Chaves VieiraIV; Danúbio Andrade Bezerra FariasV
IMédico patologista clínico; mestre em Clínica Médica; médico-assistente do Serviço de Hematologia da Universidade Federal do Ceará (UFC)
IIProfessora-doutora; professora-titular do Departamento de Clínica Médica da UFC
IIIProfessor-doutor; professor titular do Departamento de Análises Clínicas da Faculdade de Fármacia, Ondontologia e Enfermagem (FFOE) da UFC
IVPrefessora-doutora; professora titular do Departamento de Análises Clínicas da FFOE/UFC
VFarmacêutico-bioquímico; funcionário adjunto do Laboratório de Biologia Molecular da FFOE/UFC
Endereço para correspondência Endereço para correspondência Gentil Claudino de Galiza Neto HEMOCE Av. José Bastos, 3.390 CEP: 60435-160 Fortaleza-CE Tel.: (85) 3433-4400 e-mail: gentilgaliza@ig.com.br
RESUMO
No presente trabalho abordam-se vários aspectos relacionados à natureza molecular da anemia falciforme (AF), desordem hematológica de caráter hereditário. A descoberta do polimorfismo do DNA no grupamento do gene bS, originando diferentes haplótipos da doença, permitiu ampliar o conhecimento em torno da heterogeneidade clínica observada nos pacientes falcêmicos nas mais diversas regiões do mundo. Analisaram-se os diferentes haplótipos e seus parâmetros hematológicos, presentes em um grupo de 22 pacientes naturais e procedentes do estado do Ceará. A distribuição das freqüências dos haplótipos encontrados foi de 55,9% para Benin; 41,2% para República Centro-Africana (CAR); e de 2,9% para o haplótipo Senegal. Esses dados, em comparação com os demais estudos realizados no Brasil, mostram associação entre os seus valores para um alfa de 5% (p < 0,05). Os resultados obtidos possibilitam um entendimento mais completo da fisiopatologia desta doença e da sua complexidade clínica em nosso meio, bem como permitem um conhecimento mais amplo da AF em nosso país.
Unitermos: Anemia falciforme, Hemoglobinopatia, Globina bS, Haplótipos
ABSTRACT
The present work deals with numerous aspects related to the molecular nature of sickle cell anemia. The discovery of the DNA polymorphism in the bs-globin gene cluster, gave origin to different haplotypes of the disease, making possible to enlarge the knowledge about the clinical heterogenity observed on the patients with sickle cell disease, in the various regions of the world. The different haplotypes and its hematological parameters were analysed in a group of 22 patients born in the State of Ceará, northeast of Brazil. The distribution found in the haplotypes frequency was of 55.9% for the Benin, of 41.2% for the CAR, and of 2.9% for Senegal haplotype. The data, compared to the others works done in Brazil, show relations among their values to alpha of 5% (p < 0,05). The results make possible a full understanding of the pathophisiology of the illness and of its clinical complexity in our State, as well as it allows a better knowledge of the sickle cell disease in our country.
Key words: Sickle cell anemia, Hemoglobinophatie, Haplotypes, bS-globin
Introdução
A anemia falciforme (AF) é uma das doenças hematológicas herdadas mais comuns em todo o mundo, atingindo grande número de indivíduos dos mais diferentes países. Ela surgiu nos países do centro-oeste africano, da Índia e do leste da Ásia há cerca de 50 mil a 100 mil anos, entre os períodos paleolítico e mesolítico(17, 25). O fato que motivou a mutação do gene da hemoglobina normal (HbA) para o da hemoglobina S (HbS) ainda permanece desconhecido.
A simples substituição pontual de uma base nitrogenada, timina por adenina (GAT  GTT), no sexto códon do éxon 1 no DNA do cromossomo 11 (Figura 1) ocasiona o surgimento de uma hemoglobina patológica(3). A troca de bases nitrogenadas no DNA, ao invés de codificar a produção (transcrição) do aminoácido ácido glutâmico, irá, a partir daí, determinar a produção do aminoácido valina, que entrará na posição 6 da seqüência de aminoácidos que compõem a cadeia beta da hemoglobina, modificando sua estrutura molecular(4).
GTT), no sexto códon do éxon 1 no DNA do cromossomo 11 (Figura 1) ocasiona o surgimento de uma hemoglobina patológica(3). A troca de bases nitrogenadas no DNA, ao invés de codificar a produção (transcrição) do aminoácido ácido glutâmico, irá, a partir daí, determinar a produção do aminoácido valina, que entrará na posição 6 da seqüência de aminoácidos que compõem a cadeia beta da hemoglobina, modificando sua estrutura molecular(4).
Em 1978, Kan e Dozzy publicaram a primeira descrição da existência de duas mutações distintas para a hemoglobina S detectadas pela ação da enzima de restrição Hpa I no sítio de 5kb a 3 do gene beta, estabelecendo o conceito de origem multicêntrica para esta doença. Estudos subseqüentes definiram os três principais haplótipos para a anemia falciforme: CAR (República Centro-Africana), Benin e Senegal, originados em regiões distintas do continente africano(18), e um outro haplótipo, também de origem independente, foi descrito em populações provenientes da península arábica e da Índia(13). A variabilidade genética em torno da mutação permitiu uma melhor compreensão da heterogeneidade clínica da doença, além de ter importância para o estudo antropológico(28).
O tipo de variabilidade mais comum no complexo gênico das globinas alfa ou beta é aquele produzido por variações de seqüências que alteram o sítio de reconhecimento de uma enzima de restrição. Essas alterações ocorrem aproximadamente a cada cem bases ao longo do genoma. O padrão de combinação dos sítios polimórficos para qualquer cromossomo é chamado de haplótipo(2). O primeiro polimorfismo associado ao gene bS foi descrito, como referido acima, por Kan e Dozy, em 1978, no sítio para a enzima de restrição Hpa I localizado na posição 3 do gene, seguindo-se novas descrições de outros sítios de polimorfismos de restrição(1, 14).
Foram descritos 17 sítios polimórficos (Figura 2) no complexo dos genes da globina alfa e mais de duas dezenas no complexo da globina beta(2). A descoberta dos haplótipos do gene bS apresentou-se como importante elemento de análise antropológica para estudo das composições populacionais, bem como elementos de estudo clínico, os quais podem fornecer dados preditivos acerca da evolução da doença e de seu nível de gravidade(20).
Os haplótipos do gene bS têm papel importante na regulação variável da síntese da Hb fetal, na relação final entre as concentrações de HbS e HbF no adulto e na taxa de redução da HbF durante a infância. A concentração de HbF está aumentada nos haplótipos Senegal e asiático (árabe) e decresce nos haplótipos CAR e Benin, devido a uma taxa de translação no sentido 5 para 3 e de substituição da HbF por HbS mais lenta nos dois primeiros que nos dois últimos haplótipos(20).
No presente trabalho foram determinados os haplótipos do gene da bS-globina num grupo de pacientes com AF, todos oriundos do estado do Ceará, esperando assim contribuir para um melhor entendimento de alguns aspectos dessa doença em nosso meio.
Material e métodos
Participaram deste estudo 22 pacientes com diagnóstico clínico e laboratorial de AF (forma homozigótica SS) atendidos no ambulatório do Serviço de Hematologia do Hospital Universitário Walter Cantídio, da Universidade Federal do Ceará (UFC), de ambos os sexos, todos nascidos no estado do Ceará, com seus antepassados até a segunda ascendência (pais e avós) igualmente naturais desse estado. Outro critério de inclusão foi a ausência de tratamento transfusional com concentrado de hemácias por um período mínimo de 90 dias antes da coleta das amostras sangüíneas para a realização dos exames laboratoriais. Foram coletados 5ml de sangue de cada um dos pacientes em tubos de tipo vacutainer, contendo etilenedinitrilotetraacetato (EDTA). O estudo hematológico foi procedido em contador automatizado, marca Coulter, modelo T-890, sendo analisados os seguintes parâmetros: número absoluto de hemácias (Hm), concentração de hemoglobina (Hb), determinação de hematócrito (Ht), volume corpuscular médio (VCM), hemoglobina corpuscular média (HCM), concentração de hemoglobina corpuscular média (CHCM).
O estudo dos haplótipos da mutação bS no gene da cadeia betaglobínica foi realizado através da técnica da reação em cadeia mediada pela polimerase (PCR) e da digestão com enzimas de restrição específicas, seguindo as recomendações contidas no trabalho de Sutton(23), com modificações. Depois de realizada a amplificação do material genômico, realizou-se digestão enzimática utilizando-se as seguintes enzimas: Hind III, Hinc II, Ava II e Hinf I, das marcas Promega e Gibco, para determinar os haplótipos mais freqüentes: CAR (Bantu), Benin (Ben) e Senegal (Sen).
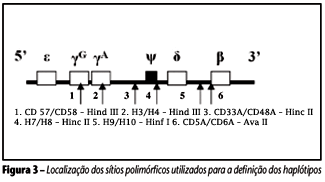
Foram estudados os seguintes sítios gênicos, respectivamente: Hind III na região IVS2 do gene gA e na região IVS2 do gene gG; Hinc II em yb e na extremidade 3 de b; Ava II na região IVS2 de b; e Hinf 1 na extremidade 5 de b(23). As posições desses sítios de polimorfismos gênicos situados no complexo gênico da betaglobina estão representadas na Figura 3.
Resultados
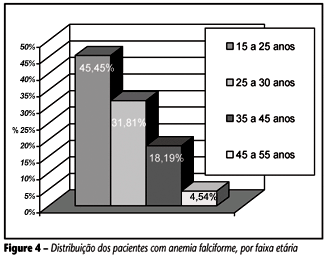
Os resultados obtidos em 22 pacientes portadores de Hb SS mostraram que a faixa etária situou-se entre 15 e 51 anos, com idade média de 29,33 ± 11,92 anos (média e desvio-padrão), sendo que o grupo masculino (n = 9) teve média de idade de 28,87 ± 13,02 anos e o feminino (n = 13), 30,36 ± 9,75 anos. Em relação à faixa etária, observamos que 77,26% dos pacientes têm idade entre 15 e 30 anos (Figura 4).
A Tabela 1 exibe os valores hematológicos médios e os índices hematimétricos de todos os pacientes (Hb, Ht, VCM, HCM, CHCM e HbF). O VCM mostrou uma média de 105,57 ± 10,05fl, demonstrando acentuada macrocitose (VCM > 100fl) para ambos os sexos, sem diferença estatística entre eles. A concentração de Hb fetal para todo o grupo foi de 7,61%, tendo concentração em nível mais elevado no grupo feminino, com 8,49%, em relação ao masculino, com 7,44%. Entretanto, essa diferença não teve significância estatística.
O padrão dos sítios polimórficos, conforme definido e apresentado na Figura 3, caracterizou os diferentes haplótipos de acordo com a presença (+) ou ausência (-) de resposta aos cortes enzimáticos (Figura 5). A caracterização do haplótipo CAR foi definida como [+ - - - + -]; o haplótipo Benin foi definido pela seqüência [- - - + + -]; e o haplótipo Senegal o foi pela combinação [+ - + + + +].
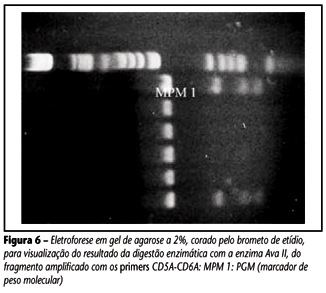
Os casos de heterozigose (Figura 6) para dois haplótipos definiram-se, de acordo com as observações anteriores de Kulozik et al.(13) e Rieder et al.(22), pela presença de um haplótipo comum, presente em combinação com um haplótipo raro, ao invés de dois haplótipos raros. No presente trabalho observou-se a presença de cinco haplótipos com a combinação [- - - - + -], sem possibilidade de serem definidos.
A distribuição geral dos genótipos para o grupo de pacientes encontra-se disposta na Figura 5. Os genótipos mais freqüentes foram Ben/Ben e CAR/Ben, ambos com 27,27%. Foram evidenciadas a ocorrência de um paciente heterozigoto para o haplótipo Senegal, o qual tem baixa freqüência no Brasil, e a presença de haplótipos indeterminados em 22,72% dos pacientes. Em relação à freqüência dos haplótipos estudados por cromossomos, observou-se a presença, em maior número, do Benin, com 43,18%, seguido pelo CAR, com 31,82%, como exposto na Tabela 2.
Os valores médios dos dados laboratoriais dos pacientes pertencentes aos haplótipos CAR, Benin e Senegal estão distribuídos na Tabela 3. Notam-se as concentrações mais elevadas de Hb (8,9g/dl) e de HbF (13,4%) para os pacientes do haplótipo Senegal.
Discussão
Estes resultados representam a primeira análise dos haplótipos da AF em pacientes naturais do Ceará. O estudo dos polimorfismos gênicos pode ser utilizado com diferentes objetivos: para a determinação da origem unicêntrica ou multicêntrica de uma mutação, para discriminar eventos epistáticos (quando outros genes interferem na expressão fenotípica do gene mutante) e para definir o caminho de fluxo de um gene específico mutante. Quando os dados e informações estão suficientemente consolidados, como no caso do grupamento do gene beta mutante, os haplótipos podem ser úteis no estudo da origem e evolução da raça humana(15).
A descrição da existência de duas mutações distintas para a HbS detectadas pela ação da enzima de restrição Hpa I no sítio de 5kb a 3 do gene beta estabeleceu o conceito de múltipla origem para esta doença(12). Estudos subseqüentes definiram os principais haplótipos para a anemia falciforme na África: CAR, Benin e Senegal(18), e um outro haplótipo foi descrito em populações provenientes da península arábica e da Índia(13). A diversidade genética assim observada permitiu a compreensão da heterogeneidade clínica da doença, além de contribuir para o estudo antropológico(28).
No Brasil, vários estudos dos haplótipos relacionados a AF têm sido realizados (Tabela 4). A distribuição das freqüências dos haplótipos em Belém, no Pará, foi de 66,7% para o haplótipo CAR, 30% para o Benin e 3,3% para o Senegal(19). Em Salvador, na Bahia, a distribuição das freqüências dos haplótipos foi de 55% para o CAR, 45% para o Benin e não houve ocorrência do haplótipo Senegal(8). Na região de Campinas, São Paulo, a distribuição das freqüências dos haplótipos foi de 64,7% para o CAR, 35,2% para o Benin e não houve ocorrência do haplótipo Senegal(9). Em Ribeirão Preto, São Paulo, a distribuição dos haplótipos foi de 73% para o CAR, 25,5% para o Benin e 1,5% de ocorrência do Senegal(26). A distribuição das freqüências dos haplótipos na cidade de Porto Alegre, no Rio Grande do Sul, foi de 79,6% para o haplótipo CAR, 18,4% para o Benin e 2% para o Senegal(24).
A distribuição das freqüências dos haplótipos encontrados, considerando apenas os que puderam ser determinados, para este grupo (Ceará) foi de: 55,9% para o haplótipo Benin, 41,2% para o CAR e 2,9% para o Senegal. Os nossos dados, em comparação com os dos demais estudos realizados no Brasil, mostram associação entre os seus valores para um alfa de 5% (a = 0,05), com diferenças estatisticamente significantes (c2 = 17,02/p < 0,05).
Avaliando qualitativamente esses resultados, observa-se uma tendência para a presença em maior número do haplótipo Benin em relação ao CAR, divergindo das freqüências encontradas nas cidades de Belém, Campinas, Ribeirão Preto e Porto Alegre, onde o haplótipo CAR é preponderante (Tabela 4). Entretanto assemelham-se aos encontrados na cidade de Salvador, e isso pode ser devido ao fato de essa cidade ter sido importante entreposto do comércio escravagista, tornando-se uma fonte importante de fornecimento de escravos para o Ceará(10).
O paciente portador do haplótipo Senegal, identificado neste estudo, mostra alguns valores hematológicos distintos dos encontrados para as médias dos indivíduos com outros haplótipos. As principais diferenças devem-se ao valor mais elevado da Hb (c = 8,9g/dl) e da HbF (c = 13,4%). Entretanto, o valor do VCM (c = 92 fL) para esse indivíduo, ao contrário, ficou abaixo dos resultados observados nos demais haplótipos. Esses valores são compatíveis com o descrito por Nagel et al.(16), que, estudando 60 indivíduos com pelo menos um cromossomo pertencente ao haplótipo Senegal, evidenciaram uma concentração de Hb de 9,6 ± 1,1g/dl, VCM de 91,2 ± 9,2% e concentração de Hb fetal igual a 9,9 ± 5,4%.
As análises hematológicas não apresentaram diferenças em relação aos valores usualmente encontrados e amplamente descritos na literatura(21) para os portadores da hemoglobinopatia SS. A exceção deveu-se à presença de valores aumentados para o VCM, principalmente relacionados ao haplótipo CAR. Sabe-se que macrocitose faz parte do quadro laboratorial da anemia falciforme em decorrência da marcante produção de reticulócitos e da presença, em grande número, das células densas, sendo agravada por uma deficiência no aporte nutricional e medicamentoso (de reposição) do substrato ácido fólico.
Não encontramos dados na literatura associando os haplótipos do gene bS ao aumento do VCM. Não podemos descartar, ainda, a possibilidade de termos, por conta da pouca contribuição dos povos mediterrâneos na formação étnica da população cearense, uma diminuta associação de anemia falciforme com talassemia alfa. Sabe-se que o valor do VCM observado nessa hemoglobinopatia é menor que o encontrado na AF(7). A associação dessas duas patologias foi descrita pela primeira vez numa família da Arábia Saudita, por Wheatherall, em 1969(21), e no Brasil foi identificada em pacientes de São Paulo, em 22%(5) e 17%(8). A menor associação da talassemia alfa com a anemia falciforme no nosso meio poderia se expressar pela falta do seu efeito redutor no VCM dos pacientes.
A média da concentração de hemoglobina fetal para o grupo de pacientes foi de 7,61 ± 1%, e equiparou-se ao observado nos demais estudos da literatura. A concentração média de HbF encontrada por Queiroz(21), na Bahia, foi de 6,45%. Em São Paulo, Figueiredo(8) encontrou uma HbF de 6,35%, e em Alagoas, Costa(6) encontrou concentração média de HbF de 9,28%.
Os estudos dos haplótipos realizados neste grupo de pacientes constituem fonte pioneira de dados referentes ao diagnóstico molecular da anemia falciforme no estado do Ceará. Contribui-se dessa forma com mais informações sobre a doença e a expressão do seu polimorfismo genético, para serem analisadas em conjunto com os demais dados provenientes de outras localidades e regiões do Brasil.
Além disso, nesse trabalho indica-se aparente predominância do grupo Benin em nosso meio, o que parece confirmar as informações obtidas através dos documentos históricos sobre nossa origem étnica. Os dados hematológicos avaliados junto ao contexto molecular da anemia falciforme serão referência para o aprofundamento do conhecimento clínico da doença, bem como valioso adjuvante no seguimento clínico dos pacientes ao longo do tempo.
Primeira submissão em 03/01/05
Última submissão em 03/01/05
Aceito para publicação em 22/08/05
Publicado em 20/10/05
Trabalho realizado junto ao Departamento de Clínica Médica da UFC, com apoio da Fundação Cearense de Apoio ao Desenvolvimento Científico e Tecnológico (FUNCAP) e do Centro de Hematologia e Hemoterapia do Ceará (HEMOCE), e baseado na dissertação de mestrado Estudo dos haplótipos da anemia falciforme e sua relação com as origens étnicas da população negra no Ceará, do ano de 2001, apresentada na UFC.
- 1. ANTONARAKIS, S. E.; BOEHM, C. D.; GIARDINA, P. J. V.; KAZAZIAN Jr., H. H. Nonrandon association of polymorphic restriction sites in the b-globin gene cluster. Proc Natl Acad Sci, v. 79, p. 137-41, 1982.
- 2. ANTONARAKIS, S. E.; KAZAZIAN Jr., H. H.; ORKIN, S. H. DNA polymorphism and molecular pathology of the human globin gene clusters. Hum Genet, v. 69, p. 1-14, 1985.
- 3. BALLAS, S. K.; MOHANDAS, N. Pathophisiology of vaso-occlusion. In: CHARACHE, S.; JONHSON, C. S. (eds.). Hematology/Oncology Clinics of North America Sickle Cell Disease, v. 10, n. 6, p. 1221-40, 1996.
- 4. BUNN, H. F.; FORGET, B. G. Hemoglobin: molecular, genetic and clinical aspects. 1. ed. W.B. Saunders Company, 1986. 690 p.
- 5. COSTA, F. F. et al. bS: gene-cluster haplotypes in sickle cell anemia from two regions of Brazil. Am J Hematology, v. 46, p. 96-7, 1993.
- 6. COSTA, P. J. M. S. Anemia falciforme: diversidade clínica e laboratorial em crianças e adolescentes portadores do haplótipo Bantu do gene da globina bS 1999, p. 64. Dissertação (Mestrado em Saúde da Criança) Universidade Federal de Alagoas, Maceió.
- 7. CHARACHE, S.; LUBIN, B.; REID, C. D.(ed.). Management and therapy of sickle cell disease. U.S. Department of Health and Human services Public Health Services National Institute of Health, August,1991.
- 8. FIGUEIREDO, M. S. Efeitos da talassemia alfa e dos haplótipos do complexo da globina beta nas alterações clínicas e laboratoriais da anemia falciforme no Brasil. 1993. 79 p. Tese (Doutorado) Escola Paulista de Medicina, Universidade Federal de São Paulo, São Paulo.
- 9. GONÇALVES, M. S. et al. Sickle cell disease in a brazilian population from São Paulo: a study of the bS haplotypes. Hum Hered, v. 44, p. 322-7, 1994.
- 10.GIRÃO, R. A abolição no Ceará. Imprensa Oficial do Ceará, Secretaria de Cultura Turismo e Desporto,1984. Capítulo III, p. 47-55.
- 11. HOFFBRAND, A. V.; PETTIT, J. E. Essential Haematology 3. ed. London, UK: Blackwell Science, 1993. 437p.
- 12. KAN, Y. M.; DOZY, A. M. Polymorphism of DNA sequence adjacent to human beta globin structural gene: relationship to sickle mutation. Proc Natl Acad Sci, v. 75, p. 5631-5, 1978.
- 13. KULOZIK, A. E. et al. Geographical survey of bS globin gene haplotypes: evidence for an independent Asian origin of the sickle-cell mutation. Am J Hum Genet, v. 39, p. 239-44, 1986.
- 14.MEARS, J. L. et al. Th sickle gene polymorphism in North Africa. Blood, v. 58, p. 599-601, 1981.
- 15. NAGEL, R. L.; RANNEY, H. M. Genetic epidemiology of structural mutations of the beta globin gene. Seminars in Hematology, v. 27, p. 342-59,1990.
- 16. NAGEL, R. L. et al. The Senegal DNA haplotype is associated with amelioration of anemia in Afican-American sickle cell anemia patients. Blood, v. 77, n. 6, p. 1371-5,1991.
- 17. NAOUM, P. C. Hemoglobinopatias e talassemias. Ed. Sarvier, 1997.
- 18. PAGNIER, J. et al. Evidence for the multicentric origin of the sickle cell hemoglobin gene in Africa. Proc Natl Acad Sci, v. 81, p. 1771-3, 1984.
- 19. PANTE-DE-SOUZA, G. et al. Origin of the hemoglobin S gene in a northern Brazilian population: the effect of slave trade and internal migrations. Genet Mol Biol, v. 21, n. 4, p. 1-10, 1998.
- 20. POWARS, R. D. bS-gene-cluster haplotypes in sickle cell anemia. In: NAGEL, R. L. (ed.). Hematology/Oncology Clinics of North America Hemoglobinopathies, v. 5, n. 3, p. 475-93, 1991.
- 21. QUEIROZ, I. M. L. P. Características clínicas, hematológicas e genéticas em pacientes homozigotos para a hemoglobinopatia S da Bahia e de São Paulo. 1996. 111p. Dissertação (Mestrado em Hematologia) Escola Paulista de Medicina, Universidade Federal de São Paulo, São Paulo.
- 22. RIEDER, R.F. et al. Effect of beta-globin gene cluster haplotype on the hematological and clinical features of sickle cell anemia. Am J Hemat, v. 36, p. 184-9, 1991.
- 23. SUTTON, M.; BOUHASSIRA, E.; NAGEL, R. L. Polymerase chain reaction amplification applied to the determination of the b-like globin gene cluster haplotypes. Am J Hematology, v. 32, p. 66-9, 1989.
- 24. WAGNER, S. C. et al. Caracterização molecular da anemia falciforme em pacientes de Porto Alegre. Rev Bras de Genet, v. 19(suppl.): 244 (abstract), 1996.
- 25. WHO. Working Group Hereditary anemias: genetics basis, clinical features, diagnosis and treatment. Bull WHO, v. 60, p. 643-60, 1982.
- 26. ZAGO, M. A.; FIGEIREDO, M. S.; OGO, S. H. Bantu bS cluster haplotype predominantes among Brasilian blacks. Am J Phys Anthropol, v. 88, p. 295-8, 1992.
- 28. ZAGO, M. A. Origem e heterogeneidade da anemia falciforme. Boletim, v. XV, n 162, p. 3-8, 1993.
Datas de Publicação
-
Publicação nesta coleção
10 Fev 2006 -
Data do Fascículo
Out 2005
Histórico
-
Recebido
22 Ago 2005