Abstract
The present study was conducted with eight cultivars of soybean (Glycine max) to determine the effect of exclusion of solar UV-B on the vegetative growth (plant height, leaf area, no. of nodes), UV absorbing compounds (implicated in UV protection) and crop yield (No. of pods and seed weight) and to determine the cultivar difference in their sensitivity to ambient UV-B. Exclusion of solar UV-B enhanced the vegetative growth and yield of all the soybean cultivars. The results showed a significant inverse association between the enhancement in vegetative growth and number of pods among the cultivars tested, indicating differences in the carbon partitioning amongst the cultivars by the way of exclusion of solar UV-B. NRC-7, Pusa-24 and JS-335 showed maximum promotion in vegetative growth and less enhancement in crop yield after solar UV-B exclusion. Kalitur, JS71-05, Hardee, PK-472 and PK-1029 showed improved performance both in terms of number of pods/plant and seed weight after solar UV-B exclusion. An enhancement in the crop yield by exclusion of solar UV-B indicated poor response of the cultivar to the ambient solar UV-B; these cultivars would be more suitable at latitudes, which received less UV-B. According to UV-SI, sensitivity of eight Indian soybean cultivars to ambient level of UV-B (280-320 nm) radiation had the following descending order; PK-472 > JS-335 > Hardee > Kalitur > JS71-05 > Pusa-24 > NRC-7 > PK-1029. These findings suggest the way to select the best-suited cultivar for particular latitude.
Exclusion; Leaf Area; Sensitivity index; UV-Absorbing substances
RESEARCH ARTICLE
Intraspecific variation in sensitivity to ambient ultraviolet-B radiation in growth and yield characteristics of eight soybean cultivars grown under field conditions
Sanjay S.BaroniyaI; Sunita KatariaII, * * Corresponding author: sunitakataria@hotmail.com ; G.P. PandeyII; Kadur N.GuruprasadII
IGovt. Shri Krishnaji Rao Pawar Post Graduate College, Dewas-455001, (M.P.)
IIPhotobiology lab, School of Life Sciences, Khandwa Road Campus, Devi Ahilya University, Indore - 452017, M.P
ABSTRACT
The present study was conducted with eight cultivars of soybean (Glycine max) to determine the effect of exclusion of solar UV-B on the vegetative growth (plant height, leaf area, no. of nodes), UV absorbing compounds (implicated in UV protection) and crop yield (No. of pods and seed weight) and to determine the cultivar difference in their sensitivity to ambient UV-B. Exclusion of solar UV-B enhanced the vegetative growth and yield of all the soybean cultivars. The results showed a significant inverse association between the enhancement in vegetative growth and number of pods among the cultivars tested, indicating differences in the carbon partitioning amongst the cultivars by the way of exclusion of solar UV-B. NRC-7, Pusa-24 and JS-335 showed maximum promotion in vegetative growth and less enhancement in crop yield after solar UV-B exclusion. Kalitur, JS71-05, Hardee, PK-472 and PK-1029 showed improved performance both in terms of number of pods/plant and seed weight after solar UV-B exclusion. An enhancement in the crop yield by exclusion of solar UV-B indicated poor response of the cultivar to the ambient solar UV-B; these cultivars would be more suitable at latitudes, which received less UV-B. According to UV-SI, sensitivity of eight Indian soybean cultivars to ambient level of UV-B (280-320 nm) radiation had the following descending order; PK-472 > JS-335 > Hardee > Kalitur > JS71-05 > Pusa-24 > NRC-7 > PK-1029. These findings suggest the way to select the best-suited cultivar for particular latitude.
Key words: Exclusion, Leaf Area, Sensitivity index, UV-Absorbing substances.
INTRODUCTION
A wealth of research effort during the past decades has been directed towards quantifying the potential effects of UV-B fluxes on plant parameters (Madronich et al., 1995; Guruprasad et al., 2007). Numerous studies have investigated the effects of elevated UV-B on plants, and have shown a diverse range of responses, including changes at the physiological, morphological, biochemical and molecular levels (Carletti et al., 2003; Paul and Gwynn-Jones, 2003; Guruprasad et al., 2007). These studies have shown deleterious effects of UV-B such as reduced photosynthesis, biomass reduction, decreased protein synthesis, damage to nucleic acids and lipids (Jansen et al., 1998; Rathore et al., 2003).
The level of ambient UV-B radiation in sunlight varies with reference to latitude and is relatively higher in tropical regions than in temperate regions. In tropics, due to small solar zenith angle and the thin stratospheric ozone layer, terrestrial plants encounter much higher level of UV-B radiation than at higher latitudes (Caldwell et al., 1989; Madronich et al., 1995). Plants in their need to capture sunlight for photosynthesis are unavoidably exposed to higher amount of UV-B radiation under tropical conditions. Soybean crop introduced to India in 1960s is being propagated at a fast rate and several new cultivars are being released. The optimum performance of a variety apart from other factors would also depend on its tolerance to solar UV-B.
An assessment of the impact of solar UV-B at given latitude is possible by growing plants after the exclusion of this radiation by using specific filters. The studies carried out by different laboratories using this approach have generally focused on UV-B impacts on plant growth and morphology and their results have been mixed. Caldwell (1968) did not find any significant effect of solar UV-B exclusion on the naturally occurring plants of alpine plant community. Tezuka et al., (1993) obtained a transient reduction in biomass accumulation in tomato plants by excluding solar UV-B in field experiments carried out in Japan, whereas Becwar et al., (1982) did not detect any effect of UV-B exclusion at a high elevation site in Colorado on final biomass of plants of potato, radish and wheat. Such exclusion studies in radish have shown an enhancement in leaf growth but decrease in the yield (Zavala and Botto, 2002). An increase in biomass and yield by exclusion of solar UV-B has also been observed in tropical crops like Cyamopsis and Vigna (Amudha et al., 2005) and in an Indian variety of soybean (JS-71-05) (Varalakshmi et al., 2003; Guruprasad et al., 2007).
Increase in the UV-B radiation is known to reduce biomass, leaf area, yield and photosynthesis (Golaszewska et al., 2003; Jayakumar et al., 2004; Pal et al., 2006; Guruprasad et al., 2007) and increased UV absorbing compound (Flint et al., 2004). Plants are more resistant to UV-B then are microbes or mammalian cells, because they produce a variety of secondary metabolites that effectively absorb UV-B and prevent it from penetrating leaf mesophyll (Caldwell et al., 1995; Mazza et al., 2000).
Responses of plants to UV-B vary not only among the species, but also among the varieties of same species (Tevini, 2000). Variations in responsiveness of different species and varieties to UV-B were also reported for a variety of plant species (Smith et al., 2000; Yanqun et al., 2003; 2004; Li et al., 2010). Smith et al. (2000) concluded that variations in UV-B sensitivity between different species represent the relative contribution of morphological, physiological and biochemical differences, but variations within species are usually more subtle.
In the present investigation eight Indian soybean cultivars have been tested for their response to ambient level of UV-B at Indore (Latitude-22.4ºN) India.
MATERIALS AND METHODS
Seeds of soybean (Glycine max) cultivars: PK 472, PK 1029, Pusa 24, JS 71-05, JS 335, Hardee, NRC 7 and Kalitur were obtained from Directorate of Research for Soybean, Indore (M.P.). The seeds, graded by weight in order to ensure uniform seed size within a cultivar by excluding the heaviest and lightest 10% of the seeds, were surface sterilized and then inoculated with a slurry of Rhizobium japonicum strain before sowing in the field area of 120 x 90 cm in three feet rows planted 0.3m apart with 0.06m plant spacing within the row under iron cages of dimensions [4 feet L × 3 feet W × 3 feet H]. All the field experiments were carried out during Rabi season in the Botanical garden of School of Life Sciences, Devi Ahilya University, Indore, India. The cages received full solar radiation for most of the day without shading. The plots were irrigated regularly to avoid water stress. Temperatures both inside and outside each enclosure where monitored daily using max/min thermometers. The average daily temperature at outside raised from 25° to 27° C during the growing period of October 2009 to January 2010. The temperature inside the cages never differed more than 2° C from the external air temperature.
For the exclusion of UV-B, sunlight was filtered through Polyester cut off filters (Garware Polyester Limited, Mumbai), these filters were wrapped on iron cages in which the plants were grown. The plants grown under UV-B exclusion filters were compared to control plants (ambient UV-B) growing in the cages, which were covered by polythene that transmits all the UV radiation in sunlight. The transmission/exclusion characteristics of these filters were measured by Shimadzu (UV-160) spectrophotometer (Figure 1).
Absolute solar irradiance without UV-B was measured using a radiometer (IL 1350, International light Inc) U.S.A. The ambient solar irradiance during experimental period at midday was 10 W/m2/ s the loss in light intensity at midday by -UV-B filters was 17% (8.3 W/m2/ s) and 20% (8 W/m2/ s) under UV-A/B filter and 5% (9.5 W/m2/ s), under polythene filter transmissible to UV (Control). The PAR intensity for normal plant growth was observed to be optimal saturating light.
Plant height, number of nodes/plant, number of pods, seed yield (g/plant) have been taken at the crop maturity in three replicates of five plants each. Plant population was uniform at the time of maturity of crop. Leaf area and UV absorbing substances (UAS) were monitored in fully matured third trifoliate leaves at fifth node. Leaf discs of 100 mm2 were taken and these discs were extracted in 10 ml of acidified methanol (79:20:1 v/v methanol, water, HCl) for measuring UV absorbing compounds, according to the procedure of Mirecki and Teramura, (1984). The absorbance was taken at 300 nm using Shimadzu Spectrophotometer.
Differences in the UV-sensitivity of the eight cultivars were ascertained by a UV sensitivity index (UV-SI) which was calculated according to the following equation (Saile-Mark and Tevini, 1997):

An UV-tolerant plant has an UV-SI of 3, whereas UV-SI values below 3 indicate an UV-sensitive plant
RESULTS
UV-B exclusion had a common effect of enhancing the plant height, number of nodes and the area of the third trifoliate leaf in all the cultivars. The area of third trifoliate leaf showed enhancement, varied between a maximum of 53 % of control (var. JS-335) to a minimum of 3.4 % of control (Pusa-24) (Figure 2) at 5th node. Average number of nodes/plant was enhanced in all the eight cultivars by the exclusion of UV-B. The enhancement varied quantitatively from 37% (Hardee) and 28% in (Kalitur) to 4% in (PK-1029) (Figure 3). The plant height was enhanced by 20% to 31% of control in six of the cultivars (PK- 472, NRC- 7, Pusa -24, JS -335,JS -7105,PK -1029) (Figure 4).
All the eight cultivars showed a decline in the amount of UV absorbing substances synthesized by the exclusion of UV-B in third trifoliate leaf in all the eight cultivars (Figure 5). The decline varied quantitatively from 32% (JS-335) to 2.5% (PK-1029) (Figure 5).
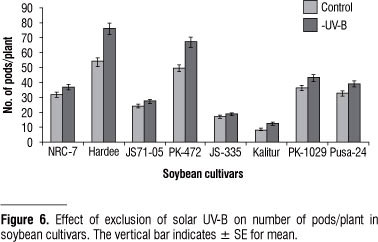
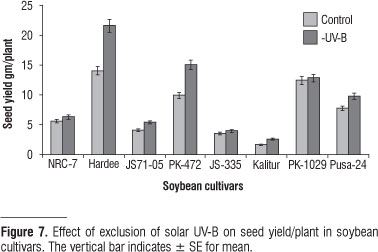
We found an enhancement in the number of pods/plant and seed yield after exclusion of solar UV-B in all the cultivars tested. Out of 8 cultivars, in 4 cultivars, there was a significant increase in the number of pods/plant from 35 to 43 % (Kalitur, Hardee and PK- 472); in the rest of the cultivars there was less than 20% enhancement (Figure 6). The seed yield was enhanced to the maximum in Hardee (53% of control) then in PK-472 (51%) and minimum in PK-1029 (4 % of control) (Figure 7).
Also, an inverse association between the enhancement in the height of the plant and increase in number of pods/plant was detected. The cultivars with higher than 25% enhancement in height showed less increase in the number of pods/plant with the exception of JS 71-05 (Figure 8).
There were intraspecific variations in the UV-B sensitivity amongst the soybean cultivars tested according to UV Sensitivity Index (UV-SI) by the exclusion of solar UV-B. The sensitivity index was numerically lowest in PK-472 (1.87) and highest for PK-1029 (2.64) rest of the cultivars exhibited UV-SI as JS-335(2.13), Hardee (2.24), Kalitur (2.29), JS 71-05 (2.29), Pusa-24(2.35) and NRC-7(2.38) (Figure 9).
DISCUSSION
Exclusion of UV-B had a general effect of enhancement in growth and yield of all the eight cultivars; although there were significant quantitative differences. In terms of yield cv. Kalitur, Hardee, JS71-05 and PK-472 showed improved performance both in terms of number of pods and seeds produced (Figure 6, 7). An enhancement in the seed yield by exclusion of solar UV-B implies a poor adaptation of the cultivar to the ambient solar UV-B. These four cultivars may give better yield only in those latitudes, which receive less of UV-B radiation. Enhancement in the biomass and yield by UV-B exclusion has been recorded earlier in India in Cymopsis, Vigna (Amudha et al., 2005) and soybean variety JS-71-05 (Varalakshmi et al., 2003; Guruprasad et al., 2007).
There was a significant inverse association between the enhancement in the vegetative growth of plant and the number of pods/plant amongst the cultivars tested. This data indicates differences in the carbon partition amongst the cultivars by way of exclusion. In general photosynthesis is enhanced in all the cultivars since the leaf expansion was enhanced by exclusion of UV-B (Figure 2). The additional carbon fixed is either directed towards the enhancement of vegetative growth or towards the production of pods and seeds. Those cultivars in which the partition of carbon is towards enhancement of growth show very less enhancement in the yield and their yield may not vary in latitudes with less of ambient UV-B radiation.
Zavala and Botto, (2002) also found that plant height was greater in radish plants grown under solar UV-B exclusion than those grown under ambient UV-B but on the other hand the percentage of dry matter partitioned to the tubers, the tuber fresh weight and the tuber diameter was consistently increased in plants grown under solar UV-B relative to those developed under UV-B exclusion. They came to the conclusion that in addition to source activity, strengthened sink demand may affect the biomass partitioned to tubers.
Sensitivity indices or response indices have established as useful indicators of plant sensitivity to UV-B radiation (enhanced or supplemental) (Saile-Mark and Tevini, 1997). The UV sensitivity index of all the cultivars tested was significantly less than 3, and it seems that all the tested cultivars are sensitive to ambient levels of UV-B. Cultivar PK-472 exhibited sensitivity index of 1.87 which seems to be highly affected by the ambient levels of UV-B and cultivar PK-1029 shows SI (2.64) is the least sensitive amongst the cultivars tested. These results indicate that ambient levels of UV-B have deleterious effect on the growth and yield characteristics in tested soybean cultivars. According to UV-SI, sensitivity of eight Indian soybean cultivars to ambient level of UV B (280-320 nm) radiation at Indore (22.40N) had the following descending order; PK-472>JS-335> Hardee>Kalitur>JS71-05>Pusa-24>NRC-7>PK-1029 (Figure 9).
A close link is also evident from the data between the partition of carbon to secondary and primary metabolism and its regulation by solar UV-B. Cultivars with lower UV-B SI, in which the amount of UAS is reduced to the maximum by UV-B exclusion (cv. JS-335, JS 71-05, Kalitur and PK-472, Figure 5); the weight of the seeds produced is enhanced to a high level (Figure 7). Whereas cultivars with higher UV-B SI in which the amount of UAS is reduced to lesser extent by exclusion of UV-B (cv. NRC-7, Pusa-24 and PK-1029) the weight of the seeds produced is less (Figure 7) A decrease in UAS was also found in radish and Cyamopsis by the exclusion of UV-B from solar radiation (Zavalla and Botto, 2002; Amudha et al., 2005). Intraspecific differences in the composition and concentration of flavonoids have been shown to determine the UV sensitivity amongst five cultivars of Cucumis sativus (Murali and Teramura, 1986) and in two cultivars of soybean (Murali et al., 1988). Thus the differences in the partition of carbon by UV-B exclusion may help in the selection of the cultivar. Exclusion studies will have significance under tropical environment where the plants receive higher amount of ambient UV-B. Exclusion studies would help in the assessment of the adaptability of a cultivar to ambient UV-B received and selection of the best-suited cultivar for given latitude. These intraspecific differences suggest that future attempts at breeding for increased tolerance to UV-B radiation might be successful.
Acknowledgements: The work received financial support from Department of Science Technology (WOS- A Scheme-SR/WOS-A/LS101/ 2009) to Kataria S. is thankfully acknowledged. Thanks to Dr. V.S.Bhatia, DRS, Indore (M.P.) for providing seeds of soybean cultivars.
Received: 12 March 2011
Accepted: 26 August 2011
- Amudha P, Jayakumar M, Kulandaivelu G (2005) Impacts of ambient solar UV (280-400 nm) radiation on three tropical legumes. J Plant Biol. 48:284-291.
- Bacwar MR, Moore FD, Burke MJ (1982) Effect of deletion and enhancement of ultraviolet-B (280-315) radiation on plant growth at 3000m elevation. Am. Soc. Hortic. Sci. 107:771-774.
- Caldwell MM (1968) Solar ultraviolet radiation as an ecological factor for alpine plants. Ecol. Monogr. 38: 243-268.
- Caldwell MM, Teramura AH, Tevini M (1989) The changing solar ultraviolet climate and the ecological consequences for higher plants. Trends Ecol. Evol.4: 363-367.
- Caldwell MM, Teramura AH, Tevini M, Bornman JF, Bjorn LO, Kulandaivellu G (1995) Effects of increased solar ultraviolet radiation on terrestrial plants. Ambio. 24:166-173.
- Carletti P, Masi A, Wonisch A, Grill D, Tausz M, Ferretti M (2003) Changes in antioxidant and pigment pool dimensions in UV-B irradiation maize seedlings. Environ. Exp. Bot. 50:149-157.
- Flint SD, Searles PS, Caldwell MM (2004) Field testing of UV biological spectral weighting functions for induction of UV - absorbing compounds in higher plants. Phtochem. Photobiol. 79, 399-403.
- Golaszewska Zuk K, Upadhyaya MK, Golaszewski J (2003) The effect of UV-B radiation on plant growth and development. Plant Soil Environ. 49 (3): 135-140.
- Guruprasad KN, Bhattacharjee S, Kataria S, Yadav S, Tiwari A, Baroniya S, Rajiv A, Mohanty P (2007) Growth enhancement of soybean (Glycine max) upon exclusion of UV-B and UV-A components of solar radiation: characterization of photosynthetic parameters in leaves. Photosyn. Res. 94:299-306.
- Jansen AK, Gaba V, Greenberg BM (1998) Higher plants and UV-B radiation: balancing damage, repair and acclimation. Trends Plant Sci. 3:131-135.
- Jayakumar M, Amudha P, Kulandaivelu G (2004) Effect of low doses of UV-A (320-400 nm) and UV-B (280-320 nm) radiation on photosynthetic activities in Phaseolus mungo L. J. Plant Biol. 47:105-110.
- Li Yuan, Lilian He, Yanqun Zu (2010) Intraspecific variation in sensitivity to ultraviolet-B radiation in endogenous hormones and photosynthetic characteristics of 10 wheat cultivars grown under field conditions. South African Journal of Botany 76:493-498.
- Madronich S, McKenzie RL, Caldwell MM, Bjorn LO (1995) Changes in ultraviolet radiation reaching the earth's surface. Ambio. 24:143-152.
- Mazza CA, Boccalandro HE, Giordano CV, Battista D, Sopel AL, Ballare CL (2000) Functional significance and induction by solar radiation of ultraviolet absorbing sunscreens in field grown soybean crops. Plant Physiol. 122:117-125.
- Mirecki RM, Teramura AH (1984) Effect of UV-B on soybean .V.The dependence of plant sensitivity on the photosynthetic photon flux density during and after leaf expansion. Plant Physiol. 74:475-480.
- Murali NS, Teramura AH (1986) Intraspecific differences in Cucumis sativus sensitivity to ultraviolet-B radiation. Physiol. Plant. 68:673-677.
- Murali NS, Teramura AH, Randall SK (1988) Response differences between two soybean cultivars with contrasting UV-B radiation sensitivities. Photochem. Photobiol. 48: 653-657.
- Pal M, Zaidi PH, Voleti SR, Raj A (2006) Solar UV-B exclusion effect on growth and photosynthetic characteristics of wheat and pea. J. New Seeds 8:19-34.
- Paul ND, Gwynn-Jones D (2003) Ecological roles of solar UV radiation: towards an integrated Approach. Trends Ecol. Evol. 18:48-55.
- Rathore D, Agrawal SB, Singh A (2003) Influence of supplemental UV-B radiation and mineral nutrients on biomass, pigments and yield of two cultivars of wheat. Int J Biotron 32:1-15.
- Saile-Mark M, Tevini M (1997) Effects of solar UV-B radiation on growth, flowering and yield of central and southern European bush bean cultivars (Phaseolus vulgaris L.). Plant Ecol. 128:115-125.
- Smith JL, Burritt DJ, Bannister P, (2000) Shoot dry weight, chlorophyll and UV-B absorbing compounds as indicators of a plant's sensitivity to UV-B radiation. Ann. Bot. 86:1057-1066.
- Tevini M (2000) UV-B effects on plant. In: Agrawal SB, Agrawal M (eds), Environmental Pollution and Plant Responses, pp. 83-97. Lewis Publishers, Boca Raton, USA.
- Tezuka T, Hotta T, Wanatabe I (1993) Growth promotion of tomato and radish plants by solar UV radiation reaching the earth's surface. Photochem. Photobiol.19:61-66.
- Varalakshmi D, Lakshmi N, Guruprasad KN (2003) Physiological changes in soybean Cv. JS 71-05 after the exclusion of UV-A and UV-B from the solar radiation. Indian J. Plant Physiology (Special Issue):602-606.
- Yanqun Z, Yuan Li, Haiyan C., Jianjun C (2003) Intraspecific differences in physiological response of 20 soybean cultivars to enhanced ultraviolet-B radiation under field conditions. Environ. Exp. Bot. 50:87-97.
- Yanqun Z, Yuan Li, Jianjun JC, Chen HY (2004) Intraspecific responses in grain quality of 10 wheat cultivars to enhanced UV-B radiation under field conditions. J. Photochem. Photobiol. B Biol. 74:95-100.
- Zavalla JA Botto, JF (2002) Impact of solar UV-B radiation on seedling emergence, chlorophyll fluorescence and growth and yield of radish (Raphanus sativus). Funct. Plant Biology 29:797-804.
Publication Dates
-
Publication in this collection
20 Jan 2012 -
Date of issue
2011
History
-
Received
12 Mar 2011 -
Accepted
26 Aug 2011