Abstract
Objective
This study sought to analyze the gene expression of Candida albicans in sound root surface and root caries lesions, exploring its role in root caries pathogenesis.
Methodology
The differential gene expression of C. albicans and the specific genes related to cariogenic traits were studied in association with samples of biofilm collected from exposed sound root surface (SRS, n=10) and from biofilm and carious dentin of active root carious lesions (RC, n=9). The total microbial RNA was extracted, and the cDNA libraries were prepared and sequenced on the Illumina Hi-Seq2500. Unique reads were mapped to 163 oral microbial reference genomes including two chromosomes of C. albicans SC5314 (14,217 genes). The putative presence of C. albicans was estimated (sum of reads/total number of genes≥1) in each sample. Count data were normalized (using the DESeq method package) to analyze differential gene expression (using the DESeq2R package) applying the Benjamini-Hochberg correction (FDR<0.05).
Results
Two genes (CaO19.610, FDR=0.009; CaO19.2506, FDR=0.018) were up-regulated on SRS, and their functions are related to biofilm formation. Seven genes ( UTP20 , FDR=0.018; ITR1 , FDR=0.036; DHN6 , FDR=0.046; CaO19.7197 , FDR=0.046; CaO19.7838 , FDR=0.046; STT4 , FDR=0.046; GUT1 , FDR=0.046) were up-regulated on RC and their functions are related to metabolic activity, sugar transport, stress tolerance, invasion and pH regulation. The use of alternative carbon sources, including lactate, and the ability to form hypha may be a unique trait of C. albicans influencing biofilm virulence.
Conclusions
C. albicans is metabolically active in SRS and RC biofilm, with different roles in health and disease.
Sequence analysis; RNA; Candida albicans; Root caries; Transcriptome
Introduction
The bacterial biofilm associated with root caries lesions must harbor microorganisms that can produce acid from carbohydrates (acidogenicity) and must be able to growth in a low-pH environment (aciduricity). 11 - Brailsford SR, Shah B, Simons D, Gilbert S, Clark D, Ines I, et al. The predominant aciduric microflora of root-caries lesions. J Dent Res. 2001;80(9):1828-33. Diverse bacteria are prevalent and involved in the etiology of root caries, albeit to date, little has been explored regarding other microorganisms domains, such as archea, fungi and virus, and their role in biofilms. Previous studies demonstrated that Streptococcus mutans , Lactobacillus species (spp.), and Veillonella spp., as well as C. albicans, are present in major proportion in root caries than in sound root surface. 22 - Marchant S, Brailsford SR, Twomey AC, Roberts GJ, Beighton D. The predominant microflora of nursing caries lesions. Caries Res. 2001;35(6):397-406. doi: 10.1159/000047482
https://doi.org/10.1159/000047482...
Actinomyces spp., Veillonella spp., Streptococcus spp., Bifidobacterium spp., Rothia, Enterococcus , Staphylococcus spp., Capnocytophaga spp., Prevotella spp. and Candida spp., were also cultivated from root caries. 11 - Brailsford SR, Shah B, Simons D, Gilbert S, Clark D, Ines I, et al. The predominant aciduric microflora of root-caries lesions. J Dent Res. 2001;80(9):1828-33. , 33 - Shen S, Samaranayake LP, Yip HK, Dyson JE. Bacterial and yeast flora of root surface caries in elderly, ethnic Chinese. Oral Microbiol. 2002;8(4):207-17. doi: 10.1034/j.1601-0825.2002.01796.x
https://doi.org/10.1034/j.1601-0825.2002...
, 44 - Mantzourani M, Fenlon M, Beighton D. Association between Bifidobacteriaceae and the clinical severity of root caries lesions. Oral Microbiol Immunol. 2009;24(1):32-7. doi: 10.1111/j.1399-302X.2008.00470.x
https://doi.org/10.1111/j.1399-302X.2008...
Candida species has been associated with dental caries, especially with early childhood caries and root caries. 55 - Carvalho FG, Silva DS, Hebling J, Spolidorio LC, Spolidorio DM. Presence of mutans streptococci and Candida spp. in dental plaque/dentin of carious teeth and early childhood caries. Arc Oral Biol. 2006;51(11):1024-8. doi: 10.1016/j.archoralbio.2006.06.001
https://doi.org/10.1016/j.archoralbio.20...
A strong association was found between the prevalence of C. albicans and dental caries. 66 - Beighton D, Brailsford S, Samaranayake LP, Brown JP, Ping FX, Grant-Mills D, et al. A multi-country comparison of caries-associated microflora in demographically diverse children. Community Dent Health. 2004;21(1):96-101. Several authors showed that the proportion of Candida species was higher in individuals with caries than in individuals without caries. Furthermore, C. albicans is an important colonizer of carious lesions and has been found frequently in dentin caries lesions rather than in biofilm or saliva. 44 - Mantzourani M, Fenlon M, Beighton D. Association between Bifidobacteriaceae and the clinical severity of root caries lesions. Oral Microbiol Immunol. 2009;24(1):32-7. doi: 10.1111/j.1399-302X.2008.00470.x
https://doi.org/10.1111/j.1399-302X.2008...
Lower salivary flow rate, a common occurrence in older adults, is one of the factors that promote favorable conditions for a presence of C. albicans in these sites. 77 - Do T, Damé-Teixeira N, Naginyte M, Marsch PD Root surface biofilms and caries. Monogr Oral Sci. 2017;26:26-34. doi: 10.1159/000479304
https://doi.org/10.1159/000479304...
However, it is still unknown whether the yeast acts as caries pathogen or plays a role as a commensal microbe. C. albicans possess some important properties that can characterize it as an important root caries pathogen. It is capable of adhering to saliva-coated hydroxyapatite and possesses strong adherence to collagen. 88 - Makihira S, Nikawa H, Tamagami M, Hamada T, Samaranayake LP. Differences in Candida albicans adhesion to intact and denatured type I collagen in vitro. Oral Microbiol Immunol. 2002;17(2):129-31. doi: 10.1046/j.0902-0055.2001.00094.x
https://doi.org/10.1046/j.0902-0055.2001...
It is as acid tolerant and acidogenic as S. mutans and Lactobacilli , which are both well-established cariogenic pathogens. 99 - Aas JA, Griffen AL, Dardis SR, Lee AM, Olsen I, Dewhirst FE, et al. Bacteria of dental caries in primary and permanent teeth in children and young adults. J Clin Microbiol. 2008;46(4):1407-17. doi: 10.1128/JCM.01410-07
https://doi.org/10.1128/JCM.01410-07...
To determine the role of C. albicans in root caries, a high-throughput sequencing of mRNA (RNA-Seq) was applied in clinical biofilms samples from two distinct conditions: sound root-surface biofilms and root carious lesions biofilms. This technique may be helpful to investigate Candida’s role in a carious biofilm.
Methodology
This study is part of the project “metatranscriptome of root caries”. 1010 - Damé-Teixeira N, Parolo CC, Maltz M, Tugnait A, Devine D, Do T. Actinomyces spp. gene expression in root caries lesions. J Oral Microbiol. 2016;6(8):32383. doi: 10.3402/jom.v8.32383
https://doi.org/10.3402/jom.v8.32383...
Briefly, volunteers to this study were divided into two groups: sound exposed root surface group (SRS; n=10) and root caries group (RC; n=30). Participants were allocated to the SRS group (n=10) if they had an exposed root surface on at least one tooth and no root caries lesions. Dental biofilms were collected with sterilized Gracey curette from all available exposed root surfaces. The number of exposed root surfaces varied among individuals. Participants recruited to the root caries (RC) group ( n =30) had one primary cavitated root lesion in need of restorative treatment. All lesions showed characteristics of present activity (soft and yellow dentin). Biofilm and carious dentin samples (of soft and infected tissue) were collected from patients during the restorative treatment. All participants were asked to refrain from tooth brushing for at least 12 hours prior to the sampling, to allow for dental biofilm accumulation, and were also asked to refrain from eating and drinking for at least 1 hour prior to the sampling. After collection, biofilm and carious dentin were immediately placed in 1 mL of RNA protect reagent (Qiagen, Hilden, North Rhine-Westphalia, Germany). The total RNA was extracted using the UltraClean ® Microbial RNA Isolation (Mo-bio, San Diego, Califórnia, USA) with on-column DNase digestion (Qiagen, Hilden, North Rhine-Westphalia, Germany). Samples with total RNA concentration <30 ng/RNA were pooled, leading to a final sample size of 10 SRS and 9 RC. The Ribo-Zero™ Meta-Bacteria Kit (Illumina, Madison, Wisconsin, USA) was used for mRNA enrichment and Illumina ® TruSeq™ library prep protocols (Illumina, San Diego, Califórnia, USA) were used to library preparation and sequencing was performed with Illumina HiSeq2500 (Illumina, San Diego, Califórnia, USA). RNA sequencing data are available in the National Center for Biotechnology Information (NCBI) Sequence Read Archive, under the accession numbers SRS779973 and SRS796739. FASTQ files were obtained for each sample and imported into the CLC Genomics Workbench 7.5.1 software (CLC bio, Aarhus, Denmark) for mapping against 163 oral microbial genomes. 1010 - Damé-Teixeira N, Parolo CC, Maltz M, Tugnait A, Devine D, Do T. Actinomyces spp. gene expression in root caries lesions. J Oral Microbiol. 2016;6(8):32383. doi: 10.3402/jom.v8.32383
https://doi.org/10.3402/jom.v8.32383...
The number of sequence reads that have been assigned to each gene is considered as the read count data.
Candida albicans genome and data analysis
The C. albicans SC5314 was the genome selected for this study. This strain was chosen for being largely studied and its genome has been fully sequenced as well. After mapping, a count table was generated containing the read count for 14,217 oral C. albicans SC5314 genes.
The putative presence of the organism in the sample was estimated by the sum of reads assigned to C. albi cans divided by the total number of genes for each sample. Samples with ≥14,217 reads were considered as valid; then samples with less than 30% of genes with at least one read were excluded from the analysis.
The number of reads and the relative median expression (RME) (25 th -75 th ) level for genes were estimated for each of the sample groups, as previously described. 1111 - Do T, Sheehy EC, Mulli T, Hughes F, Beighton D. Transcriptomic analysis of three Veillonella spp. present in carious dentin and in the saliva of caries-free individuals. Front Cell Infect Microbiol. 2015;5:25. doi: 10.3389/fcimb.2015.00025
https://doi.org/10.3389/fcimb.2015.00025...
Then, the RME was ranked to observe the most highly expressed transcripts in RC and SRS samples. To draw a profile of gene expression, the median of RME of transcripts in SRS and RC conditions were considered low expression RME between 0-10, medium 11-100, and high above 100 (percentile 10 of RME distribution). RME was calculated from the median values of normalized read counts using DE-Seq algorithm. Genes related to C. albicans virulence factors were analyzed: invasion, biofilm formation and co-aggregation, adherence and damage, morphogenesis, acid production, acid tolerance and stress response.
All RME medians for SRS and RC were ranked and all genes with median RME values ≥100 per group were analyzed for an overview of the most prevalent genes.
Differential gene expression was inferred between sample groups by applying the R package DESeq2. 1212 - Love M, Anders S, Huber W. Differential analysis of count data – the DESeq2 package [Internet]. 2016 Nov 30. [cited 2019 Oct 1]. Available from: https://www.genomatix.de/online_help/help_regionminer/DESeq2.pdf
https://www.genomatix.de/online_help/hel...
The cut-off for designating a gene as being differentially expressed was a change in transcript levels of at least 2-fold change (Log2FoldChange>1) and false discovery rate (FDR) <0.05 (padj value<0.05, Benjamini & Hochberg). Functions and putative pathogenicity in root caries of genes up-regulated in SRS and RC were analyzed.
Regarding to ethics considerations, this study was approved by the Federal University of Rio Grande do Sul research ethics committee (process n° 427.168) and by the research ethics committee of the National Research Ethics Service Committee Yorkshire & The Humber – Leeds West (protocol no. 2012002DD). All volunteers signed an informed consent form and received clinical dental assistance.
Results
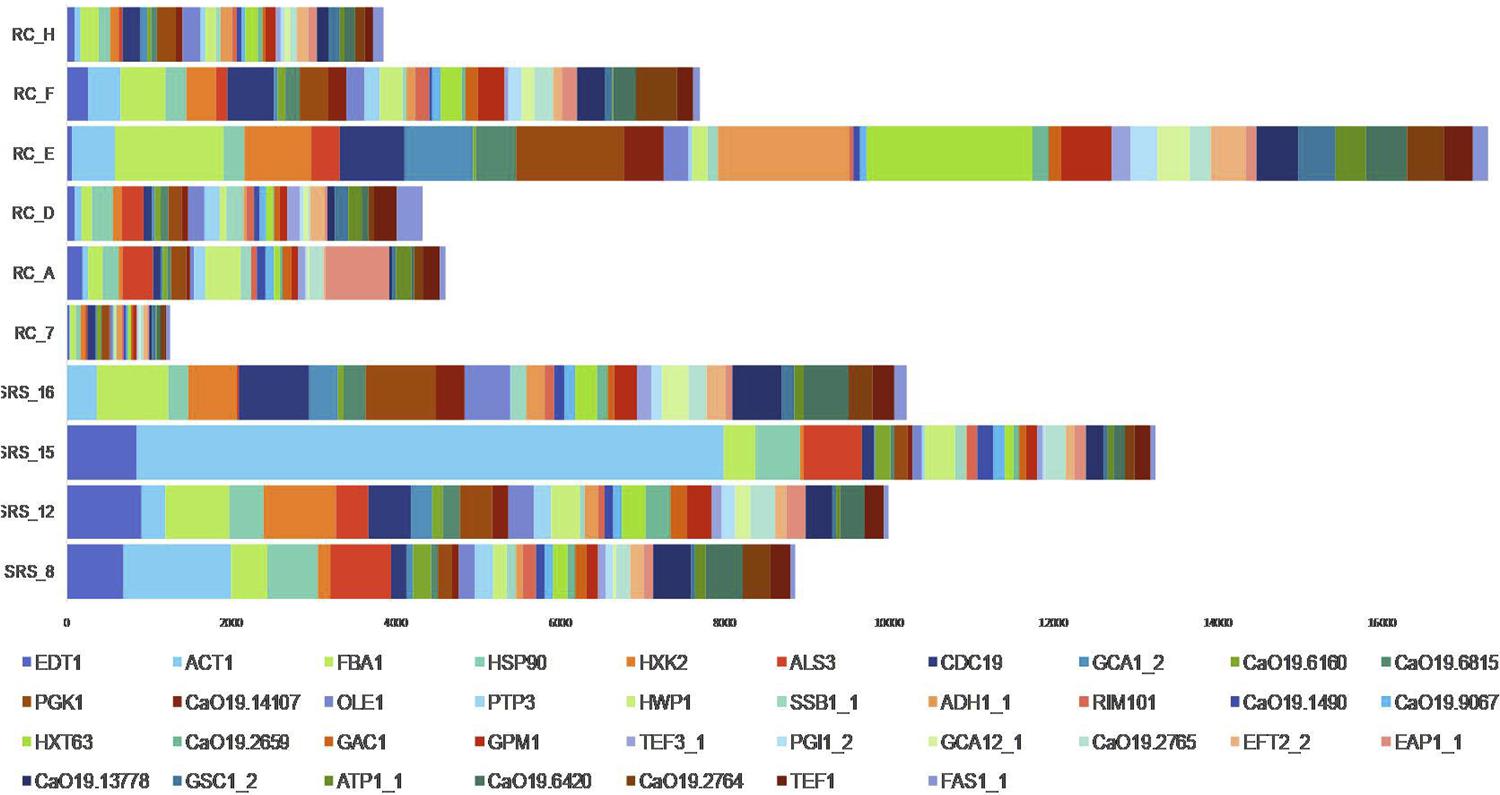
According to the cut-off point chosen to determine the putative presence of a mapped organism in each sample, C. albicans was present in n=4 biofilms from SRS and in n=6 biofilm from RC, as shown in Figure 1 . Table 1 shows that the number of reads distribution in sound and disease samples were equal (p=0.522).
Relative median expression (RME; log10) of genes in the Sound Root Surfaces (SRS; n=4) and Root Caries (RC; n=6) samples. RME was calculated from the median values of normalized read counts. The top median RME values for SRS and RC were selected and sorted, and indicate the most expressed genes by C. albicans SC5413
Gene expression per sample
Figure 1 shows an overview of the most prevalent genes in C. albicans biofilm with and without caries. A total of 37 genes with median of RME>100 were analyzed (34 in SRS and 20 in RC). A total of 17 genes have RME>100 in both health and disease conditions for all the samples ( FBA1, HSP90, HXK2, ALS3, CDC19, PGK1, OLE1, HWP1, HXT63, GPM1, CaO19.2765, EFT2_2, EAP1_1, CaO19.13778, CaO19.6420, CaO19.2764 and TEF1 ), wheras 17 genes were expressed only in SRS ( EDT1, ACT1, GCA1_2, CaO19.6160, CaO19.6815, CaO19.14107, PTP3, SSB1_1, ADH1_1, RIM101, CaO19.1490, CaO19.9067, CaO19.2659, GAC1, TEF3_1, PGI1_2 and GCA12_1 ) and just 3 genes were expressed only in RC conditions ( GSC1_2, ATP1_1 and FAS1_1 ), as shown in Table 2 .
Relative median expression (RME) and percentiles (25 th -75 th ) of genes related to virulence factors in Candida albicans in the Sound Root Surfaces (SRS; n=4) and Root Caries (RC; n=6) samples
Expression of genes related to possible cariogenic traits
C. albicans genes associated with possible virulence factors (RME and percentiles of these genes) were evaluated in both conditions ( Table 2 ). We found transcript of 51 out of 67 genes related to virulence traits that are presented in the literature as important factors. None of these genes had significant differential expression.
Differential expression analysis (DE)
The DE analysis has shown the overexpressed genes in root biofilms with and without caries ( Figure 2 ). The up-regulated genes in SRS group were CaO19.610 and CaO19.2506 . The CaO19.610 (FDR=0.009) codes for a potential DNA binding regulator of filamentous growth. The CaO19.2506 (FDR=0.018) codes for a hypothetical protein with a very weak similarity to Streptococcal proline-rich surface protein PspC.
Differential expression (Log2FoldChange) of genes up-regulated in sound root surface (SRS; negative values, green bars) and up-regulated in root caries (RC, positive values, red bars) calculated using DESeq2 algorithms. FDR<0.05. GUT1= potential glycerol kinase; STT4= hypothetical protein phosphatidylinositol-4-kinase; CaO19.7838= flocullin-like protein serine-rich; CaO19.7197= hypothetical protein; DHN6= dehydrin hypothetical protein; ITR1= potential Myo-inositol transporter; UTP20= potential U3 small nucleolar RNAs protein; CaO19.2506= hypothetical protein with a very weak similarity to Streptoccal proline-rich surface protein PspC; CaO19.610= potential DNA binding regulator of filamentous growth
The up-regulated genes in RC group were UTP20, ITR1, DHN6, CaO19.7197, CaO19.7838, STT4, and GUT1 . The UTP20 (FDR=0.018) codes for a potential U3 small nucleolar RNAs (snoRNA) protein. The ITR1 (FDR=0.036) codes for a potential active sugar transporter, potential Myo-inositol transporter, similar to S. cerevisiae ITR1 (YDR497C). The DHN6 (FDR=0.046) codes for a dehydrin hypothetical protein. The CaO19.7197 (FDR=0.046) codes for a hypothetical protein similar to S. cerevisiae YLR002C, with unknown function. The CaO19.7838 (FDR=0.046) codes for a flocculin-like protein serine-rich, repetitive ORF similar to S. cerevisiae MUC1 (YIR019C) cell surface flocculin. The STT4 (FDR=0.046) codes for a hypothetical protein phosphatidylinositol-4-kinase. The GUT1 (FDR=0.046) codes for a potential glycerol kinase Gut1p, likely carbohydrate kinase similar to S. cerevisiae GUT1 (YHL032C) glycerol kinase.
Discussion
Possible virulence traits of Candida spp. were related to several survival strategies such as the capacity to exploit and invade the host tissues, forming biofilms and co-aggregate to various microorganisms, switching form, producing acids and reacting to stress. C. albicans is metabolically active in biofilm of SRS and RC, presenting different roles in health and disease. Some genes were expressed in both conditions, which seem to be relevant to C. albicans survival to these sites. Genes overexpressed in SRS were involved in biofilm formation, while genes overexpressed in RC were involved in survival strategies that could be related to cariogenicity.
Two genes were up-regulated in SRS biofilms. The CaO19.610 codes for a potential DNA binding regulator of filamentous growth. This gene is a version of C. albicans efg1 with altered C terminus. EFG1 protein is a key transcriptional regulator in C. albicans and controls various aspects of morphogenesis and metabolism 1313 - Noffz CS, Liedschulte V, Lengeler K, Ernst JF. Functional mapping of the Candida albicans Efg1 regulator. Eukaryotic Cell. 2008;7(5):881-93. doi: doi: 10.1128/EC.00033-08
https://doi.org/10.1128/EC.00033-08...
, being required for the true hyphae growth, biofilm formation, cell adhesion and filamentous growth in C. albicans . 1414 - Gancedo JM. Control of pseudohyphae formation in Saccharomyces cerevisiae. FEMS Microbiology Reviews. 2001;25(1):107-23. doi: 10.1111/j.1574-6976.2001.tb00573.x
https://doi.org/10.1111/j.1574-6976.2001...
Efg1 gene confers to C. albicans the capacity of transition from commensal microorganism to opportunistic pathogen status. 1515 - Pierce VJ, Kumamoto AC. Variation in Candida albicans EFG1 expression enables host-dependent changes in colonizing fungal populations. MBio. 2012;3(4):e00117-12. doi: 10.1128/mBio.00117-12
https://doi.org/10.1128/mBio.00117-12...
In an in vitro experiment, efg1 had significantly higher gene expression at initial biofilm formation stage. 1616 - Barros PP, Rossoni RD, Camargo Ribeiro F, Junqueira JC, Jorge AO. Temporal profile of biofilm formation, gene expression and virulence analysis in Candida albicans strains. Mycopathologia. 2017;182(3-4):285-95. doi: 10.1007/s11046-016-0088-2
https://doi.org/10.1007/s11046-016-0088-...
Other studies showed that EFG1 is essential for the formation of a mature and stable biofilm that is resistant to antifungal therapy and to immune system, allowing the colonization of the root site. 1717 - Finkel JS, Mitchell AP. Genetic control of Candida albicans biofilm development. Nat Rev Microbiol. 2011;9(2):109-18. doi: 10.1038/nrmicro2475
https://doi.org/10.1038/nrmicro2475...
, 1818 - Nobile CJ, Fox EP, Nett JE, Sorrells TR, Mitrovich QM, Hernday AD, et al. A recently evolved transcriptional network controls biofilm development in Candida albicans. Cell. 2012;148(1-2):126-38. The CaO19.2506 codes for a hypothetical protein with a very weak similarity to streptococcal proline-rich surface protein PspC. In S. pneumoniae , PspC has a well-established importance in adherence and colonization. 1919 - Rosenow CP, Ryan JN, Weiser S, Johnson P, Fontan A, Ortqvist HR, et al. Contribution of novel choline-binding proteins to adherence, colonization, and immunogenicity of Streptococcus pneumoniae. Mol Microbiol. 1997;25(5):819-29. 10.1111/j.1365-2958.1997.mmi494.x
https://doi.org/10.1111/j.1365-2958.1997...
The possible function of CaO19.2506 is related to adhesion and coding for a membrane adhesin. Both filamentous growth and cell wall adhesion are important in biofilm formation and are required for the establishment of C. albicans in root surfaces biofilm. These characteristics could explain why C. albicans has been largely observed colonizing sound root surface. 55 - Carvalho FG, Silva DS, Hebling J, Spolidorio LC, Spolidorio DM. Presence of mutans streptococci and Candida spp. in dental plaque/dentin of carious teeth and early childhood caries. Arc Oral Biol. 2006;51(11):1024-8. doi: 10.1016/j.archoralbio.2006.06.001
https://doi.org/10.1016/j.archoralbio.20...
The role of C. albicans in root caries could be potentially more complex. Seven genes were up-regulated in root caries conditions expressing different functions. The CaO19.7197 codes for a hypothetical protein similar to S. cerevisiae YLR002C, with unknown function. Several hypothetical proteins and genes with uncharacterized function were identified in this study, highlighting the importance of more studies related to C. albicans transcriptome. The DHN6 codes for a dehydrin hypothetical protein, related to stress tolerance in plants. These proteins can be induced in vegetative tissues by different stress factors that cause cell dehydration (i.e., drought, salinity, cold, heat, low temperature, etc). 2020 - Hanin M, Brini F, Ebel C, Toda Y, Takeda S, Masmoudi K. Plant dehydrins and stress tolerance: versatile proteins for complex mechanisms. Plant Signal Behav. 2011;6(10):1503-9. doi: 10.4161/psb.6.10.17088
https://doi.org/10.4161/psb.6.10.17088...
, 2121 - Drira M, Saibi W, Amara I, Masmoudi K, Hanin M, Brini F. Wheat dehydrin K-segments ensure bacterial stress tolerance, antiaggregation and antimicrobial effects. Appl Biochem Biotechnol. 2015;175(7):3310-21. doi: 10.1007/s12010-015-1502-9
https://doi.org/10.1007/s12010-015-1502-...
The STT4 codes for a hypothetical protein phosphatidylinositol-4-kinase. The gene STT4 is essential for viability and plays an important role in the phosphatidylinositol-mediated signal transduction pathway required for cell wall integrity. 2222 - Pramanik A, Garcia E, Ospina R, Powell M, Martinez M, Alejo W, et al. Cloning, characterization and identification of the gene encoding phosphatidylinositol 4-kinase. Cell Mol Biol. 1997;43(7):1007-18. Therefore, the up-regulated genes DHN6 and STT4 could be related to the ability to survive in an extreme environment with several stress factors (low pH, carbohydrate viability, for example) such as the ones found in root cavitated caries lesions. The UTP20 codes for a potential U3 small nucleolar RNAs (snoRNA) protein. UTP20 has been reported as a component of U3 snoRNA protein complex and has been implicated in 18S rRNA processing, being essential for 18 rRNA function. 2323 - Wang Y, Liu J, Zhao H, Lü W, Zhao J, Yang L, et al. Human 1A6/DRIM, the homolog of yeast Utp20, functions in the 18S rRNA processing. Biochim Biophys Acta. 2007;1773(6):863-8. doi: 10.1016/j.bbamcr.2007.04.002
https://doi.org/10.1016/j.bbamcr.2007.04...
, 2424 - Dez C, Dlakić M, Tollervey D. Roles of the HEAT repeat proteins Utp10 and Utp20 in 40S ribosome maturation. RNA. 2007;13(9):1516-27. doi: 10.1261/rna.609807
https://doi.org/10.1261/rna.609807...
The ITR1 codes for a potential active sugar transporter, potential Myo-inositol transporter, similar to S. cerevisiae ITR1 (YDR497C). Myo-inositol is an essential substrate for C. albicans , and it can be used as carbon source. For its survival, C. albicans must be able to synthesize the essential metabolite inositol or acquire it from the host. C. albican s could not transport inositol and become nonviable in the absence of ITR1 . 2525 - Chen YL, Kauffman S, Reynolds TB. Candida albicans uses multiple mechanisms to acquire the essential metabolite inositol during infection. Infect Immun. 2008;76(6):2793-801. doi: 10.1128/IAI.01514-07
https://doi.org/10.1128/IAI.01514-07...
The CaO19.7838 codes for flocculin-like protein serine-rich, repetitive ORF similar to S. cerevisiae MUC1 (YIR019C) cell surface flocculin. MUC1 encodes cell-surface flocculin and it is required for pseudohyphal and invasive growth of C. albicans.
1414 - Gancedo JM. Control of pseudohyphae formation in Saccharomyces cerevisiae. FEMS Microbiology Reviews. 2001;25(1):107-23. doi: 10.1111/j.1574-6976.2001.tb00573.x
https://doi.org/10.1111/j.1574-6976.2001...
The up-regulation of this invasive growth gene shows the importance of this virulence trait for the colonization/penetration of C. albicans in the carious dentin. The GUT1 codes for a potential glycerol kinase Gut1p, likely carbohydrate kinase similar to S. cerevisiae GUT1 (YHL032C) glycerol kinase (NCBI). In Saccharomyces cerevisiae , glycerol utilization is mediated by two enzymes, glycerol kinase (Gut1p) and mitochondrial glycerol-3-phosphate dehydrogenase (Gut2p). The carbon source regulation of GUT1 depends on carbon source availability. The promoter activity of GUT1 was lower during growth on glucose and highest on the non-fermentable carbon sources, glycerol, ethanol, lactate, acetate and oleic acid. 2626 - Grauslund M, Lopes JM, Rønnow B. Expression of GUT1, which encodes glycerol kinase in Saccharomyces cerevisiae, is controlled by the positive regulators Adr1p, Ino2p and Ino4p and the negative regulator Opi1p in a carbon source-dependent fashion. Nucleic Acids Res. 1999;27(22):4391-8. doi: 10.1093/nar/27.22.4391
https://doi.org/10.1093/nar/27.22.4391...
UTP20, ITR1, CaO19.7838 , and GUT1 are genes related to C. albicans metabolism associated with caries progression to a cavitated status.
The overexpressed genes in RC were related to sugar transport (ITR1 – Myo-inositol) and to carbon source regulation ( GUT1 – Glycerol kinase), that were related to the use of alternative carbon sources ( Figure 2 ). The use of lactate by C. albicans could be related to the pH regulation (neutralization) in biofilm, which is important for the microbiome survival. 2727 - Danhof HA, Vylkova S, Vesely EM, Ford AE, Gozalez-Garay M, Lorenz M. Robust extracellular pH modulation by Candida albicans during growth in carboxylic acids. mBio. 2016;7(6):e01646-16. doi: 10.1128/mBio.01646-16
https://doi.org/10.1128/mBio.01646-16...
Furthermore, this neutralization of the medium could explain the CaO19.7838 overexpression, a gene related to hyphal growth since hyphal formation is stimulated by neutral pH. 2828 - Sudbery PE. Growth of Candida albicans hyphae. Nat Rev Microbiol. 2011;9(10):737-48. doi: 10.1038/nrmicro2636
https://doi.org/10.1038/nrmicro2636...
Morphogenesis is a special virulence trait of C. albicans, and hyphal form is related to pathogenesis, being more invasive and contributing to host tissue damage 2929 - Calderone RA, Fonzi WA. Virulence factors of Candida albicans. Trends Microbiol. 2001;9(7):327-35. doi: 10.1016/s0966-842x(01)02094-7
https://doi.org/10.1016/s0966-842x(01)02...
, 3030 - Höfs S, Mogavero S, Hube B. Interaction of Candida albicans with host cells: virulence factors, host defense, escape strategies, and the microbiota. J Microbiol. 2016;54(3):149-69. doi: 10.1007/s12275-016-5514-0
https://doi.org/10.1007/s12275-016-5514-...
, as well as contributing to the active cavitation of RC lesions. Besides the stimulation of hyphal growth, changes in carbon source has a significant impact on the C. albicans virulence, resulting in an increased resistance to stresses, adherence, biofilm formation, drug resistance, and immune recognition when compared with glucose-grown cells. 3131 - Ene IV, Heilmann CJ, Sorgo AG, Walker LA, de Koster CG, Munro CA, et al. Carbon source-induced reprogramming of the cell wall proteome and secretome modulates the adherence and drug resistance of the fungal pathogen Candida albicans. Proteomics. 2012;12(21):3164-79. doi: 10.1002/pmic.201200228
https://doi.org/10.1002/pmic.201200228...
Although a cariogenic environment is related to low pH conditions, the excessive production of acids could affect the biofilm metabolism. For the cariogenic biofilm survival, it is important to have a microorganism that main the viability of the biofilm, thus preventing excessive acidification even in a carious habitat.
Conclusions
Our data shows that Candida albicans SC5314 have an active metabolism in biofilm of SRS and biofilm of carious dentin of RC as well. The differential expression analysis shows that, in healthy individuals, the up-regulated genes were related to metabolic activity, sugar transport, stress tolerance, invasion and pH regulation. C. albicans may have a role in root caries progression.
Acknowledgements
The authors would like to thank Brazilian National Counsel of Technological and Scientific Development (CNPQ) (process no. 482504/2013-7), Coordination for the Improvement of Higher Level Education (CAPES) (process no 18097-12-0), Rio Grande do Sul State Foundation for Research Support (FAPERGS) (process no. 001/2013 PQG) and Leeds Teaching Hospitals Charitable Foundation (R&D/PP/12011), Dunhill Medical Trust (R245/0212). This study is derived from a thesis submitted by the first author to the Dental School of Federal University of Rio Grande do Sul as partial fulfilment of the requirements for a master’s degree in Clinical Dentistry – Cariology/Operative Dentistry. Partial results were presented at the 65thORCA Congress, held in Copenhagen (Denmark), 2018.
References
-
1- Brailsford SR, Shah B, Simons D, Gilbert S, Clark D, Ines I, et al. The predominant aciduric microflora of root-caries lesions. J Dent Res. 2001;80(9):1828-33.
-
2- Marchant S, Brailsford SR, Twomey AC, Roberts GJ, Beighton D. The predominant microflora of nursing caries lesions. Caries Res. 2001;35(6):397-406. doi: 10.1159/000047482
» https://doi.org/10.1159/000047482 -
3- Shen S, Samaranayake LP, Yip HK, Dyson JE. Bacterial and yeast flora of root surface caries in elderly, ethnic Chinese. Oral Microbiol. 2002;8(4):207-17. doi: 10.1034/j.1601-0825.2002.01796.x
» https://doi.org/10.1034/j.1601-0825.2002.01796.x -
4- Mantzourani M, Fenlon M, Beighton D. Association between Bifidobacteriaceae and the clinical severity of root caries lesions. Oral Microbiol Immunol. 2009;24(1):32-7. doi: 10.1111/j.1399-302X.2008.00470.x
» https://doi.org/10.1111/j.1399-302X.2008.00470.x -
5- Carvalho FG, Silva DS, Hebling J, Spolidorio LC, Spolidorio DM. Presence of mutans streptococci and Candida spp. in dental plaque/dentin of carious teeth and early childhood caries. Arc Oral Biol. 2006;51(11):1024-8. doi: 10.1016/j.archoralbio.2006.06.001
» https://doi.org/10.1016/j.archoralbio.2006.06.001 -
6- Beighton D, Brailsford S, Samaranayake LP, Brown JP, Ping FX, Grant-Mills D, et al. A multi-country comparison of caries-associated microflora in demographically diverse children. Community Dent Health. 2004;21(1):96-101.
-
7- Do T, Damé-Teixeira N, Naginyte M, Marsch PD Root surface biofilms and caries. Monogr Oral Sci. 2017;26:26-34. doi: 10.1159/000479304
» https://doi.org/10.1159/000479304 -
8- Makihira S, Nikawa H, Tamagami M, Hamada T, Samaranayake LP. Differences in Candida albicans adhesion to intact and denatured type I collagen in vitro. Oral Microbiol Immunol. 2002;17(2):129-31. doi: 10.1046/j.0902-0055.2001.00094.x
» https://doi.org/10.1046/j.0902-0055.2001.00094.x -
9- Aas JA, Griffen AL, Dardis SR, Lee AM, Olsen I, Dewhirst FE, et al. Bacteria of dental caries in primary and permanent teeth in children and young adults. J Clin Microbiol. 2008;46(4):1407-17. doi: 10.1128/JCM.01410-07
» https://doi.org/10.1128/JCM.01410-07 -
10- Damé-Teixeira N, Parolo CC, Maltz M, Tugnait A, Devine D, Do T. Actinomyces spp. gene expression in root caries lesions. J Oral Microbiol. 2016;6(8):32383. doi: 10.3402/jom.v8.32383
» https://doi.org/10.3402/jom.v8.32383 -
11- Do T, Sheehy EC, Mulli T, Hughes F, Beighton D. Transcriptomic analysis of three Veillonella spp. present in carious dentin and in the saliva of caries-free individuals. Front Cell Infect Microbiol. 2015;5:25. doi: 10.3389/fcimb.2015.00025
» https://doi.org/10.3389/fcimb.2015.00025 -
12- Love M, Anders S, Huber W. Differential analysis of count data – the DESeq2 package [Internet]. 2016 Nov 30. [cited 2019 Oct 1]. Available from: https://www.genomatix.de/online_help/help_regionminer/DESeq2.pdf
» https://www.genomatix.de/online_help/help_regionminer/DESeq2.pdf -
13- Noffz CS, Liedschulte V, Lengeler K, Ernst JF. Functional mapping of the Candida albicans Efg1 regulator. Eukaryotic Cell. 2008;7(5):881-93. doi: doi: 10.1128/EC.00033-08
» https://doi.org/10.1128/EC.00033-08 -
14- Gancedo JM. Control of pseudohyphae formation in Saccharomyces cerevisiae. FEMS Microbiology Reviews. 2001;25(1):107-23. doi: 10.1111/j.1574-6976.2001.tb00573.x
» https://doi.org/10.1111/j.1574-6976.2001.tb00573.x -
15- Pierce VJ, Kumamoto AC. Variation in Candida albicans EFG1 expression enables host-dependent changes in colonizing fungal populations. MBio. 2012;3(4):e00117-12. doi: 10.1128/mBio.00117-12
» https://doi.org/10.1128/mBio.00117-12 -
16- Barros PP, Rossoni RD, Camargo Ribeiro F, Junqueira JC, Jorge AO. Temporal profile of biofilm formation, gene expression and virulence analysis in Candida albicans strains. Mycopathologia. 2017;182(3-4):285-95. doi: 10.1007/s11046-016-0088-2
» https://doi.org/10.1007/s11046-016-0088-2 -
17- Finkel JS, Mitchell AP. Genetic control of Candida albicans biofilm development. Nat Rev Microbiol. 2011;9(2):109-18. doi: 10.1038/nrmicro2475
» https://doi.org/10.1038/nrmicro2475 -
18- Nobile CJ, Fox EP, Nett JE, Sorrells TR, Mitrovich QM, Hernday AD, et al. A recently evolved transcriptional network controls biofilm development in Candida albicans. Cell. 2012;148(1-2):126-38.
-
19- Rosenow CP, Ryan JN, Weiser S, Johnson P, Fontan A, Ortqvist HR, et al. Contribution of novel choline-binding proteins to adherence, colonization, and immunogenicity of Streptococcus pneumoniae. Mol Microbiol. 1997;25(5):819-29. 10.1111/j.1365-2958.1997.mmi494.x
» https://doi.org/10.1111/j.1365-2958.1997.mmi494.x -
20- Hanin M, Brini F, Ebel C, Toda Y, Takeda S, Masmoudi K. Plant dehydrins and stress tolerance: versatile proteins for complex mechanisms. Plant Signal Behav. 2011;6(10):1503-9. doi: 10.4161/psb.6.10.17088
» https://doi.org/10.4161/psb.6.10.17088 -
21- Drira M, Saibi W, Amara I, Masmoudi K, Hanin M, Brini F. Wheat dehydrin K-segments ensure bacterial stress tolerance, antiaggregation and antimicrobial effects. Appl Biochem Biotechnol. 2015;175(7):3310-21. doi: 10.1007/s12010-015-1502-9
» https://doi.org/10.1007/s12010-015-1502-9 -
22- Pramanik A, Garcia E, Ospina R, Powell M, Martinez M, Alejo W, et al. Cloning, characterization and identification of the gene encoding phosphatidylinositol 4-kinase. Cell Mol Biol. 1997;43(7):1007-18.
-
23- Wang Y, Liu J, Zhao H, Lü W, Zhao J, Yang L, et al. Human 1A6/DRIM, the homolog of yeast Utp20, functions in the 18S rRNA processing. Biochim Biophys Acta. 2007;1773(6):863-8. doi: 10.1016/j.bbamcr.2007.04.002
» https://doi.org/10.1016/j.bbamcr.2007.04.002 -
24- Dez C, Dlakić M, Tollervey D. Roles of the HEAT repeat proteins Utp10 and Utp20 in 40S ribosome maturation. RNA. 2007;13(9):1516-27. doi: 10.1261/rna.609807
» https://doi.org/10.1261/rna.609807 -
25- Chen YL, Kauffman S, Reynolds TB. Candida albicans uses multiple mechanisms to acquire the essential metabolite inositol during infection. Infect Immun. 2008;76(6):2793-801. doi: 10.1128/IAI.01514-07
» https://doi.org/10.1128/IAI.01514-07 -
26- Grauslund M, Lopes JM, Rønnow B. Expression of GUT1, which encodes glycerol kinase in Saccharomyces cerevisiae, is controlled by the positive regulators Adr1p, Ino2p and Ino4p and the negative regulator Opi1p in a carbon source-dependent fashion. Nucleic Acids Res. 1999;27(22):4391-8. doi: 10.1093/nar/27.22.4391
» https://doi.org/10.1093/nar/27.22.4391 -
27- Danhof HA, Vylkova S, Vesely EM, Ford AE, Gozalez-Garay M, Lorenz M. Robust extracellular pH modulation by Candida albicans during growth in carboxylic acids. mBio. 2016;7(6):e01646-16. doi: 10.1128/mBio.01646-16
» https://doi.org/10.1128/mBio.01646-16 -
28- Sudbery PE. Growth of Candida albicans hyphae. Nat Rev Microbiol. 2011;9(10):737-48. doi: 10.1038/nrmicro2636
» https://doi.org/10.1038/nrmicro2636 -
29- Calderone RA, Fonzi WA. Virulence factors of Candida albicans. Trends Microbiol. 2001;9(7):327-35. doi: 10.1016/s0966-842x(01)02094-7
» https://doi.org/10.1016/s0966-842x(01)02094-7 -
30- Höfs S, Mogavero S, Hube B. Interaction of Candida albicans with host cells: virulence factors, host defense, escape strategies, and the microbiota. J Microbiol. 2016;54(3):149-69. doi: 10.1007/s12275-016-5514-0
» https://doi.org/10.1007/s12275-016-5514-0 -
31- Ene IV, Heilmann CJ, Sorgo AG, Walker LA, de Koster CG, Munro CA, et al. Carbon source-induced reprogramming of the cell wall proteome and secretome modulates the adherence and drug resistance of the fungal pathogen Candida albicans. Proteomics. 2012;12(21):3164-79. doi: 10.1002/pmic.201200228
» https://doi.org/10.1002/pmic.201200228
Publication Dates
-
Publication in this collection
27 Apr 2020 -
Date of issue
2020
History
-
Received
1 Oct 2019 -
Reviewed
20 Nov 2019 -
Accepted
19 Jan 2020