Abstract
Adenosine deaminase (ADA), histamine, and IgE are endogenously present in animals. Research from this laboratory reported decreased levels of these substances in organs of mice as a consequence of sub-lethal injection of Naja kaouthia venom. This research reports that decreased ADA, histamine, and IgE levels were prevented by specific treatment and prolonged recovery periods. Adult Balb/c mice injected IM with sub-lethal venom dose were divided into five groups. Group 1 were injected with PBS; Group 2 with anti-cobra venom; and Group 3 with lethal toxin neutralizing factor (LTNF). Groups 4 and 5 were treated with IM or oral synthetic LT-10. After 24 hours, mice were sacrificed and organ homogenates were assayed for ADA, histamine, and IgE. Group 1 showed substantial reduction in levels of these substances. It was revealed that decreased levels were prevented by treatment with anti-cobra venom, LTNF, and LT-10. In a second series of experiments, venom-injected mice were sacrificed after 3, 7, and 10 days and organs assayed for ADA, histamine, and IgE levels. The recovery period to homeostasis for ADA, histamine, and IgE was 7 to 10 days.
cobra venom; mouse organs; ADA; histamine; IgE; homeostasis
ORIGINAL PAPER
Restoration of adenosine deaminase, histamine, and IgE in organs of mice injected with cobra venom followed by specific treatment and reversal period
B. V. Lipps
Ophidia Products Inc., Houston, Texas, USA
Correspondence Correspondence to B. V. Lipps Ophidia Products Inc. 11320 South Post Oak, Suite 203 Houston, Texas 77035 USA Fax: 713-663-7290 bvl@ophidia.com
ABSTRACT
Adenosine deaminase (ADA), histamine, and IgE are endogenously present in animals. Research from this laboratory reported decreased levels of these substances in organs of mice as a consequence of sub-lethal injection of Naja kaouthia venom. This research reports that decreased ADA, histamine, and IgE levels were prevented by specific treatment and prolonged recovery periods. Adult Balb/c mice injected IM with sub-lethal venom dose were divided into five groups. Group 1 were injected with PBS; Group 2 with anti-cobra venom; and Group 3 with lethal toxin neutralizing factor (LTNF). Groups 4 and 5 were treated with IM or oral synthetic LT-10. After 24 hours, mice were sacrificed and organ homogenates were assayed for ADA, histamine, and IgE. Group 1 showed substantial reduction in levels of these substances. It was revealed that decreased levels were prevented by treatment with anti-cobra venom, LTNF, and LT-10. In a second series of experiments, venom-injected mice were sacrificed after 3, 7, and 10 days and organs assayed for ADA, histamine, and IgE levels. The recovery period to homeostasis for ADA, histamine, and IgE was 7 to 10 days.
Keywords: cobra venom; mouse organs; ADA; histamine; IgE; homeostasis
INTRODUCTION
Snake envenomation is a complex process involving the direct action of venom components on the tissue and release of various endogenous mediators. Snake venoms are known to cause liberation of pharmacological substances in victims (27). There are reports showing increments in cytokine levels in sera of mice and patients bitten by Bothrops asper and Bothrops jararaca (1,18) and in sera of mice injected with Bothrops atrox venom (2). Petricevich et al. (22) reported increments in serum cytokine and nitric oxide in mice injected with B. asper and B. jararaca venoms. Both venoms induced marked increases in tumor necrosis factors and interleukins IL-1, IL-6, IL-10 in sera of injected mice. Liver and kidney of rabbits and cardiac muscles of rats were affected by Egyptian cobra venom injection (24,25). The effects of a sub-lethal dose IM injection of the Egyptian cobra snake venom on histological and histochemical pattern of rabbit kidney and liver have been demonstrated (23).
Adenosine deaminase (ADA) is an essential enzyme that controls levels of biologically active purines and 2'-desoxy adenosine in tissues and cells (6). ADA deficiency results in combined immunodeficiency by a severe T, B, and NK cell lymphopenia (7). ADA deficiency is also associated with bony and renal abnormalities, hepatocellular damage, neurological disorders, and pulmonary insufficiency (3,9). Some ADA deficient patients have been reported with elevated IgE and eosinophilia, which are implicated in allergy-asthma (5).
Fagan et al. (4) reported that monoclonal antibodies to IgG4 induce histamine release from human basophils in vitro. Evidence of histamine release was demonstrated as a principal pharmacological component of Australian wolf spider venom in comparison to the control (26). The earliest demonstration of IgE antibodies against venoms in sera of victims of fatal anaphylaxis from stings was reported (8). Both IgG and IgE antibody levels were shown to rise with bee and wasp venom immunotherapy (10).
Sub-lethal injection of cobra venom caused decrease in levels of ADA, histamine, and IgE in organs of mice (15). Lipps (14) also reported decreased levels of nerve growth factor (NGF) in organs of mice as a consequence of sub-lethal injection of cobra venom. Lipps (16) further reported the restoration of NGF levels in organs of mice injected with cobra venom, by specific treatment and reversal periods for return to normal homeostasis.
Menchaga and Perez (20) isolated and purified antihemorrhagic protein from opossum (Didelphis virginiana) serum of 68,000 kDa molecular weight. Perales et al. (21) isolated an anti-bothropic fraction (ABF) from sera of several South American Didelphidae (Didelphis marsupialis, Philander opossum, and Lutreolina crassicaudata). Lipps (11) isolated lethal toxin neutralizing factor (LTNF) from opossum serum, having a neutralizing effect on animal, plant, and bacterial toxins when tested in mice. Lipps (12) further identified the active LTNF domain consisting of ten amino acids referred to as LT-10. Synthetic LT-10 showed a neutralizing effect similar to natural LTNF.
This investigation reports that decreased ADA, histamine, and IgE levels in organs of mice caused by cobra venom injection were prevented by specific treatment with anti-cobra venom, natural and synthetic LTNFs, and prolonged reversal periods.
MATERIALS AND METHODS
Venom
Venom was collected from several adult Naja kaouthia snakes at various times during a calendar year and frozen. The stored venom was thawed and pooled. The pooled venom was aliquoted and stored at -20°C.
Animals
Adult Balb/c were purchased from Harlan Teklad, Madison, WI. Mice were used in compliance with U. S. Public Health Service policy on humane care and use of animals.
Production of anti-IgE
IgE was purchased from Scripps Laboratory. Anti-IgE was produced by immunizing adult New Zealand rabbits by IM injection of 250 µg of IgE per rabbit three times two weeks apart. The first injection consisted of a mixture of IgE and Freund's complete adjuvant (FCA). Subsequent injections consisted of IgE mixed with Freund's incomplete adjuvant (FICA).
Anti-cobra venom
Anti-cobra venom was made by immunizing adult New Zealand rabbits against Naja kaouthia venom of ELISA titer 1:24300/100 µl.
Production of polyclonal antibodies to ADA and histamine in mice
ADA and histamine antisera are not commercially available. Antibodies against ADA and histamine were produced in mice as described by Lipps and Khan (17). ADA and histamine were purchased from Sigma-Aldrich Co. Adult Balb/c mice were immunized by IM inoculation. The first injection consisted of 0.2 ml/mouse containing 100 µg of ADA or histamine in PBS mixed with an equal volume of FCA. Subsequently, three additional injections were given two weeks apart, consisting of the same concentration mixed with FICA. Immunized mice were bled two weeks after the last injection through the ophthalmic vein and serum was collected.
Natural LTNF
LTNF was isolated from opossum serum (11), with antidote property for animal, plant, and bacterial toxins.
Synthetic LTNF ( Syn LT-10)
Synthetic LT-10 was made with ten amino acids, the active domain of the natural LTNF having sequence from N-terminal: Leu-Lys-Ala-Met-Asp-Pro-Thr-Pro-Pro-Leu (12).
Treatment to prevent decrease of ADA, histamine, and IgE levels in organs of mice
Lethal dose of N. kaouthia venom was determined by injecting IM at various concentrations in mice. The lethal dose for 16 to 18g mouse was found to be 4 µg. Therefore, mice having similar weights were injected with 2 µg in 0.2 ml. Thirty mice were used; five controls were injected with 0.2 ml PBS.
Venom-injected mice were divided into five groups each consisting of five mice and were treated within half an hour. Group 1 were treated IM with 0.5 ml PBS; Group 2 with 0.5 ml anti-cobra venom; and Group 3 with 0.5 ml containing 200 µg of LTNF. Groups 4 and 5 were treated with synthetic LT-10. Group 4 were injected IM with 0.5 ml containing 500 µg of LT-10, and Group 5 were given orally the same amount of LT-10 orally three times within half an hour. For ADA, histamine, and IgE restoration, the venom injected mice were sacrificed after 24 hours and results are shown in Figures 1, 3 Figure 3. Restoration of Histamine. , and 5 Figure 5. Restoration of IgE. . For ADA, histamine, and IgE recovery, the venom injected mice were sacrificed after 3, 7 and 10 days and results are shown in Figures 2, 4 Figure 4. Recovery of Histamine. , and 6 Figure 6. Recovery of IgE. Preparation of organ suspension .
The pool of organs was homogenized in PBS with a manual homogenizer. The homogenates were centrifuged and supernatants for each pool organ were separated. Protein concentration for each supernatant was measured on a spectrophotometer using a protein kit from Bio-Rad (catalogue 500-0006). Supernatant protein concentrations were adjusted to 100 µg/ml with PBS and kept frozen for further testing.
Enzyme-linked immunosorbent assay (ELISA)
ELISA tests were carried out in 96-well microtiter plates as described by Lipps (13). The reagents for ELISA were purchased from Sigma-Aldrich Co. The stocks from mice organs were diluted with carbonate-bicarbonate buffer, pH 9.4, to 10 µg/ml as coating antigen for ELISA. The wells of the plate were coated with antigen, 100 µl/well. The plate was left at room temperature (RT) overnight, after which it was emptied and washed three times (3x) with PBS and the wells were blocked with 250 µl/well of 3% fish skin gelatin. After 30 min, the plate was emptied and washed 3x with PBS. Antisera against ADA, histamine, and IgE diluted in gelatin were added. After pilot testing, three wells received 100 µl of each dilution of antiserum, diluted two-fold from 1:300. Appropriate antigen and antibody controls were incorporated. The plate was incubated at 37°C in a humid incubator for 1 hour after which the plate was washed 3x with PBS. Horseradish peroxidase conjugated with mouse IgG for ADA and histamine assays was used. For IgE assay, horseradish peroxidase conjugated with rabbit IgG was used. Horseradish peroxidase 100 µl/well was reacted for 30 min. Finally, after washing, the wells were reacted with O-phenylenediamine-HCl for color development. The plate was read after 30 min, and the OD was recorded at 405 mm as ELISA titer per 100 µl.
RESULTS
Balb/c female mice injected IM with sub-lethal dose of cobra venom, were treated with anti-cobra venom, LTNF, and LT-10 as described above. After 24 hours, mice in all groups were sacrificed and the organ homogenates were assayed for ADA, histamine, and IgE by simple ELISA test using respective antisera and compared to control organs of mice not injected with venom. The organs tested were: brain, heart, kidney, liver, lung, muscle, pancreas, salivary gland, spleen, and ovary. Figures 1 to 6 show the organs with considerable restoration due to specific treatment and prolonged recovery period returning to normal homeostasis.
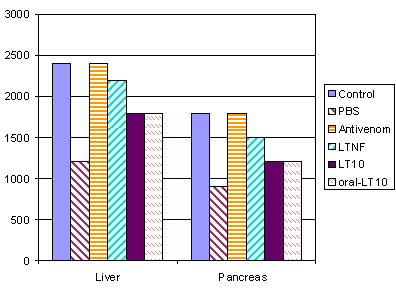
Prevention of decreased ADA levels by specific treatment and recovery period for normal homeostasis in liver and pancreas
Balb/c mice injected IM with sub-lethal dose of cobra venom were treated with anti-cobra venom, LTNF, and LT-10 as described above. After 24 hours, mice in all groups were sacrificed and organ homogenates were assayed for ADA by simple ELISA. Results of liver and pancreas are shown in Figures 1 and 2.
Figure 1 shows that treatment with anti-cobra venom and LTNF prevented decrease in ADA levels and homeostasis was restored for liver and pancreas. Oral or IM treatment with Syn-LT-10 also prevented decrease in ADA levels, to some extent, in liver, and pancreas.
Figure 2 shows that ADA homeostasis for liver and pancreas returned after 10 days.
Prevention of decreased histamine levels by specific treatment and recovery period for normal homeostasis in heart and lung
Balb/c mice injected IM with sub-lethal dose of cobra venom were treated with anti-cobra venom, LTNF, and LT-10 as described above. After 24 hours, mice in all groups were sacrificed and the organ homogenates were assayed for histamine by ELISA. Results of heart and lung are shown in Figures 3 Figure 3. Restoration of Histamine. and 4 Figure 4. Recovery of Histamine. .
Figure 3 Figure 3. Restoration of Histamine. shows that the treatment with anti-cobra venom and LTNF prevented the decrease in histamine levels and homeostasis was restored for heart and lung. Oral and IM treatment with Syn-LT-10 also prevented decrease in histamine levels in heart and lung.
Figure 4 Figure 4. Recovery of Histamine. shows that histamine homeostasis for heart and lung returned to normal after 7 days.
Prevention of decreased levels of IgE by specific treatment and recovery period for normal homeostasis in heart and lung
Balb/c mice injected IM with sub-lethal dose of cobra venom were treated with anti-cobra venom, LTNF, and LT-10 as described above. After 24 hours, mice in all groups were sacrificed and organ homogenates were assayed for IgE by ELISA. Results of heart and lung are shown in Figures 5 Figure 5. Restoration of IgE. and 6 Figure 6. Recovery of IgE. Preparation of organ suspension .
Figure 5 Figure 5. Restoration of IgE. shows that treatment with both anti-cobra venom and LTNF prevented decrease in IgE levels in heart and lung, but there was more decrease with anti-cobra venom than LTNF. Homeostasis was restored for heart and lung. Oral and IM treatment with Syn-LT-10 prevented minuscule decrease in IgE levels in heart and lung.
Figure 6 Figure 6. Recovery of IgE. Preparation of organ suspension shows that IgE recovery for heart remained low after 10 days. However, the normal IgE homeostasis for lung returned after 10 days.
DISCUSSION
Currently, ELISA tests are carried out using blood serum or blood plasma for assaying proteins from serum or plasma. When serum is used as an antigen to assay different parameters, other tests such as radioimmunoassay (RIA), radioallergosorbent test (RAST), or double sandwich ELISA can be performed. However, these tests are complicated and require more time and special reagents. In this paper, simple ELISA test was used comprising for example, an antigen ADA of mouse organ reacting with various concentrations of anti-ADA yielding ELISA titer/100 µl at the lowest concentration of antibody. This simple ELISA cannot be performed to assay any proteins from serum or plasma.
Investigations concerning changes taking place in blood serum as a consequence of envenomation in humans and experimental animals is a subject of many years of research. Increased glucose and lactate concentrations were known to be common in snake envenomation (19). Increased levels of various cytokines have been reported as a consequence of snake and scorpion envenomation (22, 28). There is yet no clear relationship between venom toxicity and increased glucose or increased levels of various cytokines in sera of animals injected with venom. Research from this laboratory showed decreased levels of various endogenously present pharmacological substances, NGF, ADA, histamine, and IgE in organs of mice injected with sub-lethal dose of cobra venom (14, 15). Nevertheless, the relationship between the decrease in these substances and toxicity remains unclear. Although, all organs were tested, few organs showed decreased ADA, histamine, and IgE levels. Why all organs did not show the effect is not clear at this time. Further research is warranted to elucidate this phenomenon.
This investigation reports the prevention of decreased ADA, histamine, and IgE levels taking place in the organs of mice injected with cobra venom. Lethality in mice injected with cobra venom is prevented by anti-cobra venom. Like wise, Lipps reported that lethality of cobra venom in mice is prevented by LTNF and LT-10 treatment (11, 12). The results of this investigation showed that decreased ADA, histamine, and IgE levels as a consequence of venom injection were prevented if the mice were treated with anti-cobra venom, LTNF, and LT-10. Therefore, decreased level or loss of ADA, histamine, and IgE may be somehow related to venom toxicity, which is not clear. Published research reported that 500 µg of synthetic peptide LT-10 consisting of ten amino acids inhibited N. kaouthia venom lethality in mice by IM route. It is pleasing to find that oral LT-10 produced similar effect in restoring ADA, histamine, and IgE levels. Thus, LT-10 can be advocated by oral route as a therapeutic agent against envenomation.
ACKNOWLEDGEMENTS
The author acknowledges her husband Frederick Lipps for preparing the graphics. This research was supported by Ophidia Products Inc.
Received September 9, 2002
Accepted February 13, 2003
- 1 BARRAVIERA B., LOMONTE B., TARKOWSKI A., HANSON LA., MEIRA DA. Acute-phase reactions, including cytokines, in patients bitten by Bothrops and Crotalus snakes in Brazil. J. Venom. Anim. Toxins, 1995, 1, 11-22.
- 2 BARROS SF., FRIEDLANSKAIA I., PETRICEVICH VL., KIPNIS TL. Local inflammation, lethality and cytokine release in mice injected with Bothrops atrox venom. Mediat. Inflamm., 1998, 7, 339-46.
- 3 BOLLINGER ME., ARREDONDO-VEGA FX., SANTISTEBAN I., SCHWARZE K., HIRSHFIELD MS., LEDERMAN HM. Brief report: hepatic dysfunction as a complication of adenosine deaminase deficiency. N. Engl. J. Med., 1996, 334, 1367-71.
- 4 FAGAN DL., SLAUGHTER CA., CAPRA JD., SULIVAN TJ. Monoclonal antibodies to immunoglobin G4 induce histamine release from human basophil in vitro. J. Allergy Clin. Immunol, 1982, 70, 399-404.
- 5 FOZARD JR., PFANNKUCH HJ., SCHUURAM HJ. Mast cell degranulation following adenosine A3 receptor activation in rats. Eur. J. Pharmacol., 1997, 298, 293-7.
- 6 FREDERIKSEN S. Specificity of adenosine deaminase toward adenosine and 2'-deoxyadenosine analogues. Arch. Bichem. Biophys., 1966, 113, 383-8.
- 7 HIRSCHHORN R. Immunodeficiency disease due to deficiency of adenosine deaminase. In: OCHS HD., SMITH CIE., PUCK JM. Eds. Primary immunodeficiency disease: a molecular and genetic approach. New York: Oxford University Press, 1999: 121-38.
- 8 HOFFMAN DR., WOOD DL., HUDSON P. Demonstration of IgE and IgG antibodies against venoms in the blood of victims of fatal anaphylaxis. J. Allergy Clin. Immunol, 1983, 73, 193-6.
- 9 JACKSON MA., BAI TR. The role of adenosine in asthma. In: JACOBSON KA., JARVIS MF. Eds. Purinergic approaches in experimental therapeutics Danvers: Wiley-Liss, 1997: 315-31.
- 10 KEMENY DM., LESSOF MH., PATEL S., YOULTEN LJ., WILLIAMS A., LAMBOURN E. IgG and IgE antibodies after immunotherapy with bee and swap venom. Int. Arch. Allergy Appl. Immunol., 1989, 88, 247-9.
- 11 LIPPS BV. Anti-lethal factor from opossum serum is a potent antidote for animal, plant and bacterial toxins. J. Venom. Anim. Toxins, 1999, 5, 56-66.
- 12 LIPPS BV. Small synthetic peptides inhibit, in mice, the lethality of toxins derived from animal, plant and bacteria. J. Venom. Anim. Toxins, 2000, 6, 77-86.
- 13 LIPPS BV. Isolation of subunits, a, b and g of the complex taipoxin from the venom of Australian Taipan snake (Oxyuranus s. scutellatus): characterization of beta taipoxin as a potent mitogen. Toxicon, 2000, 38, 1845-54.
- 14 LIPPS BV. Decreased levels of nerve growth factor in organs of mice as a consequence of sub-lethal injection of cobra venom. J. Nat. Toxins, 2001, 10, 283-90.
- 15 LIPPS BV. Sub-lethal injection of cobra venom decreased levels of adenine deaminase, histamine and IgE in organs of mice. J. Venom. Anim. Toxins, 2002, 8, 60-73.
- 16 LIPPS BV. Restoration of nerve growth factor in organs of mice injected with cobra venom followed by specific treatment and reversal period. J. Nat. Toxins, 2002, 11, 87-93.
- 17 LIPPS BV., KHAN AA. The presence of pharmacological substances myoglobin and histamine in venoms. J. Venom. Anim. Toxins, 2001, 7, 45-55.
- 18 LOMONTE B., TARKOWSKI A., HANSON LA. Host response to Bothrops asper snake venom: analysis of edema formation, inflammatory cells and cytokine release in mouse model. Inflammation, 1993, 17, 93-105.
- 19 MARSH NA., GATTULLO D., PAGLIARO P., LOSSANO G. The Gaboon viper Bitis gabonica, hemorrhagic, metabolic, cardiovascular and clinical effects of the venom. Life Sci., 1997, 61, 763-9.
- 20 MENCHAGA JM., PEREZ JC. The purification and characterization of antihemorrhagic factor in opossum (Didelphis virginiana) serum. Toxicon, 1981, 19, 623-32.
- 21 PERALES J., MOUSSATCHE H., MARANGONI S., OLIVEIRA B., DOMONT GB. Isolation and partial characterization of an anti-bothropic complex from the serum of South American Didelphidae. Toxicon, 1994, 32, 1237-49.
- 22 PETRIVICH VL., TEIXEIRA CFP., TAMBOURGI DV., GUTIERREZ JM. Increments in serum cytokine and nitric oxide levels in mice injected with Bothrops asper and Bothrops jararaca snake venoms. Toxicon, 2000, 38, 1253-66.
- 23 RAHMY TR., HEMMAID KZ. Histological and histochemical alterations in the liver following intramuscular injection with sublethal dose of Egyptian cobra venom. J. Nat. Toxins, 2000, 9, 21-32.
- 24 RAHMY TR., RAMADAN RA., FARID TM., EL-ASMAR MF. Hepatic pathogenesis due to cobra envenomation. J. Egypt. Ger. Soc. Zool, 1995, 16, 85-102.
- 25 RAHMY TR., RAMADAN RA., FARID TM., EL-ASMAR MF. Renal lesions induced by cobra envenomation. J. Egypt. Ger. Soc. Zool, 1995, 17, 251-71.
- 26 RASH LD., KING RG., HODSON WC. Evidence that histamine is the principal pharmacological component of venom from an Australian wolf spider (Lycosa godeffroyi). Toxicon, 1998, 36, 367-75.
- 27 ROTHCHILD AM., ROTHCHILD Z. Liberation of pharmacologically active substances by snake venom. In: Lee CY. Ed. Snake venoms Berlin: Springer-Verlag, 1979: 541.
- 28 SOFER S., GUERON M., WHITE RM., LIFSHITZ M., APATE RN. Interleukin-6 release following scorpion sting in children. Toxicon, 1996, 34, 389-92.




Publication Dates
-
Publication in this collection
09 Dec 2003 -
Date of issue
2003
History
-
Received
09 Sept 2002 -
Accepted
13 Feb 2003