
Abstract
Copulation in Opisthogonimus fonsecai Ruiz & Leão, 1942 (Trematoda, Digenea, Plagiorchiidae), parasite of Bothrops moojeni Hoge, 1966 (Serpentes, Viperiade), was reported. Data on the histology of the cirrus and metraterm of the copulants were presented. Morphology of the cirrus was also examined by scanning electron microscopy.
copulation; Opisthogonimus fonsecai; Trematoda; histology; SEM
ORIGINAL PAPERS
Copulation of Opisthogonimus fonsecai Ruiz & Leão, 1942 (Trematoda, Digenea, Plagiorchiidae), parasite of Bothrops moojeni Hoge, 1966 (Serpentes, Viperidae)
Silva R. J.
Departament of Parasitology, Institute of Biosciences, Unesp, Botucatu, São Paulo, Brazil
Correspondence Correspondence to R. J. SILVA Departamento de Parasitologia, Instituto de Biociências, Unesp 18618-000, Botucatu, São Paulo, Brasil Phone: 55 14 3811 6239 Email: reinaldo@ibb.unesp.br
ABSTRACT
Copulation in Opisthogonimus fonsecai Ruiz & Leão, 1942 (Trematoda, Digenea, Plagiorchiidae), parasite of Bothrops moojeni Hoge, 1966 (Serpentes, Viperiade), was reported. Data on the histology of the cirrus and metraterm of the copulants were presented. Morphology of the cirrus was also examined by scanning electron microscopy.
Key words: copulation, Opisthogonimus fonsecai, Trematoda, histology, SEM.
INTRODUCTION
Most digenean trematode species are hermaphrodites. Literature reports that self-fertilization may happen but it is uncommon, since a number of flukes usually occur in the same host. Reproduction of some of these helminths involves copulation, which happens when the cirrus of one fluke is inserted into the terminal portion of the other fluke's uterus (metraterm) (14).
Studies on the mating behavior of adult digenean trematodes have been carried out with some species of the families Philophthalmidae (5,6,7), Echinostomidae (9,10), and Paramphistomidae (11). In these reports, self and cross-insemination patterns were observed by using transplantation and autoradiography techniques. Besides, Sey et al. (13) reported the copulation of Lasiotocus sp. by scanning electron microscopy (SEM).
The copulatory organ of most digenean is a cirrus. Frequently, it has circular and longitudinal muscles in its walls and presents spines (4). The metraterm is muscular to varying degrees in different forms and may serve as a vagina (4).
Although some reports about trematodes reproduction are available in literature, histological and SEM data on the copulation of trematode parasites are scarce. Thus, the objective of the present study was to report copulation in Opisthogonimus fonsecai (Trematoda, Digenea, Plagiorchiidae).
MATERIAL AND METHODS
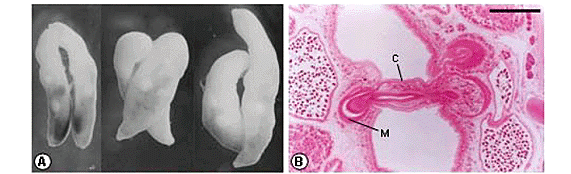
The trematodes were collected from the mouth of a Bothrops moojeni (Serpentes, Viperidae) specimen from Porto Primavera, São Paulo State, and fixed in cold AFA solution without compression (2). Some trematodes were stained with carmine and mounted in Permount resin. The O. fonsecai (Figure 1) species was identified (1,12,15). Among the collected helminths, three pairs of copulants were found (Figure 2A). One of them was studied by SEM, and the other two were analyzed after histology. For the histological study, fixed helminths were embedded in historesin (2-hydroxyethyl methacrylate); 4-mm transverse serial sections were prepared, stained with hematoxylin-eosin and examined by using a computerized image analysis system (Qwin Lite 2.5 - Leica).
RESULTS
In each of the three cases of copulation observed, only one of the copulants had its cirrus introduced into the metraterm of the other fluke. Trematodes that were receiving the cirrus had their own intromittent organ everted in one case and withdrawn in the other two cases. The trematodes were parallel or diagonal to one another. In two cases, the copulants had similar size, and in the third, they were slightly different (Figure 2A).
In one case histologically examined, the two trematodes had the genital atrium at a similar level; and in the transverse histological sections, it was observed that the cirrus of one specimen was introduced into the metraterm of the other (Figure 2B). The longest cirrus observed measured 636 mm. The extruded extent of the cirrus was 414 mm, but only 121 mm was introduced into the other fluke's metraterm (Figure 2C). The widest cirrus measured 134 mm in diameter; and a constriction, which resulted in a decrease of the cirrus diameter to 42 mm, was observed at the genital pore of the trematode that was receiving the cirrus (Figure 2C). The cirrus presented a canal (ejaculatory duct) with a lumen of 6-7 mm in diameter. This duct was covered with a mucous epithelium of 8-11.5 mm thickness and two layers of smooth musculature: a circular one, with 1-1.5 mm thickness, and a longitudinal one, with 1-3 mm thickness. Externally, the cirrus was covered with a tegument of 9.5-12 mm thickness. Under this tegument, there was a basal lamina of 0.5-1 mm thickness and two layers of smooth musculature: a circular one, with 2.5-3 mm thickness, and a longitudinal one, with 1-2 mm thickness. Internally, there was a parenchyma and also glands. The cirrus had no spines (Figure 2D).
In the other case histologically studied, the cirrus was in a diagonal position; and after the analysis of all histological serial sections, it was observed that only a small portion of the cirrus' extremity was introduced into the metraterm of the other fluke (Figure 2E). In this second case, we obtained a good section of the metraterm with a great amount of spermatozoa in the lumen (Figures 2E-F). This metraterm was surrounded by a large number of glands (Figure 2E-F) and measured 288 mm long and 125.5 mm wide. The lumen of this structure was 58 mm in diameter. There was a mucous epithelium of 8.5-15 mm thickness; a basal lamina of 1-2.5 mm thickness; and two layers of smooth muscle: a circular one, with strong muscle fibers of 15.5-21 x 1-1.5 mm thickness, and a longitudinal one of 2-5.5 mm thickness (Figure 2F).
SEM examination showed that the cirrus had no spines and was tapered. Next to the metraterm's opening, we could also observe some eggs of this trematode adhered to the cirrus (Figures 2G-H).
DISCUSSION
The male and female reproductive systems of O. fonsecai were described by Ruiz & Leão (12). Bothrops moojeni infection by this species was previously reported by Correa (3). However, there are no reports on the copulation of this trematode.
Patterns of sexual reproduction in platyhelminths were discussed by Nollen (8). According to this author, digeneans have potential for both cross and self-insemination, since they are almost exclusively hermaphrodite and, in most of them, the male and female systems share a common genital atrium or pore. In addition, he cited other papers in which insemination through Laurer's canal in the species Diphterostomum brusinae and Haploporus benedeni; mating via metraterm in Podoctyle fractum; reciprocal copulation in Fasciola hepatica; and self-insemination in Leucochloridiomorpha constatinae were observed. For many digeneans, cross-fertilization is usual. On the other hand, at least in two trematodes (Liolope copulans and Collyriclum faba), Laurer's canal was used for the reception of the other individual's cirrus during copulation and for the conveyance of the spermatozoa received during this act to the seminal receptacle (4).
In the two specimens histologically examined in this report, the cirrus was introduced only into the distal portion of the metraterm. For trematodes, it is known that co-copulants start to separate when removed from the host or fixed. This have happened to the pairs of copulants observed in this report and may explain the presence of the cirrus only in the distal portion of the metraterm or even the lack of reciprocal copulation. However, we believe that, during copulation, the full extent of the metraterm is occupied by the cirrus.
REFERENCES
Received: December 10, 2003
Accepted: July 14, 2004
Published online: February 11, 2005.
- 1 ARTIGAS PT., PEREZ MD. Sistemática dos Opisthogonimidae (Trematoda, Plagiorchoidea). Criação da Família Bieriidae N. Fam. Mem. Inst. Butantan, 1969, 34, 97-110.
- 2 BARRELLA TH., SILVA RJ. Infection by digenetic trematodes in a Bothrops moojeni population (Serpentes: Viperidae) from a fauna rescue in Porto Primavera region, São Paulo State, Brazil. Arq. Bras. Med. Vet. Zootec., 2003, 55, 243-5.
- 3 CORRÊA AAS. Fauna de trematóides parasitos de ofídios da área geográfica brasileira São Paulo: Universidade de São Paulo, Instituto de Ciências Biológicas, 1990. 188p. [Tese - Doutoramento]
- 4 DAWES B. The trematoda Cambridge: Cambridge University Press, 1968. 644p.
- 5 MOSELEY C., NOLLEN PM. Autoradiographical studies on the reproductive system of Philophtalmus hegeneri Penner and Fried, 1963. J. Parasitol, 1973, 59, 650-4.
- 6 NOLLEN PM. Autoradiographical studies on reproduction in Philophtalmus megalurus (Cort. 1914) (Trematoda). J. Parasitol, 1968, 54, 43-8.
- 7 NOLLEN PM. Autoradiographical studies on the reproductive system of Philophtalmus grali using techniques of transplantation and autoradiography. J. Parasitol, 1978, 64, 613-6.
- 8 NOLLEN PM. Pattern of sexual reproduction among parasitic platyhelminths. Parasitology, 1983, 86, 99-120.
- 9 NOLLEN PM. Echinostoma caproni: mating behavior and the timing of development and movement of reproductive cells. J. Parasitol., 1990, 76, 784-9.
- 10 NOLLEN PM. Echinostoma trivolvis: mating behavior of adults grown in hamsters. Parasitol. Res, 1993, 79, 130-2.
- 11 NOLLEN PM., PYNE JL. Observations on spermatogenesis and inseminative behavior of Megalodiscus temperatus adults in frogs. J. Parasitol, 1979, 65, 35-7.
- 12 RUIZ JM., LEÃO AT. Três novas espécies de Opisthogonimus parasitas de ofídeos brasileiros (Trematoda: Plagiorchiidae). Mem. Inst. Butantan, 1942, 16, 171-81.
- 13 SEY O., AL-GHAITH L., NAHHAS FM. Scanning electron microscopy of a copulating Monorchiid (Trematoda: Digenea). J. Helminthol, 1998, 65, 243-5.
- 14 SMYTH JD. The physiology of trematodes San Francisco: WH Freeman and Company, 1966. 256p.
- 15 TRAVASSOS L., FREITAS JFT., KOHN A. Trematódeos do Brasil. Mem. Inst. Oswaldo Cruz, 1969, 67, 1-886.
Correspondence to
Publication Dates
-
Publication in this collection
03 May 2005 -
Date of issue
Mar 2005
History
-
Received
10 Dec 2003 -
Reviewed
14 July 2004 -
Accepted
11 Feb 2005




