Abstract
TsTX is an α-type sodium channel toxin that stimulates the discharge of neurotransmitters from neurons. In the present study we investigated which neurotransmitters are released in the hippocampus after TsTX injection and if they are responsible for electrographic or histopathological effects. Microdialysis revealed that the toxin increased glutamate extracellular levels in the hippocampus; however, levels of gamma-aminobutyric acid (GABA), glycine, 5-hydroxyindoleacetic acid (5-HIAA), homovanillic acid (HVA) and 3,4-dihydroxyphenylacetic acid (DOPAC) were not significantly altered. Neurodegeneration in pyramidal cells of hippocampus and electroencephalographic alterations caused by the toxin were blocked by pretreatment with riluzole, a glutamate release inhibitor. The present results suggest a specific activity of TsTX in the hippocampus which affects only glutamate release.
scorpion toxin; microdialysis; hippocampus; neurotransmitters; neurodegeneration; riluzole
ORIGINAL PAPER
Neurotransmitter evaluation in the hippocampus of rats after intracerebral injection of TsTX scorpion toxin
Nencioni ALAI; Barreto SAII; Lebrun III; Florio JCIII; Lourenço GAI; Dorce VACI
ILaboratory of Pharmacology, Butantan Institute, São Paulo, São Paulo State, Brazil
IILaboratory of Biochemistry and Biophysics, Butantan Institute, São Paulo, São Paulo State, Brazil
IIILaboratory of Pharmacology, School of Veterinary Medicine and Zootechny, University of São Paulo, USP, São Paulo, São Paulo State, Brazil
Correspondence to Correspondence to: Valquiria Abrão Coronado Dorce Laboratório de Farmacologia, Instituto Butantan Av. Dr. Vital Brasil, 1500 São Paulo, SP, 05503-900, Brasil Phone: +55 11 3726 7222, ext. 2133. Fax: +55 11 3726 1505 Email: vdorce@butantan.gov.br
ABSTRACT
TsTX is an α-type sodium channel toxin that stimulates the discharge of neurotransmitters from neurons. In the present study we investigated which neurotransmitters are released in the hippocampus after TsTX injection and if they are responsible for electrographic or histopathological effects. Microdialysis revealed that the toxin increased glutamate extracellular levels in the hippocampus; however, levels of gamma-aminobutyric acid (GABA), glycine, 5-hydroxyindoleacetic acid (5-HIAA), homovanillic acid (HVA) and 3,4-dihydroxyphenylacetic acid (DOPAC) were not significantly altered. Neurodegeneration in pyramidal cells of hippocampus and electroencephalographic alterations caused by the toxin were blocked by pretreatment with riluzole, a glutamate release inhibitor. The present results suggest a specific activity of TsTX in the hippocampus which affects only glutamate release.
Key words: scorpion toxin, microdialysis, hippocampus, neurotransmitters, neurodegeneration, riluzole.
INTRODUCTION
The toxicity of scorpion venoms is due to neurotoxins that are composed of basic polypeptides with molecular weights of approximately 7,000 Da. These toxins affect cell permeability to ions including sodium, potassium, chloride and calcium (1-4). Sodium channel neurotoxins affect sodium conductance in various excitable tissues, thus serving as important pharmacological tools for the study of excitability and sodium channel structure (5).
Scorpion neurotoxins that act on sodium channels have been divided into two groups, α and β toxins, according to the ligand-binding sites in the channel. The α-scorpion toxins were the first to be studied. Their primary effect is interference in channel inactivation upon prolonged depolarization, increasing sodium permeability and consequently extending the duration of the action potential (6). One of the main consequences is the augmented release of neurotransmitters from neuronal endings (7).
Gomez and Diniz (8) were the first to isolate toxic polypeptides from the venom of the Brazilian scorpion Tityus serrulatus. According to Arantes et al. (9), TsTX was first isolated by Coutinho Netto in 1975. This α-type sodium channel toxin increases the release of glutamate, acetylcholine and GABA from different preparations of nervous tissue (10-13). When injected in the dorsal hippocampus of rats, it induces behavioral alterations and epileptic discharges besides neurodegeneration (14).
The aim of the present study was to investigate which neurotransmitters are released after TsTX intrahippocampal injection. To do this, microdialysis analysis was performed to evaluate extracellular levels of some cerebral neurotransmitters.
In a previous study, we observed an increase in extracellular levels of glutamate only until three hours after toxin injection (11). Based on the toxin kinetic and on the glutamate release profile observed in that study, we decided to lengthen the time of collection (15).
Moreover, since the previous work by Nencioni et al. (11) had indicated the involvement of glutamate in the convulsive/neurodegenerative effect of the toxin; in the present study, we examined the result of pretreatment with riluzole (2-amino-6-trifluoromethoxy benzothiazole, RP54274), a drug that inhibits glutamate release and presents neuroprotective, anticonvulsant, anxiolytic and anesthetic properties (16).
MATERIALS AND METHODS
Subjects
Male Wistar rats (200 to 250 g), obtained from an established colony kept by the Central Animal House Service at the Butantan Institute, were used. Upon their arrival in the laboratory (seven days before the experiments), the animals were individually housed in wire mesh cages and kept at constant temperature (22 ± 1° C), in a 12-hour light/12-hour dark cycle (lights on at 0700 h), with food and water provided ad libitum.
The animals employed in the current study were maintained in accordance with the policies of the Ethics Committee on Use of Laboratory Animals, Butantan Institute, Brazil.
TsTX Purification
Tityus serrulatus toxins were obtained according to the method described by Carvalho et al. (14). Briefly, the venom was extracted with 0.05 M ammonium bicarbonate buffer, pH 8.0, and eluted in Sephadex® G-50 M (Pharmacia Fine Chemicals, USA). Active fractions 30 to 63 (pool IV) were eluted with ammonium bicarbonate (0.05 M, pH 8.0) in Sephadex® G-25 M column (Pharmacia Fine Chemicals, USA). The peptides purified by gel filtration chromatography were separated by high performance liquid chromatography (HPLC) and the peak 8 was used (TsTX).
Chemicals
-
Ringer solution (Aster, Brazil)
-
TsTX scorpion toxin (Butantan Institute, Brazil) dissolved in Ringer solution
-
L-glutamic acid HCl (Research Biochemicals International, USA)
-
L-aspartic acid (Research Biochemicals International, USA)
-
Glycine (Sigma, USA)
-
γ -aminobutyric acid (Sigma, USA)
-
Riluzole hydrochloride (Tocris, USA) dissolved in 10% dimethyl sulfoxide (DMSO Sigma, USA)
-
Phenyl isothiocyanate (Sigma, USA)
-
Sodium acetate (Merck, Germany)
-
Triethylamine (Sigma, USA)
-
Ethylenediaminetetraacetic acid (EDTA Merck, Germany)
-
Acetonitrile (Malinkrodt, USA)
-
3,4-dihydroxybenzylamine (DHBA Sigma, USA)
-
Perchloric acid (Merck, Germany)
-
Sodium metabisulfit (Merck, Germany)
Surgery
Rats were anesthetized with an intraperitoneal (IP) injection of 3 mL/kg of a mixture of pentobarbitone (1 g) and chloral hydrate (4 g) diluted in 100 mL of 0.9% NaCl. Animals were placed in a stereotaxic frame and a microdialysis guide cannula was implanted unilaterally in their dorsal hippocampus. Guide cannulas were fixed to the skulls with stainless steel jeweler screws and dental acrylate. The guide cannulas for microdialysis were implanted in rats according to the following coordinates: 5.3 mm posterior to bregma, 3.2 mm lateral to midline, and 2 mm below dura mater (17). The cannulas were used for both toxin injection and dialysate collection. In rats undergoing behavioral studies and EEG recording, the coordinates were 4.8 mm posterior to bregma, 3.2 mm lateral to midline, and 2.8 mm below dura mater. In these rats, bipolar twisted electrodes for depth recordings were implanted contralaterally at the same coordinates and anchored to the skull as were the cannulas. Jeweler screws were inserted bilaterally in the skull over the occipital cortex for surface recordings. A screw placed in the frontal sinus was employed as reference (indifferent electrode). The surgery was conducted using aseptic techniques and animals with surgical complications were excluded. Subsequently, rats were individually housed and allowed to recover for one or two days.
Experimental Groups
A control group of six rats, used for EEG recording and histology, received a 2-µL intrahippocampal injection of Ringer solution. A second group composed of 11 rats received an intrahippocampal injection of 2 µg of TsTX diluted in 2 µL of Ringer solution. These animals were used for microdialysis experiments (five for amino acid test and six for analysis of monoamine) and histology. A third group of 11 rats received an IP injection of riluzole (5 mg/kg of body weight) and 15 minutes later a 2-µg intrahippocampal injection of TsTX. Six rats from this latter group were employed for EEG recording and histology, while the remainder were utilized for amino acid analysis.
Microdialysis
Microdialysis was performed in freely-moving rats between 24 and 48 hours after implantation of guide cannulas. The probe (CMA/11 microdialysis probe, membrane length 2.0 mm CMA Microdialysis Instruments, Sweden) was introduced into the guide cannula of each animal while the inlet was connected through 70-cm polyethylene tubing to a 2.5-mL Hamilton® syringe (Hamilton Company, USA). The outlet was connected to a 15-cm polyethylene tubing. Ringer solution was perfused at a flow rate of 2.13 µL/min and the final volume of samples was approximately 60 µL. After 30 to 45 minutes of equilibration period, the outflow was collected every 30 minutes during two hours. Subsequently, the probe was disconnected without flux interruption and the toxin was injected through the cannula. After that, the probe was connected again to each rat and twelve additional microdialysis samples were collected until the end of the experiment. Pretreated animals received an IP injection of riluzole 15 minutes before the toxin administration, without disconnecting probes. At the end of the collection, samples were dried and frozen (80° C) until the analysis. For biogenic amine determination, samples were collected in a tube with 5.0 µL of a solution composed of EDTA, sodium metabisulfite and perchloric acid.
Measurement of Neurotransmitter Concentration
Extracellular neurotransmitter levels in the dialysate were expressed as µg/µL or µg/mL. The first four samples from each animal were used as control and compared with post-treatment samples of the same individual.
Statistical significance was evaluated by one-way repeated measures analysis of variance and Tukey's post-hoc test. Values of p < 0.05 were considered significant.
Monoamines
Levels of dopamine (DA) and its metabolites (DOPAC and HVA), 5-HT and its metabolite (5-HIAA), and norepinephrine (NE) were measured by a HPLC system (model 6A, Shimadzu, Japan) with a C-18 Shim pack-ODS® column (Shimadzu, Japan), an electrochemical detector (model 6A, Shimadzu, Japan), a sample injector (valve for 20 #1), and an integrator (model 6A Chromatopac®, Shimadzu, Japan). Dihydroxybenzylamine (DHBA) was used as the internal standard. The limit of detection was 0.002 ng for DA, DOPAC, NE, 5-HT and 5-HIAA, and 0.02 ng for HVA.
Amino Acids
All samples were analyzed for amino acid content through HPLC along with ultraviolet (UV) detection after precolumn phenylisothiocyanate derivatization. The mobile phase consisted of solution A (19 g sodium acetate, 0.5 mL triethylamine and 200 µL of 1 M EDTA; for 470 mL of this solution, 30 mL acetonitrile was added), and solution B (400 mL acetonitrile, 100 mL water and 100 µL of 1 M EDTA). The gradient profile was determined to clearly identify each amino acid sample by comparison with the retention time of an amino acid standard solution containing 1 mM glutamate, 1 mM γ-aminobutyric acid (GABA) and 1 mM glycine.
The sample was mixed with 50 µL phenylisothiocyanate, and distilled water was added to a 350-µL volume. Levels of glutamate, glycine and GABA from the dialysate were determined by means of HPLC with UV detection at 254 nm.
Electroencephalographic Recordings and Behavioral Observations
Electroencephalographic recordings and behavioral observations were carried out in a glass compartment placed in a Faraday cage. Animals were connected to a PowerLab® recording apparatus (ADInstruments, Canada) and allowed to settle down for 15 minutes. Subsequently, basal electroencephalographic trace was recorded for 15 minutes. In rats treated with riluzole, the drug basal trace was analyzed for additional 15 minutes after its injection. Afterwards, an intracerebral injection of the toxin was administered through a 5 µL Hamilton® microsyringe (Hamilton Company, USA) connected with polyethylene tubing to an injection needle. Then, EEG was recorded and the behavior was observed for extra four hours.
Histology
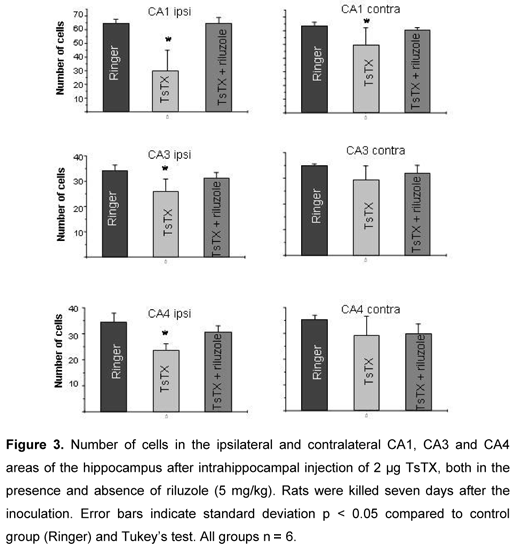
The location of implanted electrodes and guide cannulas was histologically examined. Animals that presented missing target areas were excluded. Seven days after the injection, animals were completely anesthetized with CO2 and received a cardiac perfusion (left ventricle) with phosphate-buffered saline (PBS) solution followed by 10% formalin solution. The brains were removed, stored in formalin for at least one week, and embedded in Paraplast® (Oxford Labware, USA). Coronal brain sections of 10 µm were cut on microtome from a 700 µm brain block containing the cannula track. Every sixth tissue slice was mounted on a glass slide and stained with cresyl violet. And the other five slices from each animal had their hippocampal field analyzed. The number of cells in the CA1, CA3 and CA4 hippocampal areas was examined through a light microscope at 400x magnification. A two-dimensional cell counting was performed using a 100 x 100 µm reticulum. Only pyramidal neurons located in the area of the reticulum that had a visible nucleus and nucleolus were considered intact. ANOVA followed by Tukey's test were employed for statistical analysis (p < 0.05).
RESULTS
Monoamine Levels
TsTX did not significantly affect levels of homovanillic acid (HVA), 3,4-dihydroxyphenylacetic acid (DOPAC) and 5-hydroxyindoleacetic acid (5-HIAA) (Figure 1). Levels of norepinephrine and its metabolites, serotonin and dopamine were not detected by our HPLC.
Extracellular Amino Acid Levels
Injection of TsTX toxin into the hippocampus augmented glutamate concentration in extracellular fluid (Figure 2), which reached statistical significance three hours after the injection. TsTX did not significantly affect levels of GABA and glycine. Pretreatment with riluzole (5 mg/kg body weight, IP at t = 15 min) prevented TsTX effect on hippocampal glutamate level (Figure 2).
Electroencephalographic Recordings and Behavioral Observations
Ringer solution injection into CA1 hippocampal area caused no alterations in behavior or in electroencephalographic records. Results of TsTX intrahippocampal injection were previously described and included long electroencephalographic epileptic-like discharges, immobility, orofacial movements and "wet dog shakes" in all animals (14). The effects started five to ten minutes after the injection and persisted for the entire recording period.
Rats treated with riluzole or with riluzole and toxin did not show epileptiform activity in electroencephalographic records or behavioral signs of convulsion.
Histology
Injection of Ringer solution into the hippocampus did not influence the number of cells in this area neither in the ipsilateral or contralateral regions (Figure 3). According to previous descriptions, outcomes provoked by intracerebral injection of the toxin were degeneration of pyramidal cells in ipsilateral and contralateral CA1 regions, and in CA3 and CA4 fields of the ipsilateral hippocampus (11, 14). Many dark-stained neurons were observed, which indicated the degeneration. Pretreatment with riluzole completely prevented toxin-induced cell loss. The number of cells in the ipsilateral and contralateral areas in these animals was similar to the control group (Figure 3).
DISCUSSION
Among scorpion neurotoxins, the α-group is the most studied and the more useful in the functional mapping of the sodium channel structure (18).
TsTX is an α-toxin that acts by enhancing neurotransmitter release in the central and peripheral nervous system (19, 20). Some studies have demonstrated that this toxin releases glutamate, acetylcholine and adenosine triphosphate (ATP) from rat cortical synaptosomes, and GABA and dopamine from cortical slices (10, 13, 21-24). Moreover, in vivo studies reveal that it induces electroencephalographic and histopathological alterations (14).
The present work aimed to assess the profile of in vivo neurotransmitter release in the hippocampus after TsTX injection. Based on the toxin kinetics and on the glutamate release profile registered in a previous study, in the current study the collection period was prolonged up to six hours after the injection and other neurotransmitters were analyzed (11, 15). It was observed that the toxin injected into the hippocampus did not modify extracellular levels of 5-HIAA, HVA, DOPAC, glycine or GABA. Nevertheless, extracellular levels of glutamate were significantly increased. These findings agree with Nencioni et al. (11) who observed enhanced levels of glutamate for three hours after the toxin injection and no alterations in GABA or glycine levels.
According to Nunan et al. (15), TsTX can be found in rat brain after subcutaneous (SC) injections. In adult animals the toxin increase is smaller and slower than in young ones. The highest brain concentration of the toxin was observed one hour after the inoculation in young rats and three hours later in adult ones, in which the levels were augmented up to 12 hours. These results were in accordance with ours, which revealed continuous enhancing in glutamate release that persisted until the end of the collection and probably after (six hours or more). Moreover, considering that in our study the toxin was injected directly into the brain, there was a large quantity of glutamate in the hippocampus.
Similar results had been obtained by Dorce and Sandoval (25) with crude venom, when DA, HVA, NE and 5-HIAA levels as well as glutamic acid decarboxylase (GAD) activity were determined after intravenous (IV) or intrastriatal injection of Tityus serrulatus crude venom. Subsequently, an increase in HVA levels and a decrease in GAD activity in the striatum and hypothalamus were observed. On the other hand, no alteration was registered in neurotransmitter levels in the hippocampus.
Previous data provided evidences that glutamate is the main responsible for TsTX neurotoxic effects since glutamate receptor antagonists totally or partially block the toxin effects (11). This hypothesis was corroborated by the utilization of riluzole, a substance that belongs to a class of anticonvulsant and neuroprotective agents which selectively inhibits glutamate release over release of other neurotransmitters (26, 27). Zona et al. (28) concluded that riluzole modulates Na+ currents and the late K+ current in cortical neurons; both phenomena are compatible with the anticonvulsant and neuroprotective properties of this compound. A decrease of the voltage-activated Na+ current certainly reduces neuronal excitability and enhances excitatory amino acid release (28). Under experimental conditions, the drug prevented audiogenic convulsion in DBA/2 mice, maximal electroshock and amygdala-kindled seizures in rats, as well as delayed the appearing of seizures and reduced the duration of afterdischarges (29, 30).
Riluzole injection 15 minutes before the toxin inoculation abolished all electrographic alterations as well as neuronal losses. Riluzole, when intraperitoneally injected, achieves the entire brain, including ipsi and contralateral areas of hippocampus, preventing the excitotoxic effects of the toxin. Animals did not show behavioral alterations either. They slept throughout the collection period, which is consistent with the finding that riluzole easily crosses the blood brain barrier and enhances slow-wave and rapid eye movement sleep (31). The level of extracellular glutamate remained elevated and reached statistical significance in 2 points in rats pretreated with riluzole; however, it is not enough to provoke epileptiform activity or neuronal loss.
According to Meldrum (32), enlarged release of glutamate may worsen or prolong preexisting seizure activity, while GABA release may be a compensatory inhibitory mechanism that limits the progression and spread of seizure activity (33). However, hippocampal augmentation of extracellular glutamate and GABA levels are directly related to seizure activity rather than to the convulsant agent (34). Meurs et al. (34) described that maximal glutamate increases occurred in the early stages of seizure development, and preceded maximal GABA elevation. Nevertheless, in our results, the electrographic convulsive activity did not keep a relation with the maximal extracellular glutamate level. Seizure activity onset occurred (approximately 15 minutes after toxin injection) before glutamate extracellular level reach a significant increase (approximately three hours after the injection), indicating that it is not necessary the maximal increase in glutamate extracellular levels to initiate convulsions. The convulsive activity enhanced by TsTX is not so intense to increase GABA levels. Rats treated with riluzole and TsTX maintained elevated levels of glutamate, although, they did not present behavioral or electrographic signs of convulsion neither neuronal loss. The glutamate level in these animals was not as high as in the group treated only with the toxin. It appears that riluzole can block convulsions and neurotoxicity elicited by TsTX without completely revert this increase.
In hippocampal neurons, voltage-gated sodium channels appear to be the primary, if not the only, target responsible for synaptic effects of riluzole at low micromolar concentrations. Riluzole's voltage-dependent blockade of sodium channels enhances the depression of excitatory postsynaptic currents, which does not apply to inhibitory postsynaptic currents determined by GABA. Inhibition of sodium channels account for the preferentially anti-glutamate effect of this drug. Therefore, the preferential depression of glutamate release is explained by a direct effect of riluzole on glutamatergic cells (35).
These findings constitute an evidence of a putative specific activity of TsTX, at least in the hippocampus, that affects only glutamate release.
In our opinion, outcomes of TsTX in the hippocampus may be due to its interaction with a specific subtype of sodium channel. Different actions on sodium channels produce diverse consequences on neurotransmitter release that involve distinct presynaptic calcium channels, which supports the idea that sodium channels may modulate neurotransmitter release (22). Depending on the rate of increase in channel conductance, the outcome in terms of neurotransmitter release and calcium channel type coupled to that event is different (36).
A large number of biological toxins exert their effects by modifying sodium channels properties (37). Gilles et al. (38) elucidated, for the first time, how different toxins affect mammalian central and peripheral excitable cells. They interact selectively with sodium channel subtypes in a discrete subcellular region. The study by Gilles et al. (39) revealed unexpected subtype specificity of toxins that interact with receptor site 3 (α-type toxins). Additionally, it was found that multiple sodium channel subtypes in mammalian brain can be pharmacologically distinguished by their sensitivity into certain toxins, such as ScαTxs (scorpion α-toxins) and αLTxs (spider α-latrotoxins) (39).
Numerous different isoforms of mammalian voltage-gated sodium channels have been identified (37). These multiple subtypes of sodium channels are the products of at least six distinct genes that differ in subcellular localization, developmental pattern of expression, and abundance in different brain regions (40-48).
Scorpion α-toxins provide a unique instrument for the identification of sodium channel subtypes due to their affinity for different channel subtypes (47). Thus, TsTX may be an important tool to study the functional role and distribution of channel subtypes in the hippocampus.
Received: July 4, 2008
Accepted: August 26, 2008
Abstract published online: September 18, 2008
Full paper published online: May 30, 2009
CONFLICTS OF INTEREST: There is no conflict
- 1. Rodriguez de la Vega RC, Possani LD. Overview of scorpion toxins specific for Na+ channels and related peptides: biodiversity, structure-function relationships and evolution. Toxicon. 2005;46(8):831-44
- 2. Rodriguez de la Vega RC, Possani LD. Current views on scorpion toxins specific for K+-channels. Toxicon. 2004;43(8):865-75.
- 3. Debin JA, Maggio JE, Strichartz GR. Purification and characterization of chlorotoxin, a chloride channel ligand from the venom of the scorpion. Am J Physiol. 1993;264(2):361-9.
- 4. Valdivia HH, Kirby MS, Lederer WJ, Coronado R. Scorpion toxins targeted against the sarcoplasmic reticulum calcium-release channel of skeletal and cardiac muscle. Proc Natl Acad Sci USA. 1992;89(24):12185-9.
- 5. Cestele S, Catterall WA. Molecular mechanisms of neurotoxin action on voltage-gated sodium channels. Biochimie. 2000;82(9-10):883-92.
- 6. Kirsch GE, Skattebol A, Possani LD, Brown AM. Modification of Na+ channel gating by a α-scorpion toxin from Tityus serrulatus J Gen Physiol. 1989;93(1):67-83.
- 7. Massensini AR, Romano-Silva MA, Gomez MV. Sodium channel toxins and neurotransmitter release. Neurochem Res. 2003;28(10):1607-11.
- 8. Gomez MV, Diniz CR. Separation of toxic components from the Brazilian scorpion Tityus serrulatus venom. Mem Inst Butantan. 1966;33(3):899-902.
- 9. Arantes EC, Sampaio SV, Vieira CA, Giglio JR. What is tityustoxin? Toxicon. 1992;30(7):786-9.
- 10. Massensini AR, Moraes-Santos T, Gomez MV, Romano-Silva MA. Alpha- and beta-scorpion toxins evoke glutamate release from rat cortical synaptosomes with different effects on [ Na+] i and [ Ca2+] i. Neuropharmacology. 1998;37(3):289-97.
- 11. Nencioni AL, Lebrun I, Dorce VA. A microdialysis study of glutamate concentration in the hippocampus of rats after TsTX toxin injection and blockade of toxin effects by glutamate receptor antagonists. Pharmacol Biochem Behav. 2003;74(2):455-63.
- 12. Casali TAA, Gomez RS, Moraes-Santos T, Gomez MV. Differential effects of calcium channel antagonists on tityustoxin and ouabain-induced release of [ 3H] acethylcholine from brain cortical slices. Neuropharmacology. 1995;34(6):599-603.
- 13. Nicolato R, Fernandes VMV, Moraes-Santos T, Gomez RS, Prado MAM, Romano-Silva MA, Gomez MV. Release of γ[ 3H] aminobutiric acid in rat brain cortical slices by α-scorpion toxin. Neurosci Lett. 2002;325(3):155-8.
- 14. Carvalho FF, Nencioni AL, Lebrun I, Sandoval MR, Dorce VA. Behavioral, electroencephalographic and histopathologic effects of a neuropeptide isolated from Tityus serrulatus scorpion venom in rats. Pharmacol Biochem Behav. 1998;60(1):7-14.
- 15. Nunan EA, Moraes MF, Cardoso VN, Moraes-Santos T. Effects of age on body distribution of tityustoxin from Tityus serrulatus scorpion venom in rats. Life Sci. 2003;73(3):319-25.
- 16. Kretschmer BD, Kratzer U, Schmidt WJ. Riluzole, a glutamate release inhibitor, and motor behavior. Naunyn Schmiedebergs Arch Pharmacol. 1998;358(2):181-90.
- 17. Paxinos G, Watson C. The rat brain in stereotaxic coordinates. 4th ed. San Diego: Academic Press; 1998.
- 18. Bosmans F, Tytgat J. Voltage-gated sodium channel modulation by scorpion alpha-toxins. Toxicon. 2007;49(2):142-58.
- 19. Gomez MV, Romano-Silva MA, Prado MAM. Effects of tityustoxin on central nervous system. Toxin Rev. 1995;14(3):437-56.
- 20. Lima EG, Freire-Maia L. Cardiovascular and respiratory effects induced by intracerebroventricular injection of scorpion toxin (tityustoxin) in the rat. Toxicon. 1977;15(3):225-34.
- 21. Fletcher PL, Fletcher M, Fainter LK, Terrian DM. Action of new world scorpion venom and its neurotoxins in secretion. Toxicon. 1996;34(11-12):1399-411.
- 22. Falqueto EB, Massensini AR, Moraes-Santos T, Gomez MV, Romano-Silva MA. Modulation of Na+-channels by neurotoxins produces different effects on [ 3H] ACh release with mobilization of distinct Ca2+-channels. Cell Mol Neurobiol. 2002;22(5-6):819-26.
- 23. Salgado AH, Prado MA, Moraes-Santos T, Romano-Silva MA, Gomez MV. Tityustoxin-induced release of ATP from rat brain cortical synaptosomes. Neurosci Lett. 1997;229(2):113-6.
- 24. Fernandes VM, Massensini AR, Prado MA, Silva MA, Moraes-Santos T, Gomez MV. Effects of α-scorpion toxin, tityustoxin on the release of [ 3H] dopamine of rat brain prefrontal cortical slices. Neurochem Int. 2004;44(2):91-7.
- 25. Dorce VA, Sandoval MR. Effects of Tityus serrulatus crude venom on the GABAergic and dopaminergic systems on the rat brain. Toxicon. 1994;32(12):1641-7.
- 26. Maciver MB, Amagasu SM, Mikulec AA, Monroe FA. Riluzole anesthesia: use-dependent block of presynaptic glutamate fibers. Anesthesiology. 1996;85(3):626-34.
- 27. Martin D, Thompson MA, Nadler JV. The neuroprotective agent riluzole inhibits release of glutamate and aspartate from slices of hippocampal area CA1. Eur J Pharmacol. 1993;250(3):473-6.
- 28. Zona C, Siniscalchi A, Mercuri NB, Bernardi G. Riluzole interacts with voltage-activated sodium and potassium currents in cultured rat cortical neurons. Neuroscience. 1998;85(3):931-8.
- 29. De Sarro G, Siniscalchi A, Ferreri G, Gallelli L, De Sarro A. NMDA and AMPA/kainate receptors are involved in the anticonvulsant activity of riluzole in DBA/2 mice. Eur J Pharmacol. 2000;408(1):25-34.
- 30. Yoshida M, Noguchi E, Tsuru N, Ohkoshi N. Effect of riluzole on the acquisition and expression of amygdala kindling. Epilepsy Res. 2001;46(2):101-9.
- 31. Wokke J. Riluzole. Lancet. 1996;348(9030):795-9.
- 32. Meldrum BS. The role of glutamate in epilepsy and other CNS disorders. Neurology. 1994;44(11 Suppl 8):14-23.
- 33. Treiman DM. GABAergic mechanisms in epilepsy. Epilepsia. 2001;42(Suppl 3):8-12.
- 34. Meurs A, Clinckers R, Ebinger G, Michotte Y, Smolders I. Seizure activity and changes in hippocampal extracellular glutamate, GABA, dopamine and serotonin. Epilepsy Res. 2008;78(1):50-9.
- 35. Prakriya M, Mennerick S. Selective depression of low-release probability excitatory synapses by sodium channel blockers. Neuron. 2000;26(3):671-82.
- 36. Bicalho AF, Guatimosim C, Prado MA, Gomez MV, Romano-Silva MA. Investigation of the modulation of glutamate release by sodium channels using neurotoxins. Neuroscience. 2002;113(1):115-23.
- 37. Catterall WA, Cestele S, Yarov-Yarovoy V, Yu FH, Konoki K, Scheuer T. Voltage-gated ion channels and gating modifier toxins. Toxicon. 2007;49(2):124-41.
- 38. Gilles N, Chen H, Wilson H, Le Gall F, Montoya G, Molgo J, Schönherr R, Nicholson G, Heinemann SH, Gordon D. Scorpion α and α-like toxins differentially interact with sodium channels in mammalian CNS and periphery. Eur J Neurosci. 2000;12(8):2823-32.
- 39. Gilles N, Blanchet C, Shichor I, Zaninetti M, Lotan I, Bertrand D, Gordon D. A scorpion α-like toxin that is active on insects and mammals reveals an unexpected specificity and distribution of sodium channel subtypes in rat brain neurons. J. Neurosci. 1999;19(20):8730-9.
- 40. Schaller KL, Krzemien DM, Yarowsky PJ, Krueger BK, Caldwell JH. A novel, abundant sodium channel expressed in neurons and glia. J Neurosci. 1995;15(5 Pt 1):3231-42.
- 41. Sangameswaran L, Fish LM, Koch BD, Rabert DK, Delgado SG, Ilnicka M, Jakeman LB, Novakovic S, Wong K, Sze P, Tzoumaka E, Stewart GR, Herman RC, Chan H, Eglen RM, Hunter JC. A novel tetrodotoxin-sensitive voltage-gated sodium channel expressed in rat and human dorsal root ganglia. J Biol Chem. 1997;272(23):14805-9.
- 42. Toledo-Aral JJ, Moss BL, He ZJ, Koszowski AG, Whisenand T, Levinson SR, Wolf JJ, Silos-Santiago I, Halegoua S, Mandel G. Identification of PN1, a predominant voltage-dependent sodium channel expressed principally in peripheral neurons. Proc Natl Acad Sci USA. 1997;94(4):1527-32.
- 43. Westenbroeck RE, Merrick DK, Catterall WA. Differential subcellular localization of the RI and RII Na+ channel subtypes in the central neurons. Neuron. 1989;3(6):695-704.
- 44. Black JA, Yokoyama S, Higashida H, Ransom Br, Waxman Sg. Sodium channel mRNAs I, II and III in the CNS: cell-specific expression. Brain Res Mol Brain Res. 1994;22(1-4):275-89.
- 45. Denac H, Mevissen M, Scholtysik G. Structure, function and pharmacology of voltage-gated sodium channels. Naunyn Schmiedebergs Arch Pharmacol. 2000;362(6):453-79.
- 46. Felts PA, Yokoyama S, Dib-Hajj S, Black JA, Waxman SG. Sodium channel α-subunit mRNAs I, II, III, NaG, Na6 and hNE (PNI): different expression patterns in developing rat nervous system. Mol Brain Res. 1997;45(1):71-82.
- 47. Gordon D, Gurevitz M. The selectivity of scorpion α-toxins for sodium channel subtypes is determined by subtle variations at the interacting surface. Toxicon. 2003; 41(2):125-8.
- 48. Gordon D, Merrick D, Auld V, Dunn R, Goldin AL, Davidson N, Catterall W. Tissue-specific expression of RI and RII sodium channel subtypes. Proc Natl Acad Sci USA. 1987;84(23):8682-6.
Correspondence to:
Publication Dates
-
Publication in this collection
09 June 2009 -
Date of issue
2009
History
-
Received
04 July 2008 -
Accepted
26 Aug 2008