Abstract
Background:
The Asiatic pit vipers from the Trimeresurus complex are medically important venomous snakes. These pit vipers are often associated with snakebite that leads to fatal coagulopathy and tissue necrosis. The cytotoxic venoms of Trimeresurus spp.; however, hold great potential for the development of peptide-based anticancer drugs.
Methods:
This study investigated the cytotoxic effect of the venom from Trimeresurus purpureomaculatus, the mangrove pit viper (also known as shore pit viper) which is native in Malaysia, across a panel of human cancer cell lines from breast, lung, colon and prostate as well as the corresponding normal cell lines of each tissue.
Results:
The venom exhibited dose-dependent cytotoxic activities on all cell lines tested, with median inhibition concentrations (IC50) ranging from 0.42 to 6.98 µg/mL. The venom has a high selectivity index (SI = 14.54) on breast cancer cell line (MCF7), indicating that it is significantly more cytotoxic toward the cancer than to normal cell lines. Furthermore, the venom was fractionated using C18 reversed-phase high-performance liquid chromatography and the anticancer effect of each protein fraction was examined. Fraction 1 that contains a hydrophilic low molecular weight (approximately 7.5 kDa) protein was found to be the most cytotoxic and selective toward the breast cancer cell line (MCF7). The protein was identified using liquid chromatography-tandem mass spectrometry as a venom disintegrin, termed purpureomaculin in this study.
Conclusion:
Taken together, the findings revealed the potent and selective cytotoxicity of a disintegrin protein isolated from the Malaysian T. purpureomaculatus venom and suggested its anticancer potential in drug discovery.
Keywords:
Shore pit viper;
Trimeresurus purpureomaculatus
; Disintegrin; Selective cytotoxicity; Anti-neoplastic activity
Background
Snake venom is widely regarded as an advanced “biochemical weapon” that exists in nature. Venomous snakes rely on venom for survival, where it is employed for predation (primary function), digestion and defense [11. Casewell NR, Wuster W, Vonk FJ, Harrison RA, Fry BG. Complex cocktails: the evolutionary novelty of venoms. Trends in ecology & evolution. 2013;28(4):219-29. Epub 2012/12/12. doi: 10.1016/j.tree.2012.10.020. PubMed PMID: 23219381.
https://doi.org/10.1016/j.tree.2012.10.0...
]. Rapid evolution drives the emergence of diverse pharmacologically active snake venom proteins that are adapted to target the normal physiology of prey by exerting various toxic effects [22. Ferraz CR, Arrahman A, Xie C, Casewell NR, Lewis RJ, Kool J, et al. Multifunctional toxins in snake venoms and therapeutic implications: From pain to hemorrhage and necrosis. Frontiers in ecology and evolution. 2019;7(218). doi: 10.3389/fevo.2019.00218.
https://doi.org/10.3389/fevo.2019.00218...
]. The prey of venomous snakes consists in parts of small mammals such as rodents but may also include birds, lizards and amphibians [33. Das I. A field guide to the reptiles of South-East Asia: Bloomsbury Publishing; 2015., 44. Marlon R. 107+ ular Indonesia: panduan visual dan identifikasi lapangan. 2014.]. It is noteworthy that the mammalian system-targeting property of snake venom proteins is often specific and selective, thus making them a natural repertoire of therapeutic molecules [55. Mohamed Abd El-Aziz T, Garcia Soares A, Stockand JD. Snake venoms in drug discovery: Valuable therapeutic tools for life saving. Toxins. 2019;11(10):564. doi: 10.3390/toxins11100564. PubMed PMID: 31557973.
https://doi.org/10.3390/toxins11100564...
, 66. Munawar A, Ali S, Akrem A, Betzel C. Snake venom peptides: Tools of biodiscovery. Toxins. 2018;10(11):474.].
At present, advanced cancers that are fast-growing and capable of metastasizing is a major cause of mortality globally [77. Siegel RL, Miller KD, Jemal A. Cancer Statistics, 2017. CA Cancer J Clin. 2017;67(1):7-30. doi: 10.3322/caac.21387. PubMed PMID: 28055103.
https://doi.org/10.3322/caac.21387...
, 88. Ferlay J, Soerjomataram I, Dikshit R, Eser S, Mathers C, Rebelo M, et al. Cancer incidence and mortality worldwide: sources, methods and major patterns in GLOBOCAN 2012. International journal of cancer. 2015;136(5):E359-86. Epub 2014/09/16. doi: 10.1002/ijc.29210. PubMed PMID: 25220842.
https://doi.org/10.1002/ijc.29210...
]. In this context, snake venom proteins have the potential for drug discovery in line with the search for novel peptide-based anticancer agents with high efficacy and selectivity in targeting cancer cells [99. Li L, Huang J, Lin YH. Snake venoms in cancer therapy: Past, present and future. Toxins. 2018;10(9):346. doi: 10.3390/toxins10090346. PubMed PMID: 30158426.
https://doi.org/10.3390/toxins10090346...
, 1010. Boohaker RJ, Lee MW, Vishnubhotla P, Perez JM, Khaled AR. The use of therapeutic peptides to target and to kill cancer cells. Current medicinal chemistry. 2012;19(22):3794-804. Epub 2012/06/26. doi: 10.2174/092986712801661004. PubMed PMID: 22725698; PubMed Central PMCID: PMCPmc4537071.
https://doi.org/10.2174/0929867128016610...
]. Notable examples typically involved venom proteins from vipers and pit vipers (family Viperidae), e.g. disintegrin from Agkistrodon contortrix venom [1111. Trikha M, De Clerck YA, Markland FS. Contortrostatin, a snake venom disintegrin, inhibits beta 1 integrin-mediated human metastatic melanoma cell adhesion and blocks experimental metastasis. Cancer research. 1994;54(18):4993-8. Epub 1994/09/15. PubMed PMID: 7520832.], phospholipases A2 from Cerastes cerastes [1212. Zouari-Kessentini R, Srairi-Abid N, Bazaa A, El Ayeb M, Luis J, Marrakchi N. Antitumoral potential of Tunisian snake venoms secreted phospholipases A2. Biomed Res Int. 2013;2013:391389. Epub 2013/03/20. doi: 10.1155/2013/391389. PubMed PMID: 23509718; PubMed Central PMCID: PMCPMC3581298.
https://doi.org/10.1155/2013/391389...
] and L-amino acid oxidases from Bothrops sp. [1313. Costa TR, Burin SM, Menaldo DL, de Castro FA, Sampaio SV. Snake venom L-amino acid oxidases: an overview on their antitumor effects. J Venom Anim Toxins Incl Trop Dis. 2014;20:23. Epub 2014/06/19. doi: 10.1186/1678-9199-20-23. PubMed PMID: 24940304; PubMed Central PMCID: PMCPMC4060840.
https://doi.org/10.1186/1678-9199-20-23...
].
A common pharmacological feature of the venoms of viperid snakes, including the Asian arboreal pit vipers from the Trimeresurus complex, is their ability to derange hemostasis and induce local tissue necrosis [1414. Mong R, Tan HH. Snakebite by the shore pit viper (Trimeresurus purpureomaculatus) treated with polyvalent antivenom. Wilderness & environmental medicine. 2016;27(2):266-70. Epub 2016/04/12. doi: 10.1016/j.wem.2016.01.001. PubMed PMID: 27061038.
https://doi.org/10.1016/j.wem.2016.01.00...
-1616. Chan T, Hung LK. Digital gangrene following a green pit viper bite. The Southeast Asian journal of tropical medicine and public health. 2010;41(1):192-4. Epub 2010/06/29. PubMed PMID: 20578498.]. The tissue-necrotizing effect associated with Trimeresurus pit viper envenomation suggests cytotoxic activity in the venom that may be further explored for anticancer potential. A recent study with Thai Trimeresurus purpureomaculatus venom reported high cytotoxicity of the venom toward SHSY5Y cell line, and moderate cytotoxicity of BPP-related peptides, PLA2 and peptide-rich fraction of the venom [1717. Ozverel CS, Damm M, Hempel B-F, Göçmen B, Sroka R, Süssmuth RD, et al. Investigating the cytotoxic effects of the venom proteome of two species of the Viperidae family (Cerastes cerastes and Cryptelytrops purpureomaculatus) from various habitats. Comparative Biochemistry and Physiology part C: Toxicology & Pharmacology. 2019;220:20-30. doi: https://doi.org/10.1016/j.cbpc.2019.02.013.
https://doi.org/10.1016/j.cbpc.2019.02.0...
]. Another publication reported that L-amino acid oxidase (LAAO, 55−60 kDa) from Malaysian Trimeresurus purpureomaculatus venom induced cytotoxicity in three colon cell lines (SW480, SW620 and CCD-18Co), but no selectivity was observed between the cancerous cell lines (SW480, SW620) and the non-cancerous, normal cell line (CCD-18Co) [1818. Zainal Abidin SA, Rajadurai P, Hoque Chowdhury ME, Othman I, Naidu R. Cytotoxic, anti-proliferative and apoptosis activity of l-amino acid oxidase from Malaysian Cryptelytrops purpureomaculatus (CP-LAAO) venom on human colon cancer cells. Molecules. 2018;23(6).].
Our recent quantitative proteomic analysis of the Malaysian T. purpureomaculatus venom (MTP) revealed that LAAO constituted about 3% of the total venom proteins, while disintegrin, a compound with potential inhibitory effect on cancer cell growth, was present at a higher abundance in the venom (approximately 14%) [1919. Liew JL, Tan NH, Tan CH. Proteomics and preclinical antivenom neutralization of the mangrove pit viper (Trimeresurus purpureomaculatus, Malaysia) and white-lipped green pit viper (Trimeresurus albolabris, Thailand) venoms. Acta tropica. 2020;209:105528. doi: https://doi.org/10.1016/j.actatropica.2020.105528.
https://doi.org/10.1016/j.actatropica.20...
]. The LAAO content reported previously for the Malaysian T. purpureomaculatus venom could not be compared as it was estimated with a qualitative method [2020. Zainal Abidin SA, Rajadurai P, Chowdhury ME, Ahmad Rusmili MR, Othman I, Naidu R. Proteomic characterization and comparison of Malaysian Tropidolaemus wagleri and Cryptelytrops purpureomaculatus venom using shotgun-proteomics. Toxins 2016;8(10). Epub 2016/10/21. doi: 10.3390/toxins8100299. PubMed PMID: 27763534; PubMed Central PMCID: PMCPmc5086659.
https://doi.org/10.3390/toxins8100299...
]. In a more recent study, relative protein abundances of 0.91% LAAO and 0.10% disintegrin were identified in the Thai T. purpureomaculatus venom [1717. Ozverel CS, Damm M, Hempel B-F, Göçmen B, Sroka R, Süssmuth RD, et al. Investigating the cytotoxic effects of the venom proteome of two species of the Viperidae family (Cerastes cerastes and Cryptelytrops purpureomaculatus) from various habitats. Comparative Biochemistry and Physiology part C: Toxicology & Pharmacology. 2019;220:20-30. doi: https://doi.org/10.1016/j.cbpc.2019.02.013.
https://doi.org/10.1016/j.cbpc.2019.02.0...
].
Disintegrin is a much smaller (4-15 kDa, monomer or dimer) cysteine-rich and RGD-containing polypeptide that targets specifically on integrin - a ubiquitously expressed cell surface receptor that regulates cell motility, survival, proliferation, angiogenesis and cell invasion [2121. Arruda Macêdo JK, Fox JW, de Souza Castro M. Disintegrins from snake venoms and their applications in cancer research and therapy. Current protein & peptide science. 2015;16(6):532-48. doi: 10.2174/1389203716666150515125002. PubMed PMID: 26031306.
https://doi.org/10.2174/1389203716666150...
, 2222. Rivas-Mercado EA, Garza-Ocañas L. Disintegrins obtained from snake venom and their pharmacological potential. University medicine magazine. 2017;19(74):32-7. doi: https://doi.org/10.1016/j.rmu.2017.02.004.
https://doi.org/10.1016/j.rmu.2017.02.00...
]. The antineoplastic potential of snake venom disintegrins have been shown in several studies, e.g. contortrostatin (disintegrin isolated from Agkistrodon contortrix venom) [2323. Swenson S, Costa F, Ernst W, Fujii G, Markland FS. Contortrostatin, a snake venom disintegrin with anti-angiogenic and anti-tumor activity. Pathophysiology of haemostasis and thrombosis. 2005;34(4-5):169-76. Epub 2006/05/19. doi: 10.1159/000092418. PubMed PMID: 16707922.
https://doi.org/10.1159/000092418...
], saxatilin (disintegrin from Gloydius saxatilis venom) [2424. Hong SY, Koh YS, Chung KH, Kim DS. Snake venom disintegrin, saxatilin, inhibits platelet aggregation, human umbilical vein endothelial cell proliferation, and smooth muscle cell migration. Thrombosis research. 2002;105(1):79-86. Epub 2002/02/28. PubMed PMID: 11864711.], PAIEM (disintegrin isolated from Echis multisquamatis venom) [2525. Chernyshenko V, Petruk N, Korolova D, Kasatkina L, Gornytska O, Platonova T, et al. Antiplatelet and antiproliferative action of disintegrin from Echis multisquamatis snake venom. Croatian medical journal. 2017;58(2):118-27.] and a disintegrin from Crotalus durissus collilineatus venom [2626. de Oliveira IS, Manzini RV, Ferreira IG, Cardoso IA, Bordon KdCF, Machado ART, et al. Cell migration inhibition activity of a non-RGD disintegrin from Crotalus durissus collilineatus venom. Journal of venomous animals and toxins including tropical diseases. 2018;24(1):28.]. The disintegrins act by inhibiting angiogenesis, invasion and migration of cancer cells. The finding of disintegrin in the Malaysian T. purpureomaculatus venom is significant as the peptide could be a promising local source of anticancer candidate that has not been characterized previously.
In this study, the Malaysian T. purpureomaculatus venom was suggested to be cytotoxic to a wider range of cancer cell lines including those of the human breast, lung, colon and prostate. The selective anticancer activity, if any, could be partly contributed by the disintegrin present in the venom. Hence, this study aimed to investigate the cytotoxicity of Malaysian T. purpureomaculatus venom across four common human cancerous and the corresponding normal cell lines. In addition, the disintegrin was purified from the venom, and its amino acid sequence as well as anticancer properties were characterized.
Methods
Venom Sample
The venom of mangrove pit viper (Trimeresurus purpureomaculatus, MTP) was a pooled sample from ten adult snake specimens (four males; six females) from Peninsular Malaysia. The venom obtained was freeze-dried and stored at −20 °C until use. Venom stock used in each cytotoxic experiment was prepared freshly from lyophilized venom and subjected to a quick spin for 15 seconds prior to the treatment.
Chemicals and Materials
All chemicals and reagents used in the studies were of analytical grade. 3,(4,5-dimethythiazol-2-yl)-2,3-diphenyl tetrazolium bromide (MTT) was purchased from Sigma Aldrich (Missouri, USA) and dimethyl sulfoxide (DMSO) was supplied by Merck (Darmstadt, Germany). Trypsin-EDTA was supplied by Nacalai Tesque (Kyoto, Japan). The positive control in the MTT assay was 5-fluorouracil (5-FU) (Sigma-Aldrich, Missouri, USA). The reversed-phase HPLC column LiChrospher® WP 300 RP-18 (5 μm) was purchased from Merck (Darmstadt, Germany) and HPLC solvents were from (Thermo Scientific, Massachusetts, USA). Trifluoroacetic acid (TFA) was purchased from Sigma Aldrich (Missouri, USA). Ammonium bicarbonate, dithiothreitol (DTT) and iodoacetamide (IAA), which were used in protein digestion, were purchased from Sigma-Aldrich (Missouri, USA), Trypsin protease (MS grade) was purchased from Thermo Scientific Pierce™ (Massachusetts, USA) and desalting C18 pipette tips was purchased from Merck Millipore ZipTip® (MilliporeSigma, Massachusetts, USA). Protein ladder of PM2700 ExcelBand™ 3-color Broad Range Protein Marker was purchased from SMOBIO Technology, Inc. (Hsinchu, Taiwan).
Cell Culture
Both normal and cancer cell lines were obtained from American Type Culture Collection (ATCC, Virginia, USA). MCF7 (ATCC® HTB-22™; human breast adenocarcinoma cell line), HT-29 (ATCC® HTB-38™; human colon colorectal adenocarcinoma cell line), A549 (ATCC® CCL-185™; human lung carcinoma cell line) and PC-3 (ATCC® CRL-1435™; human prostate adenocarcinoma cell line) were used as the cancer cell panel, while 184B5 (ATCC® CRL-8799™; human breast normal cell line), CCD-18Co (ATCC® CRL-1459™; human colon normal cell line), MRC5 (ATCC® CCL-171™; human lung normal cell line) and RWPE-1 (ATCC® CRL-11609™; human prostate normal cell line) were used as the normal cell panel in the cytotoxic assay.
MCF7 and HT-29 cell lines were cultured in Dulbecco’s modified Eagle’s medium (DMEM) (Nacalai Tesque, Kyoto, Japan), supplemented with 10% fetal bovine serum (FBS) (TICO Europe, Amstelveen, Netherlands) and 100 µg/mL penicillin-streptomycin (Nacalai Tesque, Kyoto, Japan), while A549 and PC-3 were maintained using RPMI 1640 medium with L-glutamine (Lonza, Verviers, Belgium), with 10% FBS and 100 µg/mL penicillin-streptomycin. Normal cell lines such as CCD-18Co and MRC5 were maintained in Eagle’s minimum essential medium (EMEM) (Sigma-Aldrich, Missouri, USA), with 10% FBS and 100 µg/mL penicillin-streptomycin. The 184B5 was maintained in mammary epithelial cell growth medium (MEGM) with bullet kit (Lonza, Basel, Switzerland), supplemented with 10% FBS and 100 µg/mL penicillin-streptomycin, while RWPE-1 was maintained in Keratinocyte-SFM (serum-free medium) (Thermo and Scientific, Massachusetts, USA), with only 100 µg/mL penicillin-streptomycin added to the medium. The cell lines were cultured in a 5% carbon dioxide (CO2) incubator (Shel Lab, Oregon, USA) at 37 ºC, with pH of 7.2 - 7.5 and relative humidity of about 95%. The growth of cells was monitored routinely by observing cell morphology under an inverted microscope (Leica Microsystems, Wetzlar, Germany). All of the cell-related work was carried out using aseptic techniques under sterile condition.
Cell Viability Assay
Cell viability was studied with the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay. The cells of the respective cell lines were detached from the culture flasks using an appropriate amount of trypsin-EDTA and desired number of cells were seeded in a 96-well microplate (15,000-150,000 cells/mL) for overnight incubation. The attached cells were then treated with serial dilutions (ranging from 0.10-31.62 μg/mL) of T. purpureomaculatus venom for 72 hours. After treatment, 10% of MTT reagent was added into each well without removing the previous medium. The plate was covered with aluminum foil and returned to the CO2 incubator for 3 hours incubation. The MTT solution was pipetted out gently after the incubation and 200 μL of dimethyl sulfoxide (DMSO) was added into each well to solubilize the formazan crystal. Absorbance was measured using Hidex Plate CHAMELEON™V (Hidex, Turku, Finland) multilabel microplate reader at 570 nm. The median inhibition concentration (IC50) was determined from the dose-response curve plotted with percentage of cell viability against venom concentration (μg/mL). In the assay, 5-fluorouracil (5-FU) was used as a positive reference. The percentage of cell viability was calculated with the following formula:
Selectivity Index Determination
The degrees of selective cytotoxicity of the venom in cancer cell lines were indicated with selectivity index (SI), determined as follows:
Fractionation of T. purpureomaculatus Venom and Bioassay-Guided Cytotoxicity Study
Two hundred microliters of T. purpureomaculatus venom solution (10 mg/mL in ultrapure water) were injected into LiChrosper® WP 300-RP-18 reversed-phase column (5 μm column particle size) through the Shimadzu LC-20AD high performance liquid chromatography system. The column was pre-equilibrated with solvent B [0.1% trifluoroacetic acid (TFA) in acetonitrile] followed by solvent A (0.1% TFA in water). The elution began with the stepwise linear gradient (0-5% of B for 10 min, followed by 5-15% B for 20 min, 15-60% B for the next 180 minutes, 60-70% B for 10 minutes and 75-100% of B over 245 min) in 0.1% TFA in acetonitrile (ACN) for 245 minutes (flow rate: 1 mL/min). Protein elution was monitored at 215 nm.
The protein fractions were collected manually, subsequently freeze-dried and stored at −20 °C until use. Each protein fraction was reconstituted in ultrapure water and the protein concentration was estimated by Nanodrop Spectrophotometer 2000 (ThermoFisher™, Massachusetts, USA) prior to the experiment. The cytotoxic fraction of T. purpureomaculatus venom was screened with a bioassay-guided method modified from Shahbazi [2727. Shahbazi B, Najafabadi ZS, Goudarzi H, Sajadi M, Tahoori F, Bagheri M. Cytotoxic effects of Pseudocerastes persicus venom and its HPLC fractions on lung cancer cells. Journal of venomous animals and toxins including tropical diseases. 2019;25.]. The cytotoxic activities of the venom fraction were tested at a standard dose of 20 µg each on human breast cancer cell line (MCF7) according to the cell viability assay described above. The amount of protein from each venom fraction tested (20 µg) was referred from a previous study by Bradshaw [2828. Bradshaw MJ, Saviola AJ, Fesler E, Mackessy SP. Evaluation of cytotoxic activities of snake venoms toward breast (MCF-7) and skin cancer (A-375) cell lines. Cytotechnology. 2016;68(4):687-700. Epub 2014/11/20. doi: 10.1007/s10616-014-9820-2. PubMed PMID: 25407733; PubMed Central PMCID: PMCPmc4960119.
https://doi.org/10.1007/s10616-014-9820-...
]. Cell images were captured 72 hours after treatment using inverted microscope (Leica Microsystems, Wetzlar, Germany).
Sodium Dodecyl Sulphate-Polyacrylamide Gel Electrophoresis (SDS-PAGE)
Gel electrophoresis was carried out according to the protocol of Laemmli [2929. Laemmli UK. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature. 1970;227(5259):680.]. Approximately 10 μg of protein fraction was loaded onto a 15% acrylamide gel and the electrophoresis was carried out under reducing conditions at 90 V for 2 hours. Protein ladder PM2700 ExcelBand™ 3-color Broad Range Protein Marker was used as molecular weight standards in the electrophoresis (5-245 kDa). Coomassie Brilliant Blue R-250 staining was used for the visualization of protein.
Protein Digestion and Mass Spectrometry Analysis of the Cytotoxic-Contributing Fraction of MTP Venom
The most cytotoxic protein fraction of MTP venom was subjected to in-solution tryptic digestion and nano-ESI LC-MS/MS analysis as previously reported [3030. Tan CH, Tan KY, Ng TS, Quah ESH, Ismail AK, Khomvilai S, et al. Venomics of Trimeresurus (Popeia) nebularis, the Cameron Highlands pit viper from Malaysia: Insights into venom proteome, toxicity and neutralization of antivenom. Toxins. 2019;11(2):95.]. Ten micrograms of the protein (10 µL of 1 mg/mL protein concentration) first underwent reduction and alkylation by addition of 15 μL of 50 mM ammonium bicarbonate and 1.5 μL of 100 mM dithiothreitol (DTT). The mixture was heated at 95 ºC for 5 min before addition of 3 μL of 100 mM iodoacetamide (IAA). The mixture was later incubated in dark at room temperature for another 20 min. Digestion was carried out by adding one microliter of 0.1 mg/mL trypsin solution to the reaction tube and incubated at 37 ºC for 3 hours. Additional 1 µL of trypsin solution was added for overnight incubation at 30 ºC. After digestion, the digested peptides were desalted for removal of salts and contaminants.
The desalted peptides (approximately 10 µg) were then reconstituted in 7 µL of 0.1% formic acid in water. One microliter of the solution (containing approximately 1.4 µg of peptides) was subjected to nano-electrospray ionization MS/MS via Agilent 1260 HPLC-Chip/MS Interface coupled with Agilent 6550 Accurate-Mass Q-TOF LC/MS system (Agilent Technologies, Santa Clara, California, USA). The sample peptides were separated in a large capacity chip Zorbax 300 Å, C18, 160 nl enrichment column, 75 μm × 150 mm analytical column and 5 μm particles (Agilent part no. G4240-62010). Parameters were set as follows: injection volume at 1 µL per sample, flow rate from capillary pump at 4 µL/min and 0.4 µL/min from Nano pump (G2226A), gradient used: 5-50% solution B (0.1% formic acid in acetonitrile) for 11 min, 50-70% B for next 4 min, and 70% B for 3 min and ion polarity was set to positive ionization mode. Drying gas flow and temperature were set at 11 L/min and 290 ºC. Fragmentor voltage was set at 175 V while for capillary voltage, it was set at 1800 V. MS scan range of 200-3000 m/z and MS/MS scan range of 50-3200 m/z were acquired in the tandem mass spectrometry mode, with precursor charge selection set as doubly charge state and above with the exclusion of precursor 1221.9906 m/z (z = 1) for internal mass calibration and reference ions set at 299.2944 (z = 1). Data with a MH+ mass range between 50 and 3200 were extracted and analyzed in Agilent Spectrum Mill MS Proteomics Workbench software packages (Agilent Technologies, Santa Clara, CA, USA). Setting for cysteine carbamidomethylation is set as a fixed modification, while methionine oxidation is set as variable modification.
The mass spectrometry derived peptide masses were searched against a non-redundant NCBI database of Serpentes (taxid: 8570) combined with the venom-gland transcriptome database of T. purpureomaculatus. The protein identification was validated by the following filters: protein score > 20, peptide score > 10 and score peak intensity (SPI) > 70%. Proteins with “Distinct Peptide” ≥ 2 were considered significant matches.
Bioinformatic Analyses
Sequence analysis of disintegrin
Multiple sequence alignment of disintegrins was performed using MUSCLE program and Jalview software v2.11 to indicate the region of similarity. The percentage of similarity was calculated using EMBL-EBI Clustal Omega (https://www.ebi.ac.uk/Tools/msa/clustalo/). The disintegrin sequence of Malaysian Trimeresurus purepureomaculatus (MTP) venom was derived from the venom-gland transcriptome (entry is available in NCBI GenBank database, accession ID: QJA41976.1). Other related disintegrin sequences were retrieved from Uniprot Knowledgebase (https://www.uniprot.org/) [3131. Consortium TU. UniProt: a worldwide hub of protein knowledge. Nucleic acids research. 2018;47(D1):D506-D15. doi: 10.1093/nar/gky1049.
https://doi.org/10.1093/nar/gky1049...
].
In silico physicochemical characterization of disintegrin
The physicochemical properties of the disintegrin from MTP venom were characterized using the Expasy ProtParam online tool (https://web.expasy.org/protparam/) according to Roly et al. [3232. Roly ZY, Islam MM, Reza MA. A comparative in silico characterization of functional and physicochemical properties of 3FTx (three finger toxin) proteins from four venomous snakes. Bioinformation. 2014;10(5):281-7. doi: 10.6026/97320630010281. PubMed PMID: 24966535.
https://doi.org/10.6026/97320630010281...
]. Parameters including theoretical molecular weight, isoelectric point (pI), instability index and grand average of hydropathicity (GRAVY) value were determined.
Statistical Analysis
The median inhibition concentration (IC50) value was determined using GraphPad Prism 5 statistical software (GraphPad Software Inc., California, USA) and the values were expressed as mean ± S.E.M. of three replicates. Comparative data were statistically analyzed using Student’s unpaired t-test (at 95% confidence interval) with GraphPad Prism 5 software (GraphPad Software Inc., California, USA).
Results
Cytotoxicity of Trimeresurus purpureomaculatus Venom
MTP venom exhibited dose-dependent cytotoxic effects toward all cancer cell lines tested (median inhibition concentrations, IC50 = 0.42−2.50 µg/mL). The effects were generally stronger in the cancer cells compared to the corresponding normal cells (IC50 = 0.70-6.98 µg/mL) (Table 1). The venom cytotoxicity was most potent to the colorectal adenocarcinoma cell line (IC50 = 0.42 ± 0.06 µg/mL), followed by the breast cancer cell line (IC50 = 0.48 ± 0.02 µg/mL) and prostate cancer cell line (IC50 = 1.60 ± 0.18 µg/mL). The venom was least cytotoxic to the lung cancer cell line (IC50 = 2.50 ± 0.20 µg/mL). The venom was also significantly more potent compared to 5-FU in inhibiting the growth of breast, lung and prostate cancer cells (p < 0.05). In comparison to the IC50 in normal cell lines, the cytotoxic effect was not selective toward colon, lung and prostate cancer cells as indicated by a selectivity index (SI) ≤ 10. The selectivity index of the venom was, however, much higher (SI = 14.54) for the breast cancer cell line (MCF7), implying that the venom was 15-fold more cytotoxic to MCF7 than it was to the corresponding normal cell line (184B5) (Table 1, Figure 1).

Dose-dependent growth inhibitory effect of Trimeresurus purpureomaculatus (MTP) venom in human breast cell lines. Median inhibition concentrations (IC50) were determined from the dose-response curve. Values were presented as means ± S.E.M. (n = 3). MCF7: human breast cancer cell line, 184B5: human breast normal cell line.
Cytotoxicity of the Chromatographic Fractions of Trimeresurus purpureomaculatus Venom
MTP venom was resolved using C18 reversed-phase high performance liquid chromatography (RP-HPLC) into 12 fractions as shown in Figure 2A. The cytotoxic effect of each fraction was further tested on MCF7 cell line in view of the high selectivity index (SI) of MTP venom in breast cancer cell line. MCF7 cells treated with protein from Fraction 1 (20 µg) showed the most potent inhibitory effect (cell viability reduced by 65%) compared to the non-treated control (**p < 0.01). Proteins from Fraction 2 and 12 exhibited moderate cytotoxic effect (cell viability reduced by 25−30%) whereas other fractions did not show significant cytotoxicity in the MCF7 cell line (Figure 2B). In comparison, 5-FU at IC50 of 9 µg/mL induced a 50% reduction in the cell viability study.

(A) C18 reversed-phase high performance liquid chromatography of venom from the Malaysian T. purpureomaculatus. (B) Cell viability of human breast cancer cells (MCF7) after 72-hour treatment with the HPLC fractions of Malaysian T. purpureomaculatus venom. Positive and negative controls were 5-fluorouracil (5-FU) and treatment-free, respectively, in the assay.
Identification and Cytotoxicity of Disintegrin from Trimeresurus purpureomaculatus Venom
Figure 3 shows the SDS-PAGE profile of the cytotoxic protein isolated in Fraction 1. A homogenous protein band was observed and the molecular mass was estimated to be 7585 Da by its relative migration distance (Rf). LC-MS/MS identified Fraction 1 to contain only one protein, which is a disintegrin of snake venom and it is termed as “purpureomaculin” in the present study. The mass spectrometry data file of purpureomaculin is available in Additional file 1
Additional file 1.
LC-MS/MS of purpureomaculin isolated from Malaysian Trimeresurus purpureomaculatus venom (Fraction 1)
, and the primary data was deposited to the ProteomeXchange Consortium via the iProX partner repository [3333. Ma J, Chen T, Wu S, Yang C, Bai M, Shu K, et al. iProX: an integrated proteome resource. Nucleic acids research. 2019;47(D1):D1211-D7. doi: 10.1093/nar/gky869. PubMed PMID: 30252093.
https://doi.org/10.1093/nar/gky869...
] with the dataset identifier PXD018463.

Protein content of Fraction 1 (F1) of Malaysian T. purpureomaculatus venom was validated under 15% reducing gel electrophoresis. Protein ladder PM2700 ExcelBand™ 3-color Broad Range Protein Marker was used for molecular weight standards (5−245 kDa).
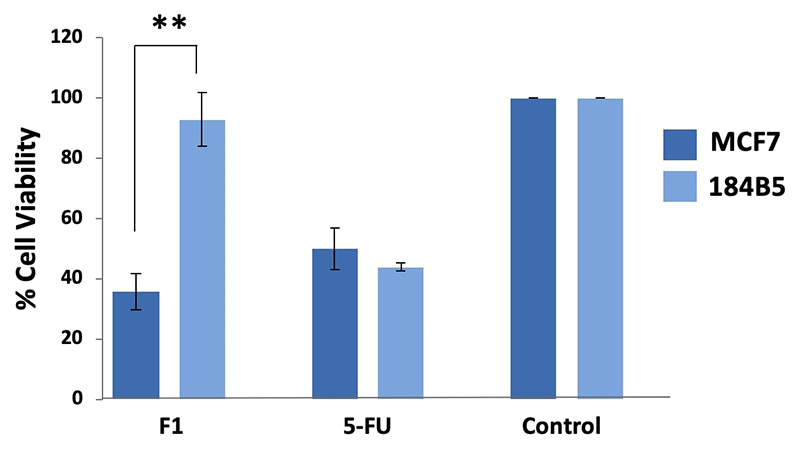
Purpureomaculin at 20 µg significantly inhibited the growth of MCF7 cell line by 65% (**p < 0.01), while no significant cytotoxic effect was observed in the corresponding normal breast cells (184B5 cell line) (Figure 4). Microscopic examination revealed the presence of detached, clumped and rounded floater cells in the purpureomaculin-treated breast cancer cells following a treatment period of 72 hours. On the contrary, the normal cell line (184B5) treated with purpureomaculin was observed to remain healthy and viable, with slight reduced growth compared to the untreated control (Figure 5).

Cell viability of human breast cell lines, MCF7 (cancerous) and 184B5 (normal) after 72-hour treatment with 20 µg of Fraction 1 (F1, purpureomaculin) purified from Trimeresurus purpureomaculatus venom. Positive control: 5-fluorouracil (5-FU); negative control: treatment-free well.

Morphological changes in human breast cell lines, (A) MCF7 (cancerous) and (B) 184B5 (normal) after 72-hour treatment of Fraction 1 (purpureomaculin) of Trimeresurus purpureomaculatus venom. Scale bars = 30 µm.
Sequence Analysis and in silico Characterization of Purpureomaculin, the Cytotoxic Disintegrin from T. purpureomaculatus Venom
The tryptic peptides of purpureomaculin (K)LLPGAQCGEGLCCDQCSFMKK(G) and (R)ARGDDLDDYCNGISAGCPR(N) from Fraction 1 were matched to the disintegrin transcript “CL53.Contig5_CP” from the Malaysian T. purpureomaculatus venom-gland transcriptomic database. A disintegrin with full amino acid sequence was identified. It is a mature chain polypeptide of 73 amino acid residues: EAGEDCDCGSPANPCCNAATCKLLPGAQCGEGLCCDQCSFMKKGTICRRARGDDLDDYCNGISAGCPRNPLHA.
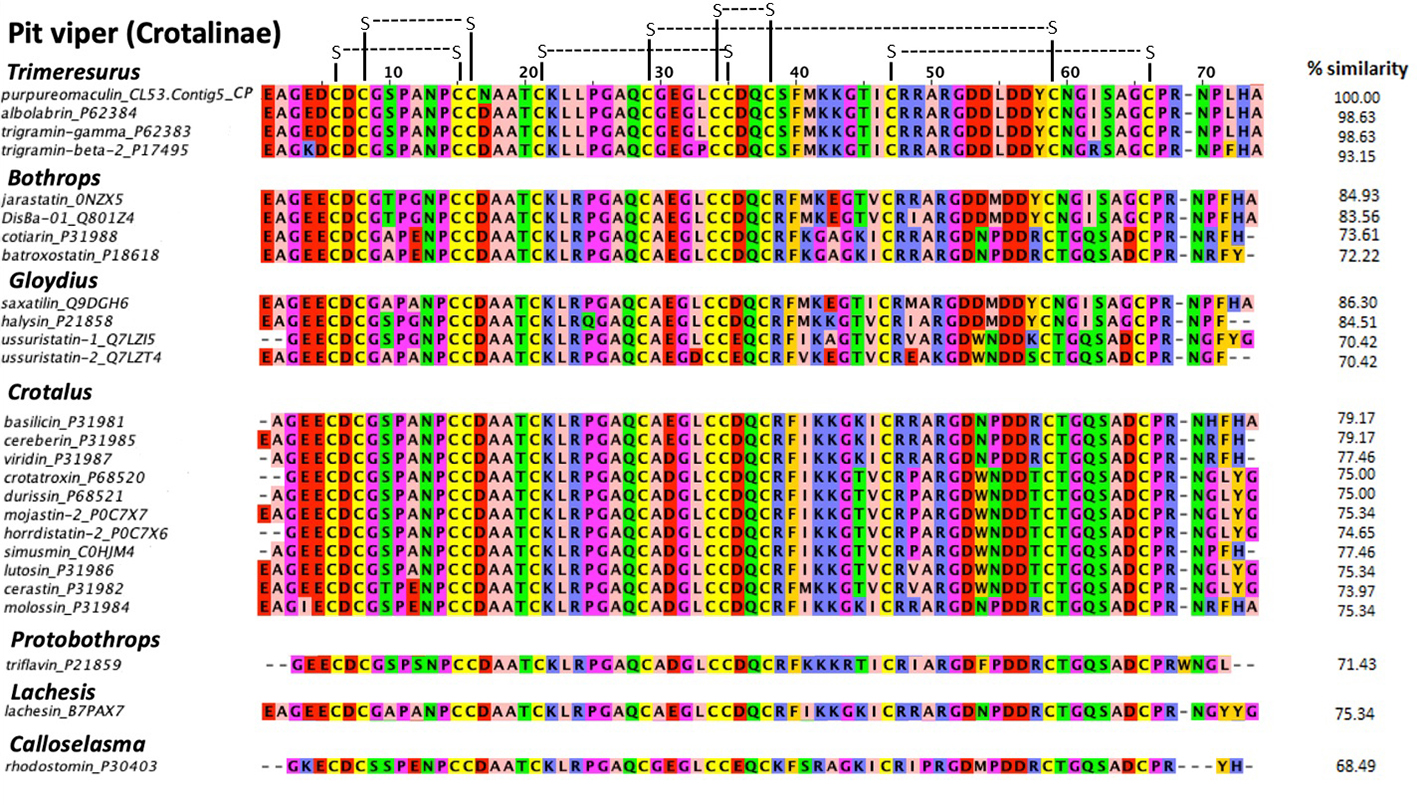
Multiple sequence alignment showed that purpureomaculin is highly homologous (98.6%) with albolabrin from Trimeresurus albolabris venom and trigramin-gamma from Trimeresurus gramineus venom, where purpureomaculin varies from albolabrin and trigramin-gamma by only one amino acid at the 17th position of the sequence (substitution of Asp to Asn in purpureomaculin) (Figure 6). Six disulfide bonds could be formed within the sequence, pairing cysteine residues at the following positions: (6, 15), (8, 16), (21, 35), (29, 59), (34, 38), (47, 66). Purpureomaculin also harbored the tripeptide RGD, a short sequence motif of biological interest formed by amino acid residues 51−53. Compared with the disintegrin sequences of other viperids, high similarity (> 70%) was observed in phylogenetically related pit viper genera and species, e.g. Trimeresurus complex.

Multiple sequence alignment of purpureomaculin (CL53.Contig5_CP) with other disintegrin sequences of Crotalinae. Purpureomaculin is a disintegrin isolated from the Malaysian Trimeresurus purpureomaculatus venom, while other disintegrin sequences were obtained from UniProtKB. The percentages of similarity (% similarity) of disintegrin sequences in comparison to purpureomaculin were calculated using Omega Clustal software. Zappo color scheme indicates residue properties: pink: aliphatic/hydrophobic; orange: aromatic; blue: positive; red: negative; green: hydrophilic; magenta: proline/glycine (conformationally special); yellow: cysteine. Disulfide bonds were illustrated in black connecting lines.
Table 2 shows the physicochemical properties of purpureomaculin analyzed in silico using the ProtParam Tool. Purpureomaculin has an estimated molecular weight of 7572.52 Da and is weakly acidic (pI = 4.79).
Discussion
In this study, the potent cytotoxicity of Malaysian T. purpureomaculatus venom was shown in a panel of cancer (MCF7, HT-29, PC3, A549) and normal (184B5, CCD-18Co, RWPE-1, MRC5) cell lines. MTP venom was particularly toxic to breast cancer cell line (MCF7) and colon cell lines (HT-29 and CCD-18Co), indicating high specificity of the venom toward these tested cells. The venom also demonstrated selective cytotoxicity, particularly in the human breast cancer cells (MCF7). This suggests that the venom contains toxin(s) or protein(s) that could be cancer-specific in breast tumor, thus potentiating the venom cytotoxicity at a lower dose in the cancer cells compared to the corresponding normal cells. The cytotoxic effect of Malaysian T. purpureomaculatus venom also appeared to be more potent than other pit viper venoms, including venoms of Protobothrops flavoviridis [3535. Damm M, Hempel B-F, Nalbantsoy A, Süssmuth RD. Comprehensive snake venomics of the Okinawa Habu pit viper, Protobothrops flavoviridis, by complementary mass spectrometry-guided approaches. Molecules. 2018;23(8):1893.], Trimeresurus macrops and Trimeresurus hageni [3636. Kumkate S, Chanhome L, Thiangtrongjit T, Noiphrom J, Laoungboa P, Khow O, et al. Venomics and cellular toxicity of Thai pit vipers (Trimeresurus macrops and T. hageni). Toxins 2020;12(1). Epub 2020/01/23. doi: 10.3390/toxins12010054. PubMed PMID: 31963345; PubMed Central PMCID: PMCPMC7020458.
https://doi.org/10.3390/toxins12010054...
], albeit different human cell lines were utilized in each study.
Further investigation revealed that most of the venom components (fractionated with HPLC) showed variable anticancer activity, implying that the overall venom cytotoxicity could be a result of synergistic interactions of multiple components in the venom. The most potent anti-neoplastic activity was found in the protein fraction containing snake venom disintegrin. This was a purified protein validated through SDS-PAGE and LC-MS/MS, and termed purpureomaculin in line with the naming of most snake venom-derived disintegrin protein (prefix “species-” + post-fix “-in”). The finding of purpureomaculin anticancer activity is consistent with earlier studies that reported the inhibitory activity of snake venom disintegrins on the proliferation, metastasis and adhesion of cancer cells [2121. Arruda Macêdo JK, Fox JW, de Souza Castro M. Disintegrins from snake venoms and their applications in cancer research and therapy. Current protein & peptide science. 2015;16(6):532-48. doi: 10.2174/1389203716666150515125002. PubMed PMID: 26031306.
https://doi.org/10.2174/1389203716666150...
, 2525. Chernyshenko V, Petruk N, Korolova D, Kasatkina L, Gornytska O, Platonova T, et al. Antiplatelet and antiproliferative action of disintegrin from Echis multisquamatis snake venom. Croatian medical journal. 2017;58(2):118-27., 3737. Hammouda MB, Montenegro MF, Sánchez-del-Campo L, Zakraoui O, Aloui Z, Riahi-Chebbi I, et al. Lebein, a snake venom disintegrin, induces apoptosis in human melanoma cells. Toxins. 2016;8(7):206.]. The selective cytotoxic effect of purpureomaculin in breast cancer cells (MCF7) corroborates the selective cytotoxicity of MTP venom in the same cancer cell line observed in this study.
The present findings varied when compared with previous studies on other cancer cells using T. purpureomaculatus venoms from Malaysia and Thailand [1717. Ozverel CS, Damm M, Hempel B-F, Göçmen B, Sroka R, Süssmuth RD, et al. Investigating the cytotoxic effects of the venom proteome of two species of the Viperidae family (Cerastes cerastes and Cryptelytrops purpureomaculatus) from various habitats. Comparative Biochemistry and Physiology part C: Toxicology & Pharmacology. 2019;220:20-30. doi: https://doi.org/10.1016/j.cbpc.2019.02.013.
https://doi.org/10.1016/j.cbpc.2019.02.0...
, 1818. Zainal Abidin SA, Rajadurai P, Hoque Chowdhury ME, Othman I, Naidu R. Cytotoxic, anti-proliferative and apoptosis activity of l-amino acid oxidase from Malaysian Cryptelytrops purpureomaculatus (CP-LAAO) venom on human colon cancer cells. Molecules. 2018;23(6).]. IC50 values with wide discrepancy were observed between previous studies (Malaysian T. purpureomaculatus IC50 (72 hours): 15.99−29.43 µg/mL in human colon cell lines [1818. Zainal Abidin SA, Rajadurai P, Hoque Chowdhury ME, Othman I, Naidu R. Cytotoxic, anti-proliferative and apoptosis activity of l-amino acid oxidase from Malaysian Cryptelytrops purpureomaculatus (CP-LAAO) venom on human colon cancer cells. Molecules. 2018;23(6).]; Thai T. purpureomaculatus (IC50 (48 hours): 0.25−5.24 µg/mL in human neuroblastoma, cervical, colon, breast, bladder, glioblastoma and kidney cell lines [17]) and the present work (Malaysian T. purpureomaculatus: IC50 (72 hours): 0.42-6.98 µg/mL in breast, lung, colon and prostate cell lines) (Table 3). Nevertheless, the cytotoxic activity of the present Malaysian T. purpureomaculatus venom in human colon cells appeared to be comparable to the Thai venom sample reported by Ozverel [1717. Ozverel CS, Damm M, Hempel B-F, Göçmen B, Sroka R, Süssmuth RD, et al. Investigating the cytotoxic effects of the venom proteome of two species of the Viperidae family (Cerastes cerastes and Cryptelytrops purpureomaculatus) from various habitats. Comparative Biochemistry and Physiology part C: Toxicology & Pharmacology. 2019;220:20-30. doi: https://doi.org/10.1016/j.cbpc.2019.02.013.
https://doi.org/10.1016/j.cbpc.2019.02.0...
]. In contrast, much higher IC50 values (by a difference of > 10 folds) in human colon cells were reported previously in the other Malaysian T. purpureomaculatus venom sample [1818. Zainal Abidin SA, Rajadurai P, Hoque Chowdhury ME, Othman I, Naidu R. Cytotoxic, anti-proliferative and apoptosis activity of l-amino acid oxidase from Malaysian Cryptelytrops purpureomaculatus (CP-LAAO) venom on human colon cancer cells. Molecules. 2018;23(6).]. The discrepancy could be due to the variation in venom composition, the use of different colon cell lines, or the influence of the condition of venom preparation. In our experience, repeated freeze-thaw cycles of the MTP venom led to inconsistent and somewhat deteriorating venom cytotoxic activity (unpublished); hence, all venom samples used in the present study were freshly reconstituted from lyophilized stock sample prior to treating the cells in the MTT experiment. The additional measure, presumably, avoided any possible protein degradation [3838. Mills JB, Mant CT, Hodges RS. One-step purification of a recombinant protein from a whole cell extract by reversed-phase high-performance liquid chromatography. Journal of chromatography A. 2006;1133(1-2):248-53. Epub 09/01. doi: 10.1016/j.chroma.2006.08.042. PubMed PMID: 16945380.
https://doi.org/10.1016/j.chroma.2006.08...
] or inactivation of the venom bioactive components by unfavorable pH or temperature [3939. Kang TS, Georgieva D, Genov N, Murakami MT, Sinha M, Kumar RP, et al. Enzymatic toxins from snake venom: structural characterization and mechanism of catalysis. The FEBS journal. 2011;278(23):4544-76. Epub 2011/04/08. doi: 10.1111/j.1742-4658.2011.08115.x. PubMed PMID: 21470368.
https://doi.org/10.1111/j.1742-4658.2011...
].
In addition, previous studies had demonstrated variable anticancer activities for venom disintegrin from pit vipers. For instance, lebein (disintegrin isolated from Macrovipera lebetina venom) has been shown to significantly reduce the cell viability of colon adenocarcinoma cell lines after 72-hour incubation [4040. Zakraoui O, Marcinkiewicz C, Aloui Z, Othman H, Grepin R, Haoues M, et al. Lebein, a snake venom disintegrin, suppresses human colon cancer cells proliferation and tumor-induced angiogenesis through cell cycle arrest, apoptosis induction and inhibition of VEGF expression. Molecular carcinogenesis. 2017;56(1):18-35. Epub 2016/01/30. doi: 10.1002/mc.22470. PubMed PMID: 26824338.
https://doi.org/10.1002/mc.22470...
]. A more recent study demonstrated that disintegrins isolated from Cerastes cerastes venom displayed varying IC50 values (1.60−8.17 µg/mL) in SHSY5Y neuroblastoma cell line [1717. Ozverel CS, Damm M, Hempel B-F, Göçmen B, Sroka R, Süssmuth RD, et al. Investigating the cytotoxic effects of the venom proteome of two species of the Viperidae family (Cerastes cerastes and Cryptelytrops purpureomaculatus) from various habitats. Comparative Biochemistry and Physiology part C: Toxicology & Pharmacology. 2019;220:20-30. doi: https://doi.org/10.1016/j.cbpc.2019.02.013.
https://doi.org/10.1016/j.cbpc.2019.02.0...
]. Tzabcanin (disintegrin isolated from Crotalus simus tzabcan venom) exhibited low cytotoxicity against Colo-205 cells and was not cytotoxic to MCF7 cells [4141. Saviola AJ, Modahl CM, Mackessy SP. Disintegrins of Crotalus simus tzabcan venom: Isolation, characterization and evaluation of the cytotoxic and anti-adhesion activities of tzabcanin, a new RGD disintegrin. Biochimie. 2015;116:92-102.]. On the contrary, the disintegrin purpureomaculin of T. purpureomaculatus was found to be potent in inhibiting the growth of MCF7 cells (current study). The anticancer activities of disintegrin from many closely related species such as trigramin (disintegrin isolated from Trimeresurus gramineus venom) and albolabrin (disintegrin isolated from Trimeresurus albolabris venom) remain unknown to date, and warrant investigation for insights in the anticancer potential of the venom peptide from this large Asiatic pit viper complex.
The multiple sequence alignment revealed that the venom disintegrins of pit vipers (Crotalinae) are mainly of intermediate length with 12 conserved cysteine residues, forming 6 disulfide bonds that support the folding and stability of the protein [4242. Calvete JJ, Schaefer W, Soszka T, Lu W, Cook JJ, Jameson BA, et al. Identification of the disulfide bond pattern in albolabrin, an RGD-containing peptide from the venom of Trimeresurus albolabris: Significance for the express of platelet aggregation inhibitory activity. Biochemistry. 1991;30(21):5225-9.]. Disintegrins from Trimeresurus, Bothrops, Gloydius, Crotalus, Protobothrops and Lachesis complexes in particular showed a higher rate of amino acid conservation, indicating the phylogenetic relationship among different genera within the Crotalinae subfamily. At the intra-generic level of Trimeresurus, purpureomaculin sequence was found almost identical (similarity: 98.6%) to albolabrin and trigramin-gamma (disintegrins from Trimeresurus albolabris and Trimeresurus gramineus). Like albolabrin and trigramin, purpureomaculin is structurally categorized as the medium-sized disintegrins which contain about 70 amino acids and 6 disulfide cysteine bonds, as described by Calvete [4242. Calvete JJ, Schaefer W, Soszka T, Lu W, Cook JJ, Jameson BA, et al. Identification of the disulfide bond pattern in albolabrin, an RGD-containing peptide from the venom of Trimeresurus albolabris: Significance for the express of platelet aggregation inhibitory activity. Biochemistry. 1991;30(21):5225-9.]. The high similarity of purpureomaculin to these disintegrins indicates structural and functional resemblance between these disintegrins, and that purpureomaculin is likely to exert its cytotoxic activity similar to trigramin and albolabrin by inhibiting cell-matrix adhesion through the binding of RGD tripeptide domain to the cellular integrin receptors [4343. Knudsen KA, Tuszynski GP, Huang T-F, Niewiarowski S. Trigramin, an RGD-containing peptide from snake venom, inhibits cell-substratum adhesion of human melanoma cells. Experimental cell research. 1988;179(1):42-9. doi: https://doi.org/10.1016/0014-4827(88)90346-1.
https://doi.org/10.1016/0014-4827(88)903...
, 4444. Soszka T, Knudsen KA, Beviglia L, Rossi C, Poggi A, Niewiarowski S. Inhibition of murine melanoma cell-matrix adhesion and experimental metastasis by albolabrin, an RGD-containing peptide isolated from the venom of Trimeresurus albolabris. Experimental cell research. 1991;196(1):6-12. Epub 1991/09/01. doi: 10.1016/0014-4827(91)90449-5. PubMed PMID: 1879472.
https://doi.org/10.1016/0014-4827(91)904...
]. One remarkable difference of purpureomaculin is the amino acid substitution at the 17th position. It is noted that amino acid at this position is highly conserved throughout the pit viper genera, and the substitution of the aspartic acid (D) to the neutral asparagine (N) in purpureomaculin may have a unique evolutionary implication on the role of the protein expressed. The consequence of the amino acid substitution deserves further investigation.
The sequence similarities between purpureomaculin and other disintegrin sequences are lower (69−86%) when comparing with venom disintegrins of different genera other than Trimeresurus. All pit viper disintegrin sequences (including purpureomaculin) contains RGD-flanking motif, a tripeptide domain known to bind to a wide range of integrin receptors. In particular, disintegrins with RGD tripeptides were known to bind key integrin receptors such as α2β1 [4545. Eble JA, Niland S, Dennes A, Schmidt-Hederich A, Bruckner P, Brunner G. Rhodocetin antagonizes stromal tumor invasion in vitro and other alpha2beta1 integrin-mediated cell functions. Journal of the international society for matrix biology. 2002;21(7):547-58. Epub 2002/12/12. PubMed PMID: 12475639.], α4β1 [4646. Danen EHJ, Marcinkiewicz C, Cornelissen IMHA, van Kraats AA, Pachter JA, Ruiter DJ, et al. The disintegrin eristostatin interferes with integrin α4β1 function and with experimental metastasis of human melanoma cells. Experimental cell research. 1998;238(1):188-96. doi: https://doi.org/10.1006/excr.1997.3821.
https://doi.org/10.1006/excr.1997.3821...
], αvβ3 [4747. Chung KH, Kim SH, Han Ky, Sohn YD, Chang SI, Baek KH, et al. Inhibitory effect of salmosin, a Korean snake venom derived disintegrin, on the integrin αv‐mediated proliferation of SK‐Mel‐2 human melanoma cells. Journal of pharmacy and pharmacology. 2003;55(11):1577-82., 4848. Ramos OH, Kauskot A, Cominetti MR, Bechyne I, Salla Pontes CL, Chareyre F, et al. A novel alpha(v)beta (3)-blocking disintegrin containing the RGD motive, DisBa-01, inhibits bFGF-induced angiogenesis and melanoma metastasis. Clinical & experimental metastasis. 2008;25(1):53-64. Epub 2007/10/24. doi: 10.1007/s10585-007-9101-y. PubMed PMID: 17952617.
https://doi.org/10.1007/s10585-007-9101-...
] and αvβ5 [4949. Zhou Q, Nakada MT, Brooks PC, Swenson SD, Ritter MR, Argounova S, et al. Contortrostatin, a homodimeric disintegrin, binds to integrin alphavbeta5. Biochemical and biophysical research communications. 2000;267(1):350-5. Epub 2000/01/07. doi: 10.1006/bbrc.1999.1965. PubMed PMID: 10623623.
https://doi.org/10.1006/bbrc.1999.1965...
], leading to inhibition of integrin-mediated functions in cancer cells. Among these, the high expression of integrins αvβ3 and αvβ5 are commonly associated with tumorigenesis [5050. Bianconi D, Unseld M, Prager GW. Integrins in the spotlight of cancer. Int J Mol Sci. 2016;17(12):2037. doi: 10.3390/ijms17122037. PubMed PMID: 27929432.
https://doi.org/10.3390/ijms17122037...
]. A distinct feature of MCF7 cell line is that it lacks integrin αvβ3 receptor, but showed hyperexpression of integrins αvβ5 receptor [5151. Taherian A, Li X, Liu Y, Haas T. Differences in integrin expression and signaling within human breast cancer cells. BMC cancer. 2011;11:293. doi: 10.1186/1471-2407-11-293.
https://doi.org/10.1186/1471-2407-11-293...
]. We hypothesized that the selective cytotoxicity of purpureomaculin is at least in part mediated through the binding of integrin receptors (e.g. αvβ5 receptor) that leads to the inhibition of cancer cell adhesion, angiogenesis and metastasis. This explains the microscopic observation in the present study, where purpureomaculin caused significant detachment of the treated breast cancer cells (MCF7), forming round-shape floaters while sparing the architecture of the normal breast cells (184B5).
In addition, the primary sequence of purpureomaculin was analyzed in silico to further understand the physicochemical properties of the identified disintegrin. Purpureomaculin was computed to have a molecular weight of 7572.5 Da, similar to that determined from the reducing SDS-PAGE of the protein (7585 Da). The molecular mass determined, however, may be slightly varied from that examined by intact mass profiling using a top-down mass spectrometry analysis. The intact mass profiling of purpureomaculin hence should be carried out in the future to establish the exact mass. On the other hand, the computed instability index (II) that falls below 40 indicates that purpureomaculin is theoretically stable in vitro. The instability index could be applied to estimate the in vivo half-life of a protein [5252. Guruprasad K, Reddy BB, Pandit MW. Correlation between stability of a protein and its dipeptide composition: a novel approach for predicting in vivo stability of a protein from its primary sequence. Protein Eng. 1990;4(2):155-61.]: Proteins whose II < 40 have an in vivo half-life beyond 16 h, whereas proteins whose II > 40 are usually eliminated faster from the body with an in vivo half-life of less than 5 h [5353. Rogers S, Wells R, Rechsteiner M. Amino acid sequences common to rapidly degraded proteins: the PEST hypothesis. Science. 1986;234(4774):364-8. Epub 1986/10/17. doi: 10.1126/science.2876518. PubMed PMID: 2876518.
https://doi.org/10.1126/science.2876518...
]. The II value obtained for purpureomaculin in this work was about 25, predicting a long in vivo half-life beyond 16 h for this disintegrin. The aliphatic index (AI) represents the relative volume of aliphatic side chains (alanine, valine, isoleucine and leucine) in the amino acid sequence of a protein. The AI value computed for purpureomaculin reflects high protein thermostability. On the other hand, the GRAVY parameter predicts for the feature hydrophobic (positive values) or hydrophilic (negative values) of a protein. Purpureomaculin has a negative value of GRAVY, implying that it is more of a hydrophilic protein - this is also consistent with the early elution of this protein from the reversed-phase HPLC. Together, the physicochemical parameters predict that purpureomaculin is a relatively stable protein with an extended in vivo half-life, and readily dissolves in a polar solvent. Moreover, the small molecular size of the disintegrin peptide (7572.52 Da) implies that it is likely less antigenic, more accessible to cancer cell environment and amenable to structural modification e.g. nano-carrier conjugation in the advanced development of peptide-based anticancer agent [5454. Mishra J, Panda JJ. Short peptide-based smart targeted cancer nanotherapeutics: a glimmer of hope. Therapeutic delivery. 2019;10(3):135-8. Epub 2019/03/27. doi: 10.4155/tde-2019-0005. PubMed PMID: 30909857.
https://doi.org/10.4155/tde-2019-0005...
, 5555. Bhawani SA, Husaini A, Ahmad FB, Asaruddin MR. Polymer based protein therapeutics. Current protein & peptide science. 2018;19(10):972-82. Epub 2017/08/23. doi: 10.2174/1389203718666170821162823. PubMed PMID: 28828988.
https://doi.org/10.2174/1389203718666170...
].
Conclusion
The present study demonstrated the potent cytotoxicity of the Malaysian T. purpureomaculatus venom and unveiled its selective anticancer activity in the human breast cancer cell line (MCF7). Purpureomaculin, a disintegrin identified from the Malaysian T. purpureomaculatus venom, was found to be most cytotoxic and selective among several protein fractions of the venom, presumably due to its inhibitory action on cell-cell adhesion. In silico characterization of purpureomaculin sequence predicted a stable, hydrophilic small polypeptide molecule, with features that may favorably unleash its pharmaceutical potential in the development of a peptide-based anticancer therapeutic. Nevertheless, further studies are required to investigate the peptide delivery system and immunogenicity of the peptide, which are the main obstacles of most current peptide-based therapeutics.
Abbreviations
5-FU: 5-fluorouracil; ACN: acetonitrile; ATCC: American Type Culture Collection; DMEM: Dulbecco’s modified Eagle’s medium; DMSO: dimethyl sulfoxide; DTT: dithiothreitol; EMEM: Eagle’s minimum essential médium; FBS: fetal bovine serum; IAA: iodoacetamide; IC50: median inhibition concentration; LAAO: L-amino acid oxidase; MEGM: mammary epithelial cell growth medium; MTP: Malaysian T. purpureomaculatus; pI: isoelectric point; RP-HPLC: reversed-phase high performance liquid chromatography; SI: selectivity index; SPI: score peak intensity; TFA: trifluoroacetic acid.
References
- 1. Casewell NR, Wuster W, Vonk FJ, Harrison RA, Fry BG. Complex cocktails: the evolutionary novelty of venoms. Trends in ecology & evolution. 2013;28(4):219-29. Epub 2012/12/12. doi: 10.1016/j.tree.2012.10.020. PubMed PMID: 23219381.
» https://doi.org/10.1016/j.tree.2012.10.020 - 2. Ferraz CR, Arrahman A, Xie C, Casewell NR, Lewis RJ, Kool J, et al. Multifunctional toxins in snake venoms and therapeutic implications: From pain to hemorrhage and necrosis. Frontiers in ecology and evolution. 2019;7(218). doi: 10.3389/fevo.2019.00218.
» https://doi.org/10.3389/fevo.2019.00218 - 3. Das I. A field guide to the reptiles of South-East Asia: Bloomsbury Publishing; 2015.
- 4. Marlon R. 107+ ular Indonesia: panduan visual dan identifikasi lapangan. 2014.
- 5. Mohamed Abd El-Aziz T, Garcia Soares A, Stockand JD. Snake venoms in drug discovery: Valuable therapeutic tools for life saving. Toxins. 2019;11(10):564. doi: 10.3390/toxins11100564. PubMed PMID: 31557973.
» https://doi.org/10.3390/toxins11100564 - 6. Munawar A, Ali S, Akrem A, Betzel C. Snake venom peptides: Tools of biodiscovery. Toxins. 2018;10(11):474.
- 7. Siegel RL, Miller KD, Jemal A. Cancer Statistics, 2017. CA Cancer J Clin. 2017;67(1):7-30. doi: 10.3322/caac.21387. PubMed PMID: 28055103.
» https://doi.org/10.3322/caac.21387 - 8. Ferlay J, Soerjomataram I, Dikshit R, Eser S, Mathers C, Rebelo M, et al. Cancer incidence and mortality worldwide: sources, methods and major patterns in GLOBOCAN 2012. International journal of cancer. 2015;136(5):E359-86. Epub 2014/09/16. doi: 10.1002/ijc.29210. PubMed PMID: 25220842.
» https://doi.org/10.1002/ijc.29210 - 9. Li L, Huang J, Lin YH. Snake venoms in cancer therapy: Past, present and future. Toxins. 2018;10(9):346. doi: 10.3390/toxins10090346. PubMed PMID: 30158426.
» https://doi.org/10.3390/toxins10090346 - 10. Boohaker RJ, Lee MW, Vishnubhotla P, Perez JM, Khaled AR. The use of therapeutic peptides to target and to kill cancer cells. Current medicinal chemistry. 2012;19(22):3794-804. Epub 2012/06/26. doi: 10.2174/092986712801661004. PubMed PMID: 22725698; PubMed Central PMCID: PMCPmc4537071.
» https://doi.org/10.2174/092986712801661004 - 11. Trikha M, De Clerck YA, Markland FS. Contortrostatin, a snake venom disintegrin, inhibits beta 1 integrin-mediated human metastatic melanoma cell adhesion and blocks experimental metastasis. Cancer research. 1994;54(18):4993-8. Epub 1994/09/15. PubMed PMID: 7520832.
- 12. Zouari-Kessentini R, Srairi-Abid N, Bazaa A, El Ayeb M, Luis J, Marrakchi N. Antitumoral potential of Tunisian snake venoms secreted phospholipases A2. Biomed Res Int. 2013;2013:391389. Epub 2013/03/20. doi: 10.1155/2013/391389. PubMed PMID: 23509718; PubMed Central PMCID: PMCPMC3581298.
» https://doi.org/10.1155/2013/391389 - 13. Costa TR, Burin SM, Menaldo DL, de Castro FA, Sampaio SV. Snake venom L-amino acid oxidases: an overview on their antitumor effects. J Venom Anim Toxins Incl Trop Dis. 2014;20:23. Epub 2014/06/19. doi: 10.1186/1678-9199-20-23. PubMed PMID: 24940304; PubMed Central PMCID: PMCPMC4060840.
» https://doi.org/10.1186/1678-9199-20-23 - 14. Mong R, Tan HH. Snakebite by the shore pit viper (Trimeresurus purpureomaculatus) treated with polyvalent antivenom. Wilderness & environmental medicine. 2016;27(2):266-70. Epub 2016/04/12. doi: 10.1016/j.wem.2016.01.001. PubMed PMID: 27061038.
» https://doi.org/10.1016/j.wem.2016.01.001 - 15. Hutton RA, Looareesuwan S, Ho M, Silamut K, Chanthavanich P, Karbwang J, et al. Arboreal green pit vipers (genus Trimeresurus) of South-east Asia: bites by T. albolabris and T. macrops in Thailand and a review of the literature. Transactions of the Royal Society of Tropical Medicine and Hygiene. 1990;84(6):866-74. doi: https://doi.org/10.1016/0035-9203(90)90111-Q.
» https://doi.org/10.1016/0035-9203(90)90111-Q. - 16. Chan T, Hung LK. Digital gangrene following a green pit viper bite. The Southeast Asian journal of tropical medicine and public health. 2010;41(1):192-4. Epub 2010/06/29. PubMed PMID: 20578498.
- 17. Ozverel CS, Damm M, Hempel B-F, Göçmen B, Sroka R, Süssmuth RD, et al. Investigating the cytotoxic effects of the venom proteome of two species of the Viperidae family (Cerastes cerastes and Cryptelytrops purpureomaculatus) from various habitats. Comparative Biochemistry and Physiology part C: Toxicology & Pharmacology. 2019;220:20-30. doi: https://doi.org/10.1016/j.cbpc.2019.02.013
» https://doi.org/10.1016/j.cbpc.2019.02.013 - 18. Zainal Abidin SA, Rajadurai P, Hoque Chowdhury ME, Othman I, Naidu R. Cytotoxic, anti-proliferative and apoptosis activity of l-amino acid oxidase from Malaysian Cryptelytrops purpureomaculatus (CP-LAAO) venom on human colon cancer cells. Molecules. 2018;23(6).
- 19. Liew JL, Tan NH, Tan CH. Proteomics and preclinical antivenom neutralization of the mangrove pit viper (Trimeresurus purpureomaculatus, Malaysia) and white-lipped green pit viper (Trimeresurus albolabris, Thailand) venoms. Acta tropica. 2020;209:105528. doi: https://doi.org/10.1016/j.actatropica.2020.105528
» https://doi.org/10.1016/j.actatropica.2020.105528 - 20. Zainal Abidin SA, Rajadurai P, Chowdhury ME, Ahmad Rusmili MR, Othman I, Naidu R. Proteomic characterization and comparison of Malaysian Tropidolaemus wagleri and Cryptelytrops purpureomaculatus venom using shotgun-proteomics. Toxins 2016;8(10). Epub 2016/10/21. doi: 10.3390/toxins8100299. PubMed PMID: 27763534; PubMed Central PMCID: PMCPmc5086659.
» https://doi.org/10.3390/toxins8100299 - 21. Arruda Macêdo JK, Fox JW, de Souza Castro M. Disintegrins from snake venoms and their applications in cancer research and therapy. Current protein & peptide science. 2015;16(6):532-48. doi: 10.2174/1389203716666150515125002. PubMed PMID: 26031306.
» https://doi.org/10.2174/1389203716666150515125002 - 22. Rivas-Mercado EA, Garza-Ocañas L. Disintegrins obtained from snake venom and their pharmacological potential. University medicine magazine. 2017;19(74):32-7. doi: https://doi.org/10.1016/j.rmu.2017.02.004
» https://doi.org/10.1016/j.rmu.2017.02.004 - 23. Swenson S, Costa F, Ernst W, Fujii G, Markland FS. Contortrostatin, a snake venom disintegrin with anti-angiogenic and anti-tumor activity. Pathophysiology of haemostasis and thrombosis. 2005;34(4-5):169-76. Epub 2006/05/19. doi: 10.1159/000092418. PubMed PMID: 16707922.
» https://doi.org/10.1159/000092418 - 24. Hong SY, Koh YS, Chung KH, Kim DS. Snake venom disintegrin, saxatilin, inhibits platelet aggregation, human umbilical vein endothelial cell proliferation, and smooth muscle cell migration. Thrombosis research. 2002;105(1):79-86. Epub 2002/02/28. PubMed PMID: 11864711.
- 25. Chernyshenko V, Petruk N, Korolova D, Kasatkina L, Gornytska O, Platonova T, et al. Antiplatelet and antiproliferative action of disintegrin from Echis multisquamatis snake venom. Croatian medical journal. 2017;58(2):118-27.
- 26. de Oliveira IS, Manzini RV, Ferreira IG, Cardoso IA, Bordon KdCF, Machado ART, et al. Cell migration inhibition activity of a non-RGD disintegrin from Crotalus durissus collilineatus venom. Journal of venomous animals and toxins including tropical diseases. 2018;24(1):28.
- 27. Shahbazi B, Najafabadi ZS, Goudarzi H, Sajadi M, Tahoori F, Bagheri M. Cytotoxic effects of Pseudocerastes persicus venom and its HPLC fractions on lung cancer cells. Journal of venomous animals and toxins including tropical diseases. 2019;25.
- 28. Bradshaw MJ, Saviola AJ, Fesler E, Mackessy SP. Evaluation of cytotoxic activities of snake venoms toward breast (MCF-7) and skin cancer (A-375) cell lines. Cytotechnology. 2016;68(4):687-700. Epub 2014/11/20. doi: 10.1007/s10616-014-9820-2. PubMed PMID: 25407733; PubMed Central PMCID: PMCPmc4960119.
» https://doi.org/10.1007/s10616-014-9820-2 - 29. Laemmli UK. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature. 1970;227(5259):680.
- 30. Tan CH, Tan KY, Ng TS, Quah ESH, Ismail AK, Khomvilai S, et al. Venomics of Trimeresurus (Popeia) nebularis, the Cameron Highlands pit viper from Malaysia: Insights into venom proteome, toxicity and neutralization of antivenom. Toxins. 2019;11(2):95.
- 31. Consortium TU. UniProt: a worldwide hub of protein knowledge. Nucleic acids research. 2018;47(D1):D506-D15. doi: 10.1093/nar/gky1049.
» https://doi.org/10.1093/nar/gky1049 - 32. Roly ZY, Islam MM, Reza MA. A comparative in silico characterization of functional and physicochemical properties of 3FTx (three finger toxin) proteins from four venomous snakes. Bioinformation. 2014;10(5):281-7. doi: 10.6026/97320630010281. PubMed PMID: 24966535.
» https://doi.org/10.6026/97320630010281 - 33. Ma J, Chen T, Wu S, Yang C, Bai M, Shu K, et al. iProX: an integrated proteome resource. Nucleic acids research. 2019;47(D1):D1211-D7. doi: 10.1093/nar/gky869. PubMed PMID: 30252093.
» https://doi.org/10.1093/nar/gky869 - 34. Ozverel CS, Damm M, Hempel B-F, Göçmen B, Sroka R, Süssmuth RD, et al. Investigating the cytotoxic effects of the venom proteome of two species of the Viperidae family (Cerastes cerastes and Cryptelytrops purpureomaculatus) from various habitats. Comp Biochem Physiol C Toxicol Pharmacol. 2019;220:20-30. doi: https://doi.org/10.1016/j.cbpc.2019.02.013
» https://doi.org/10.1016/j.cbpc.2019.02.013 - 35. Damm M, Hempel B-F, Nalbantsoy A, Süssmuth RD. Comprehensive snake venomics of the Okinawa Habu pit viper, Protobothrops flavoviridis, by complementary mass spectrometry-guided approaches. Molecules. 2018;23(8):1893.
- 36. Kumkate S, Chanhome L, Thiangtrongjit T, Noiphrom J, Laoungboa P, Khow O, et al. Venomics and cellular toxicity of Thai pit vipers (Trimeresurus macrops and T. hageni). Toxins 2020;12(1). Epub 2020/01/23. doi: 10.3390/toxins12010054. PubMed PMID: 31963345; PubMed Central PMCID: PMCPMC7020458.
» https://doi.org/10.3390/toxins12010054 - 37. Hammouda MB, Montenegro MF, Sánchez-del-Campo L, Zakraoui O, Aloui Z, Riahi-Chebbi I, et al. Lebein, a snake venom disintegrin, induces apoptosis in human melanoma cells. Toxins. 2016;8(7):206.
- 38. Mills JB, Mant CT, Hodges RS. One-step purification of a recombinant protein from a whole cell extract by reversed-phase high-performance liquid chromatography. Journal of chromatography A. 2006;1133(1-2):248-53. Epub 09/01. doi: 10.1016/j.chroma.2006.08.042. PubMed PMID: 16945380.
» https://doi.org/10.1016/j.chroma.2006.08.042 - 39. Kang TS, Georgieva D, Genov N, Murakami MT, Sinha M, Kumar RP, et al. Enzymatic toxins from snake venom: structural characterization and mechanism of catalysis. The FEBS journal. 2011;278(23):4544-76. Epub 2011/04/08. doi: 10.1111/j.1742-4658.2011.08115.x. PubMed PMID: 21470368.
» https://doi.org/10.1111/j.1742-4658.2011.08115.x - 40. Zakraoui O, Marcinkiewicz C, Aloui Z, Othman H, Grepin R, Haoues M, et al. Lebein, a snake venom disintegrin, suppresses human colon cancer cells proliferation and tumor-induced angiogenesis through cell cycle arrest, apoptosis induction and inhibition of VEGF expression. Molecular carcinogenesis. 2017;56(1):18-35. Epub 2016/01/30. doi: 10.1002/mc.22470. PubMed PMID: 26824338.
» https://doi.org/10.1002/mc.22470 - 41. Saviola AJ, Modahl CM, Mackessy SP. Disintegrins of Crotalus simus tzabcan venom: Isolation, characterization and evaluation of the cytotoxic and anti-adhesion activities of tzabcanin, a new RGD disintegrin. Biochimie. 2015;116:92-102.
- 42. Calvete JJ, Schaefer W, Soszka T, Lu W, Cook JJ, Jameson BA, et al. Identification of the disulfide bond pattern in albolabrin, an RGD-containing peptide from the venom of Trimeresurus albolabris: Significance for the express of platelet aggregation inhibitory activity. Biochemistry. 1991;30(21):5225-9.
- 43. Knudsen KA, Tuszynski GP, Huang T-F, Niewiarowski S. Trigramin, an RGD-containing peptide from snake venom, inhibits cell-substratum adhesion of human melanoma cells. Experimental cell research. 1988;179(1):42-9. doi: https://doi.org/10.1016/0014-4827(88)90346-1
» https://doi.org/10.1016/0014-4827(88)90346-1 - 44. Soszka T, Knudsen KA, Beviglia L, Rossi C, Poggi A, Niewiarowski S. Inhibition of murine melanoma cell-matrix adhesion and experimental metastasis by albolabrin, an RGD-containing peptide isolated from the venom of Trimeresurus albolabris. Experimental cell research. 1991;196(1):6-12. Epub 1991/09/01. doi: 10.1016/0014-4827(91)90449-5. PubMed PMID: 1879472.
» https://doi.org/10.1016/0014-4827(91)90449-5 - 45. Eble JA, Niland S, Dennes A, Schmidt-Hederich A, Bruckner P, Brunner G. Rhodocetin antagonizes stromal tumor invasion in vitro and other alpha2beta1 integrin-mediated cell functions. Journal of the international society for matrix biology. 2002;21(7):547-58. Epub 2002/12/12. PubMed PMID: 12475639.
- 46. Danen EHJ, Marcinkiewicz C, Cornelissen IMHA, van Kraats AA, Pachter JA, Ruiter DJ, et al. The disintegrin eristostatin interferes with integrin α4β1 function and with experimental metastasis of human melanoma cells. Experimental cell research. 1998;238(1):188-96. doi: https://doi.org/10.1006/excr.1997.3821
» https://doi.org/10.1006/excr.1997.3821 - 47. Chung KH, Kim SH, Han Ky, Sohn YD, Chang SI, Baek KH, et al. Inhibitory effect of salmosin, a Korean snake venom derived disintegrin, on the integrin αv‐mediated proliferation of SK‐Mel‐2 human melanoma cells. Journal of pharmacy and pharmacology. 2003;55(11):1577-82.
- 48. Ramos OH, Kauskot A, Cominetti MR, Bechyne I, Salla Pontes CL, Chareyre F, et al. A novel alpha(v)beta (3)-blocking disintegrin containing the RGD motive, DisBa-01, inhibits bFGF-induced angiogenesis and melanoma metastasis. Clinical & experimental metastasis. 2008;25(1):53-64. Epub 2007/10/24. doi: 10.1007/s10585-007-9101-y. PubMed PMID: 17952617.
» https://doi.org/10.1007/s10585-007-9101-y - 49. Zhou Q, Nakada MT, Brooks PC, Swenson SD, Ritter MR, Argounova S, et al. Contortrostatin, a homodimeric disintegrin, binds to integrin alphavbeta5. Biochemical and biophysical research communications. 2000;267(1):350-5. Epub 2000/01/07. doi: 10.1006/bbrc.1999.1965. PubMed PMID: 10623623.
» https://doi.org/10.1006/bbrc.1999.1965 - 50. Bianconi D, Unseld M, Prager GW. Integrins in the spotlight of cancer. Int J Mol Sci. 2016;17(12):2037. doi: 10.3390/ijms17122037. PubMed PMID: 27929432.
» https://doi.org/10.3390/ijms17122037 - 51. Taherian A, Li X, Liu Y, Haas T. Differences in integrin expression and signaling within human breast cancer cells. BMC cancer. 2011;11:293. doi: 10.1186/1471-2407-11-293.
» https://doi.org/10.1186/1471-2407-11-293 - 52. Guruprasad K, Reddy BB, Pandit MW. Correlation between stability of a protein and its dipeptide composition: a novel approach for predicting in vivo stability of a protein from its primary sequence. Protein Eng. 1990;4(2):155-61.
- 53. Rogers S, Wells R, Rechsteiner M. Amino acid sequences common to rapidly degraded proteins: the PEST hypothesis. Science. 1986;234(4774):364-8. Epub 1986/10/17. doi: 10.1126/science.2876518. PubMed PMID: 2876518.
» https://doi.org/10.1126/science.2876518 - 54. Mishra J, Panda JJ. Short peptide-based smart targeted cancer nanotherapeutics: a glimmer of hope. Therapeutic delivery. 2019;10(3):135-8. Epub 2019/03/27. doi: 10.4155/tde-2019-0005. PubMed PMID: 30909857.
» https://doi.org/10.4155/tde-2019-0005 - 55. Bhawani SA, Husaini A, Ahmad FB, Asaruddin MR. Polymer based protein therapeutics. Current protein & peptide science. 2018;19(10):972-82. Epub 2017/08/23. doi: 10.2174/1389203718666170821162823. PubMed PMID: 28828988.
» https://doi.org/10.2174/1389203718666170821162823
-
Availability of data and material
Data generated and analyzed during this study were included in this published article. -
Funding
The present study was supported by research grants RF007C-2018 and PV058-2018 from the University of Malaya. -
Ethics approval
Not applicable. -
Consent for publication
Not applicable.
Supplementary material
Additional file 1. LC-MS/MS of purpureomaculin isolated from Malaysian Trimeresurus purpureomaculatus venom (Fraction 1)Publication Dates
-
Publication in this collection
17 July 2020 -
Date of issue
2020
History
-
Received
04 Feb 2020 -
Accepted
29 May 2020