Abstracts
Although dozens of fish ladders have been constructed at dams of Brazilian reservoirs, there are few studies evaluating their efficiency as a tool for the conservation of Neotropical ichthyofauna, especially for migratory species. Therefore, the present study evaluated the selectivity of the species that entered and ascended the fish ladder located next to Lajeado Dam (Luis Eduardo Magalhães Hydroelectric Power Plant) on the Tocantins River. Samples were taken monthly from November, 2002 through October, 2003, in the resting pools of the ladder, using cast nets, and in the downstream stretch, using gillnets. The selectivity of the ladder in attracting fish was evaluated by comparing the occurrence, relative abundance, dominance and the congruence of abundance ranks of migratory and non-migratory species in the ladder and in the stretch of river immediately downstream. Species richness and fish abundance in the resting pools were used to evaluate selectivity along the ladder. The effects on selectivity by temporal variations in water level downriver and maximum flow velocity in the fish ladder were also analyzed. Out of the 130 species recorded downriver, 62.3% were caught in the ladder, and migratory species were clearly favored. However, more than 2/3 of the catch belonged to only three species (Rhaphiodon vulpinus, Psectrogaster amazonica and Oxydoras niger). Although the majority of the species that entered the ladder were able to reach its top, there was a sharp reduction in abundance of individuals towards the top. Temporal variations in the water level below the dam influenced richness and abundance of fish concentrated downstream and in the ladder, with lower values during periods of low water. In the ladder, a maximum flow velocity of 2.3 m/s, although also selective, proved to be more appropriate for fish ascension than a velocity of 2.8 m/s. It was concluded that the entry and ascension of the fish in the ladder were not congruent with their proportions in the downriver stretch: fish samples in the ladder were clearly dominated by a few species, including some that do not need to be translocated. Thus, selectivity constitutes an important bottleneck to initiatives for translocating fish aimed at conserving their stocks or biodiversity. It is urgent to review the decision-making process for the construction of fish passages and to evaluate the functioning of those already operating.
Migratory fish; Fish conservation; Biodiversity; Reservoir; Lajeado Dam; Tocantins River
Dezenas de escadas de peixes foram construídas em barragens de reservatórios brasileiros, mas são raros os estudos acerca de suas eficiências como instrumentos de conservação da ictiofauna Neotropical, em especial de espécies migradoras. Neste contexto, o presente estudo teve como objetivo avaliar a seletividade específica no ingresso e ascensão de peixes na escada localizada junto à barragem de Lajeado (UHE Luis Eduardo Magalhães, rio Tocantins). Amostragens foram realizadas mensalmente de novembro de 2002 a outubro de 2003 nos tanques de descanso da escada, utilizando tarrafas, e no trecho a jusante, utilizando redes de espera. A avaliação da seletividade no ingresso da escada foi realizada através da comparação da ocorrência, abundância relativa, dominância e congruência dos ranks de abundância de espécies migradoras e não migradoras na escada e no trecho imediatamente a jusante. A riqueza e abundância específica nos diferentes tanques de descanso foram utilizadas para avaliar a seletividade ao longo da escada. Os efeitos das variações temporais do nível hidrométrico de jusante e da velocidade de fluxo na seletividade foram também analisados. Das 130 espécies registradas a jusante, 63,2% foram capturadas na escada, com claro favorecimento das espécies migradoras. Entretanto, mais de 2/3 das capturas pertenceram a apenas três espécies (Rhaphiodon vulpinus, Psectrogaster amazonica e Oxydoras niger). Embora a maioria das espécies que ingressa na escada possa alcançar seu topo, constatou-se uma redução pronunciada na abundância. Variações temporais no nível da água a jusante da barragem influenciaram a riqueza e a abundância de peixes que se concentram na entrada e dentro da escada, com redução nos valores durante períodos de níveis baixos. Já na escada, a velocidade de fluxo de 2,3 m.s-1, embora também seletiva, mostrou-se mais adequada à ascensão de peixes em relação a de 2,8 m.s-1. Conclui-se, portanto, que o ingresso e a ascensão de peixes na escada não são congruentes com a composição e relações de abundância das espécies no trecho a jusante. O conjunto de espécies que efetivamente ascende a escada é dominado por poucas espécies, podendo incluir entre as dominantes aquelas que não necessitam ser transpostas. Dessa maneira, a seletividade se constitui num importante gargalo nas iniciativas de transposição de peixes visando a conservação dos estoques ou da biodiversidade, sendo urgente a revisão do processo decisório sobre a construção destes dispositivos e a avaliação acerca do funcionamento daquelas já construídas.
Selectivity of fish ladders: a bottleneck in Neotropical fish movement
Carlos Sérgio AgostinhoI; Angelo Antônio AgostinhoII; Fernando PeliciceII; Deusimar Augusto de AlmeidaIII; Elineide Eugênio MarquesIII
INúcleo de Estudos Ambientais (Neamb), Universidade Federal do Tocantins, Campus de Porto Nacional, Rua 3, Quadra 17, Jardim dos Ipês, 77500000 Porto Nacional, TO, Brazil. agostinhocs@uft.edu.br
IINupelia, Universidade Estadual de Maringá, Av. Colombo, 5790, 87020900 Maringá, PR, Brazil. agostinhoaa@nupelia.uem.br
IIIPósGraduação Ciências do Ambiente, Universidade Federal do Tocantins. Av. NS 15, ALCNO 14, Bloco 3, Sala 15, 77010.970 Palmas, TO, Brazil
ABSTRACT
Although dozens of fish ladders have been constructed at dams of Brazilian reservoirs, there are few studies evaluating their efficiency as a tool for the conservation of Neotropical ichthyofauna, especially for migratory species. Therefore, the present study evaluated the selectivity of the species that entered and ascended the fish ladder located next to Lajeado Dam (Luis Eduardo Magalhães Hydroelectric Power Plant) on the Tocantins River. Samples were taken monthly from November, 2002 through October, 2003, in the resting pools of the ladder, using cast nets, and in the downstream stretch, using gillnets. The selectivity of the ladder in attracting fish was evaluated by comparing the occurrence, relative abundance, dominance and the congruence of abundance ranks of migratory and nonmigratory species in the ladder and in the stretch of river immediately downstream. Species richness and fish abundance in the resting pools were used to evaluate selectivity along the ladder. The effects on selectivity by temporal variations in water level downriver and maximum flow velocity in the fish ladder were also analyzed. Out of the 130 species recorded downriver, 62.3% were caught in the ladder, and migratory species were clearly favored. However, more than 2/3 of the catch belonged to only three species (Rhaphiodon vulpinus, Psectrogaster amazonica and Oxydoras niger). Although the majority of the species that entered the ladder were able to reach its top, there was a sharp reduction in abundance of individuals towards the top. Temporal variations in the water level below the dam influenced richness and abundance of fish concentrated downstream and in the ladder, with lower values during periods of low water. In the ladder, a maximum flow velocity of 2.3 m/s, although also selective, proved to be more appropriate for fish ascension than a velocity of 2.8 m/s. It was concluded that the entry and ascension of the fish in the ladder were not congruent with their proportions in the downriver stretch: fish samples in the ladder were clearly dominated by a few species, including some that do not need to be translocated. Thus, selectivity constitutes an important bottleneck to initiatives for translocating fish aimed at conserving their stocks or biodiversity. It is urgent to review the decisionmaking process for the construction of fish passages and to evaluate the functioning of those already operating.
Key words: Migratory fish, Fish conservation, Biodiversity, Reservoir, Lajeado Dam, Tocantins River.
RESUMO
Dezenas de escadas de peixes foram construídas em barragens de reservatórios brasileiros, mas são raros os estudos acerca de suas eficiências como instrumentos de conservação da ictiofauna Neotropical, em especial de espécies migradoras. Neste contexto, o presente estudo teve como objetivo avaliar a seletividade específica no ingresso e ascensão de peixes na escada localizada junto à barragem de Lajeado (UHE Luis Eduardo Magalhães, rio Tocantins). Amostragens foram realizadas mensalmente de novembro de 2002 a outubro de 2003 nos tanques de descanso da escada, utilizando tarrafas, e no trecho a jusante, utilizando redes de espera. A avaliação da seletividade no ingresso da escada foi realizada através da comparação da ocorrência, abundância relativa, dominância e congruência dos ranks de abundância de espécies migradoras e não migradoras na escada e no trecho imediatamente a jusante. A riqueza e abundância específica nos diferentes tanques de descanso foram utilizadas para avaliar a seletividade ao longo da escada. Os efeitos das variações temporais do nível hidrométrico de jusante e da velocidade de fluxo na seletividade foram também analisados. Das 130 espécies registradas a jusante, 63,2% foram capturadas na escada, com claro favorecimento das espécies migradoras. Entretanto, mais de 2/3 das capturas pertenceram a apenas três espécies (Rhaphiodon vulpinus, Psectrogaster amazonica e Oxydoras niger). Embora a maioria das espécies que ingressa na escada possa alcançar seu topo, constatouse uma redução pronunciada na abundância. Variações temporais no nível da água a jusante da barragem influenciaram a riqueza e a abundância de peixes que se concentram na entrada e dentro da escada, com redução nos valores durante períodos de níveis baixos. Já na escada, a velocidade de fluxo de 2,3 m.s1, embora também seletiva, mostrouse mais adequada à ascensão de peixes em relação a de 2,8 m.s1. Concluise, portanto, que o ingresso e a ascensão de peixes na escada não são congruentes com a composição e relações de abundância das espécies no trecho a jusante. O conjunto de espécies que efetivamente ascende a escada é dominado por poucas espécies, podendo incluir entre as dominantes aquelas que não necessitam ser transpostas. Dessa maneira, a seletividade se constitui num importante gargalo nas iniciativas de transposição de peixes visando a conservação dos estoques ou da biodiversidade, sendo urgente a revisão do processo decisório sobre a construção destes dispositivos e a avaliação acerca do funcionamento daquelas já construídas.
Introduction
Fish passes have been used as a strategy to attenuate the effects of the blockade that dams impose on the movements of fish in many river systems all over the world. Most of these mechanisms are based on ladders, i.e., structures that reduce water velocity and gradient so that fish can ascend and pass the dam (Agostinho et al., 2007a).
There are few studies on the efficiency of fish passages in South America; nevertheless, dozens of these mechanisms were constructed, in spite of their high cost and great effort. In most cases, fishways were constructed under legal constraints, and they were considered an end in themselves, not an instrument to guide management policies (Agostinho et al., 2002). Fish ladders recently constructed, either required by environmental control agencies or an initiative of the electrical power sector, are being monitored, and some observations have already been published (Godinho et al., 1991; Fernandez et al., 2004; Vono et al., 2004; Britto & Sirol, 2005; Capeleti & Petrere Jr., 2006). The first results have stimulated discussions on the subject and identified some important problems (Agostinho et al., 2003). One of the controversial aspects refers to the criteria used to assess their efficiency, which are generally based on the number of fish that ascend a ladder.
In an ideal situation, these mechanisms should have low species selectivity, allowing the maintenance of fish movements, leading to a population structure similar to the one observed prior to the river impoundment, or if no natural barrier was present before, with the same proportions of species as downstream. This would be especially desirable for large migratory fish, the group most affected by impoundments which may intercept their migration routes to spawning habitats. The divergence of these proportions may cause dramatic population imbalances for the fish assemblages upriver (Oldani & Baigún, 2002) and downriver from the dam (Agostinho et al., 2002).
Among the most critical aspects for the efficiency of fish passages is the mechanism that attracts the fish, which must allow shoals to recognize its entrance promptly (Clay, 1995). If fish do not recognize the entrance, they may remain in the vicinity for a prolonged period, delaying migration and jeopardizing spawning, or even resulting in no ascension of the ladder. The physical and chemical water conditions (e.g., temperature, water velocity and dissolved oxygen) below dams, where the shoals migrating upriver generally accumulate, are frequently stressful for fish and can lead to reabsorption of gonadal products by females (Agostinho et al., 1993).
The attraction mechanism of ladders is primarily hydraulic and must successfully compete with the attraction prompted by the tailrace or spillways in order to be effective. A fish ladder represents a loss of water for power generation, and its functioning may conflict with the hydroelectric function of a reservoir, especially in dischargelimited systems. However, because the more important upriver migrations occur in the rainy season, this conflict is lessened (C. S. Agostinho et al., 2007). Nevertheless, fishes show different preferences and abilities with respect to water flow, which may render a discharge adequate for a given group of fishes but restrictive to others (CastroSantos, 2004). Moreover, the dynamics of currents downriver from reservoirs, related to the distribution of fish shoals in this stretch of river, limits the alternatives for positioning the ladder entrance. The relationships of these dynamics with the operational procedures of a dam and the limitations imposed by the water intake on the positioning of the fish ladder exit into the reservoir are additional complicating factors.
In addition to attraction, the effectiveness of the fish passage also depends on the water velocity, turbulence, temperature, turbidity and large air bubbles along its length (Rodriguez et al., 2006). After the fish enter they must be able to overcome these obstacles and reach the reservoir, which depends on the different abilities of species to pass currents, and this frequently constitutes a source of selectivity in this stage of their journey.
The objective of this study was to evaluate the efficiency of the design and operation of a fish ladder located at Lajeado Dam on the Tocantins River. In particular, its efficiency was evaluated in regard to the attraction and ascension of the fish species based on their presence and abundance. This study sought to answer the following questions: (i) Is the ladder used equally by all the migratory species that arrive in the river stretch below the dam? (ii) Are the longdistance migratory species attracted more efficiently than the other species? (iii) Do the species that enter the ladder respond similarly to the distance to be traveled up to its top, and to the water velocity? (iv) Do changes in the water velocity in the ladder and in the water level downriver affect the attraction and the ascent of fish?
Material and Methods
Sampling localities
Sampling was carried out at the fish ladder constructed at Lajeado Dam (Hydroelectric Power Plant Luís Eduardo Magalhães) Fig.1, and in the river stretch immediately downstream from the dam. The ladder is a weir and orifice type, 874 m long and 5 m wide, with a 5% slope, for a total elevation gain of 36.8 m. It is located on the left bank, next to the spillway exit and the powerhouse. The ladder consists of an attraction channel (98 m to the first weir) and 92 weirs with submerged orifices (0.8 x 0.8 m), interspersed with two surface sills (0.5 x 1.0 m), in addition to five resting pools (still water rest). The first resting pool is excavated in rock (134 m from the first weir; 14.4 x 17.0 m), and the other pools are constructed of concrete (10 m x 10 m), located at distances of 278, 440, 595 and 725 m from the first weir. Because of the variations in water level (normal minimum = 211.5 m and normal maximum = 212.3 m), the ladder has four tilting gates to insure the maximum allowable elevation gain of 0.4 m per weir, in addition to four sluices and two main gates (elevation 210 m). During the sampling period the discharge was around 3.3 m3.s1, corresponding to a mean water velocity of 0.44 m.s1 in the pools and to a mean maximum flow velocity of 2.3 m.s1 in the bottom orifices and a mean depth of 1.5 m.
The stretch of river sampled downstream from Lajeado Dam was located about 1000 m below the ladder's attraction channel. In this stretch, the Tocantins River is about 400 m wide and has a mean depth of 20 m, with a rocky bed and sandy deposits in the backwaters.
Sampling and data analysis
Sampling was carried out from November, 2002 through October, 2003. In the ladder, sampling was conducted monthly in the five resting pools, using cast nets with a mesh size of 4.0 cm between alternate knots and a perimeter of 15 m, every 6 hours (12:00, 18:00, 24:00 and 06:00 h), with an effort of 10 casts each time and pool, beginning from the lowest pool. The fish caught were identified and measured, and those captured at 06:00 h had their gonadal development identified. Additionally, visual evaluations of fish movements were performed for fiveminute periods through the counting windows in the fifth pool, at 12:00, 18:00 and 06:00 h.
Collections downstream from the dam were made with gillnets with different mesh sizes (2.4 to 16 cm between alternate knots), set out for a 24hour cycle each month, and checked at 8:00, 16:00 and 22:00 h.
The species caught were classified according to their reproductive strategy, as (i) longdistance migrants or (ii) sedentary/shortdistance migrants (nonmigrants), based on the literature (Vazzoler & Menezes, 1992; Vazzoler, 1996; Carolsfeld et al., 2003).
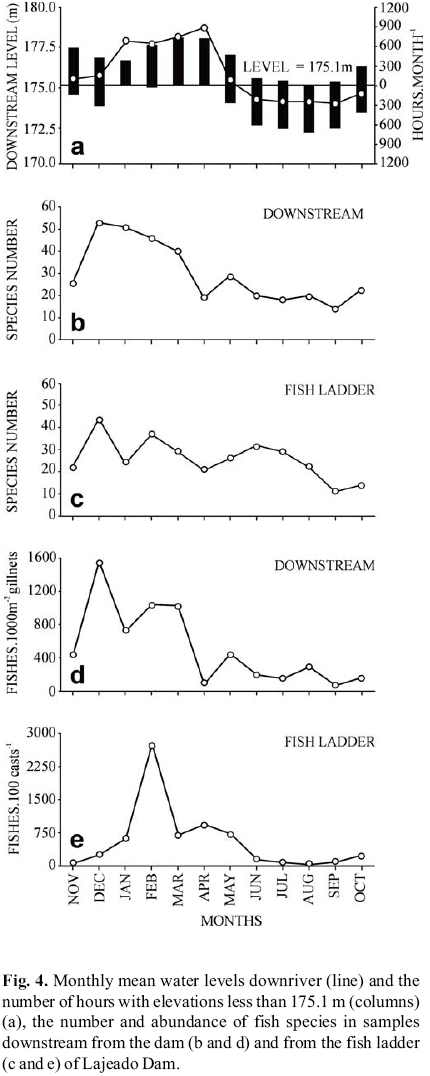
The abundance of fish was indexed by the catch per unit effort (CPUE), expressed as the number of individuals caught in 100 casts for the samples obtained in the ladder, and as the number of individuals per 1000 m2 gillnet.24 hours1 for the downstream collections. Occurrence, considered as the number of months in which each species was recorded during the study period, also includes the records of the visual evaluations in the fifth pool of the fish ladder.
To evaluate the selection of the species at the entrance of the ladder, the occurrence of each species and its relative abundance (% CPUE of the total catch in the period) in the ladder and downriver were compared. The dominance of the species in the assemblage, based on the equitability values derived from the Shannon index (Krebs, 1999), was also calculated for these two environments.
The hierarchy in the distribution of abundances was determined by the rank of abundances of the species downriver and in the ladder, for the migratory and nonmigratory species separately. In order to assess whether this hierarchy differed between the two sampling environments, the congruence (using Spearman correlation) of the downriverladder ranks was determined. On the occasion when a species was present downriver but absent from the ladder, we assigned to the species the last place in the ladder ranks. Similarly, when a species was absent downriver but present in the ladder, we assigned the last place in the downriver ranks. An analysis of variance (ANOVA) was applied to test if the mean congruence of the downriverladder ranks differed statistically between migratory and nonmigratory species, considering the months as replicates. Significant differences were taken as P < 0.05, using the program Statistica 5.0 (Statsoft, 2000).
To evaluate selection in the ascension of the ladder, the richness and mean abundance (CPUE) of the species were calculated for each resting pool. Additionally, to evaluate the performance of the most abundant species ascending the ladder, their occurrence and mean abundance were calculated separately for each of the five pools. A chisquare test was applied to the abundance data of these species in each pool, considering as expected values the total values of CPUE divided by the number of pools, assuming that in an ideal situation of free transit, the abundance of individuals of a given species would be similar over the length of the ladder. These results could be affected, to some extent, by the entrance of some fish from the reservoir, but according to Agostinho et al. (2007b) the downstream displacement represented only 0.39% of the number that ascended.
A possible influence of the variations in downriver water level on the selectivity of the ladder was explored graphically, interpreting the temporal variation in richness and abundance of the species over the months, in the ladder and downriver from the dam. The data for water level downstream from the dam were provided by INVESTCO S/A, the company that operates the L. E. Magalhães (Lajeado) Hydropower Plant.
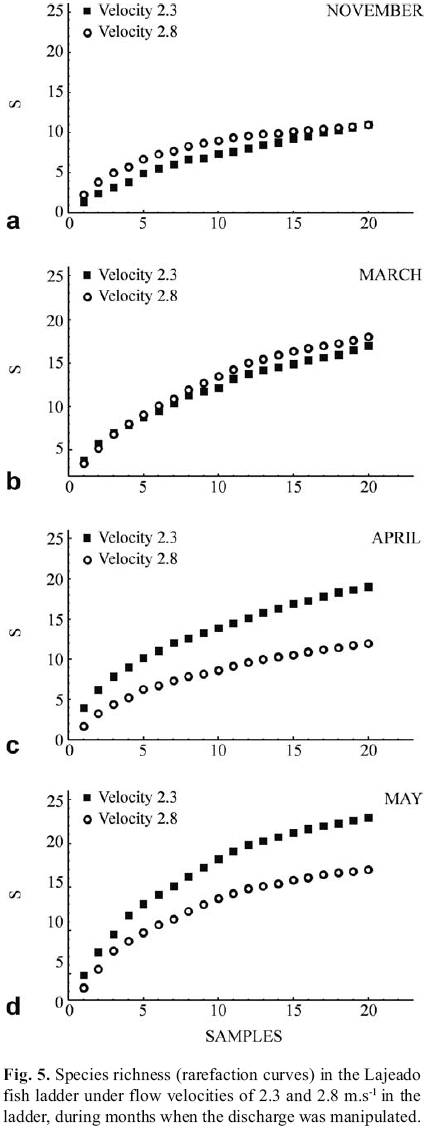
The effect of the mean flow velocity on the attraction of the ladder was determined through tests carried out with discharges of 3.3 and 4.6 m3.s1, which result in mean velocities of 2.3 and 2.8 m.s1, respectively, in the bottom openings (maximum flow velocity). These experiments were conducted in November, 2002 and in March, April and May of 2003. The selection of discharge levels into the ladder was limited by information regarding the operational maximum, considering the recommended discharge for the power plant. The effect of the two flow velocities on the number of fish species ascending the ladder was evaluated by rarefaction curves (species x samples), calculated after 50 randomizations of the original data matrix. The program EstimateS 5.0 (Colwell, 1997) was used for this analysis. In addition, the total species richness for each month and each value of the flow velocity in the ladder was estimated using the MichaelisMenten hyperbolic equation. In this case, the asymptote of the adjusted curve represents the total expected species richness based on the data set analyzed.
Results
Selection in the attraction
The fish fauna in the stretch downriver from Lajeado Dam was composed of 130 species, of which 81 (62.3%) were recorded in the ladder. The proportion of longdistance migratory species in the ladder was higher than nonmigratory; they comprised 75.0% of the 32 species present downriver. The abundance of migratory species surpassed that of nonmigratory species in the ladder by a 3:1 ratio, an opposite trend compared to that recorded downriver (Table 1).
With respect to abundance, there was markedly high dominance in the catches in the ladder: only three species comprised 2/3 of the total catch. The estimated equitability values, that indicate the proportion in abundance among species, were 0.73 downstream and 0.53 in the ladder, indicating a considerably greater dominance in the latter.
The occurrence of migratory species in the 12 months of sampling shows that the most abundant species were also the most frequent in the monthly samples conducted in the ladder, with the exception of Psectrogaster amazonica (Table 1). For some species, occurrence was higher in the ladder than in the downriver stretch.
In regard to the hierarchy of abundances, the congruence of the downriverladder ranks (Spearman correlation) varied from 0.47 to 0.56 for the migratory species, with a mean of 0.10 ± 0.30 SD. For the nonmigratory species, all the relationships between the downstreamladder ranks were negative, with congruence rankings varying from 0.71 to 0.05, with a mean of 0.38 ± 0.24 SD. Mean congruences were significantly higher for migratory than for nonmigratory species (ANOVA; F1; 22 = 18.26; p = 0.0031).
Selection along the ladder
The number of species that after entering succeeded in ascending to the different points of the ladder, did not differ markedly either for migratory species or for the sedentary/shortdistance migratory ones (Fig. 2). Thus, the majority of the species recorded in the lower stretches also occurred at the uppermost resting pools. Nevertheless, important variations were observed in abundance values, with a general tendency to decrease towards the top of the ladder, except for the migratory Rhaphiodon vulpinus (Fig. 2).
The ability to overcome the obstacles posed by the water dynamics (velocity and turbulence) was analyzed considering that in an ideal situation of free transit, the abundance of the individuals of a given species would be similar along the ladder. Four species, which together comprised 78% of the catches, were selected for this evaluation (Fig. 3). Three of them (Auchenipterus nuchalis, Oxydoras niger and Psectrogaster amazonica) tended to decrease in frequency in the monthly samples and in abundance toward the top of the ladder. Rhaphiodon vulpinus showed an opposite tendency; it accumulated in the last resting pool, where it reached abundance values at least three times higher than that of the other three species. The differences between the actual CPUE values and the expected values (similar values along the ladder), calculated for each species, were all significant (chisquare test; c2 = 87.9; df = 4; p < 0.05).
Influence of the downstream water level on selectivity
The number of species and their abundances, both in the ladder and in the river channel downstream, were related to the temporal variations in water level downriver, affected by the water discharged during operation of the dam and by the seasonality of the hydrological cycle (Fig. 4). Thus, when the mean monthly elevation of the downstream stretch was less than 175.1 m (threshold of the upper sill of the first weir) or remained below this level for many hours per month, species richness was lower downriver and the number of species in the ladder decreased. The lowest abundance values were also observed on these occasions. On the other hand, the variations in number of species in the ladder were less pronounced, with relatively high numbers from June through August of 2003. This finding is probably related to an increased number of species residing in the ladder, or even to individuals that entered the ladder from the reservoir. The tendencies toward a small number of species in April, 2003 in the downriver samples are results of the reduction in sampling effort.
Influence of water flow velocity on selectivity
Manipulation of the two flow velocities within the predicted range for operation of the ladder, carried out over four months, revealed that species richness, estimated from the rarefaction curve, tended to be higher at the velocity of 2.3 m.s1 (discharge 3.3 m.s1) than at 2.8 m.s1 (discharge 4.6 m3.s1) (Fig. 5). An exception was March, 2003, when torrential rains fell and water transparency was reduced, and slightly opposite tendencies were observed. The values of total richness estimated from the MichaelisMenten equation for the velocities of 2.3 and 2.8 m.s1 were, respectively, 18.8 and 13.8 species in November, 2003, 25.3 and 17.3 in April, and 30.5 and 22.4 in May. In March, these values were 22.4 and 25.9 species, respectively. The mean abundance of fish was also higher during periods of lower discharge (CPUEvelocity:2,3=744 ind./100 casts, Svelocity:2,3=558; CPUEvelocity:2,8=505, Svelocity:2,8=483).
Discussion
Fish passages are designed to reestablish connectivity between populations, or their habitats, which were fragmented by dams. Although the constructions of these devices have vague objectives in regard to the species targeted, they can be considered efficient when shoals of fish migrating upriver can reach the stretches that they originally occupied, and migrants are translocated in similar proportions to those existing downriver from a dam. However, the selectivity of these structures has been shamefully neglected in evaluations of these facilities for the conservation of Neotropical fish fauna (Agostinho et al., 2002).
Efficiency and effectiveness of fish passages are better evaluated by mark and recapture experiments, especially using radiotelemetry, because the major interest is the passage, not the presence. However, the large number of species and sizes, dimension of the river and the huge effort and cost involved, make it difficult or even impossible to use of this technique. However, fish presence and abundance below and in the different positions of the ladder gave satisfactory indication about which species and how many individuals are attracted and can reach this fish passage and thus on its selectivity. Netting fish at a particular site can, therefore, be used to quantify migration or movements in a particular locality (Quiros & Vidal, 2000)
Comparisons of the observed percentages of species present downriver that entered the ladder, in the present study and those published for two other basins in the Neotropics, indicate only moderate selectivity (62% of total downstream species). Meanwhile, evaluations of the experimental ladder at Itaipu Dam on the Paraná River (Fernandez et al., 2004) revealed a percentage entry of only 42% of the downstream species. Higher values were recorded at Igarapava Dam on the Grande River (Vono et al., 2004) and in the Tijuco River, a small tributary of the Grande (Salto Moraes Dam; Godinho et al., 1991), with percentages of 71% and 82%, respectively. The higher proportion of migratory species present downstream that entered the ladder, and their consequently higher proportion in the ladder when compared to downriver, is expected due to the more pronounced rheophilic behavior of species with this strategy. On the other hand, comparison of abundance ranks showed that the structure of the downstream community is strongly altered in the ladder.
The selectivity of this mechanism appears to be very important, even for migratory species: the low congruence values indicated that there was little similarity in the distribution of abundances between these two compartments (ladder downriver). Even considering the strong swimming ability, characteristic of migratory species, only a part of them succeed in recognizing the entrance of the ladder. The analysis of the nonmigratory species, which are less prone to ascend the ladder, emphasized the strong selectivity of the mechanism. The negative congruence of ranks reflected the tendency of the more abundant species downriver to be relatively rare on the ladder, and viceversa. Thus, because of this drastic alteration in the abundance hierarchy, the ladder at Lajeado Dam was highly selective in translocating the downstream species, especially the nonmigratory ones. The high dominance in the ladder is demonstrated by the fact that nearly 70% of the individuals recorded in it belonged to only three species. Future population imbalances in upstream stretches may occur through the continual alteration of this hierarchy, and consequently through modification of species interactions downriver and upriver from the ladder.
The lack of variation in the number of species along the ladder, was also observed for the experimental ladder at Itaipu (Fernandez et al., 2004), along with the decreasing gradient in abundance. These findings suggest that, although many species are able to reach the top of the ladder, the turbulence and velocity of the water offer some resistance to ascending movement for a considerable part of the populations, leading to a concentration of individuals in the first sections. This was particularly evident for three of the four most abundant species, especially for Psectrogaster amazonica, which comprised almost 40% of the captures in pool 2 but was rare in pool 5. The exception was Rhaphiodon vulpinus, which showed an inverse density gradient, attributed to the intense predation pressure existing at the entrance of the reservoir (personal observation). Godinho et al. (1991), on the other hand, reported that at Salto Moraes Dam, there was a notable reduction in the number of species and individuals towards the top of the ladder, and only 17.6% of the species and 2% of the individuals that entered it reached the top. Contrary to the observations at Lajeado, the ladder at Salto Moraes (which is about 10 times shorter: 78.3 m) is less selective to the entry of fish, but more selective to their ascent. This identifies the need for studies of selectivity in ladders to quantify not only the attraction, but the ability of individual fish in reaching the upper end of these passages.
The operational procedures of hydroelectric dams, along with the rainfall seasonality, can cause profound and recurring changes in water level downstream, affecting the local abundance of fish, altering currents and modifying the efficiency in attracting fish to the ladder. In the present study, both species richness and abundance of individuals were related to the river level downstream of the Lajeado Dam. The abundance of fish in the ladder was severely reduced when water level remained low, although the decline in number of species was significant only at the end of the period. In the case of the Lajeado ladder, an important factor is that water levels below 175.1 m, through a fault of the project and planning, leave the sill of the first pool out of water and impede the entrance of fish in the ladder. Thus, the observation that a certain number of species were present in the ladder and this number did not change markedly during the first three months when the water level remained below this threshold, may be explained by the fact that many species were living in the ladder (Agostinho et al., 2007a), or were entering from the reservoir. The latter possibility is, however, less probable, given that the species which enter the ladder from the reservoir are few and present only in low numbers (Agostinho et al., 2007b). The importance of increasing the discharge downriver in attracting fish to enter and ascend a ladder was described by Pompeu & Martinez (2006) for the Santa Clara power plant, in the Mucuri River in the Atlantic basin.
The flow velocity in the ladder, which can represent an obstacle to fish ascension, also plays an important role in attracting fish (Clay, 1995; Larinier & Murmulla, 2003). For the Lajeado fish ladder, the species richness as well as abundances increased at moderate flow velocities (2.3 m.s1). This tendency, which was predicted in the original ladder project, may be slightly modified depending on environmental conditions. Thus, in the only month of the experimental period (March) in which intense rains and low water transparency occurred, there was a slight inversion in these tendencies, with increased attraction in conditions of greater water velocity. Albeit the hydrological level was higher and more constant in March and April than the other months studied, species richness and abundances were substantially higher in March. Thus, when shoals concentrated below the dam, the differences in flow velocity are not important, probably because of the motivation to migrate.
Although some refinement is needed, the studies conducted at the Lajeado Dam fish ladder indicate that several species that live below the entrance are attracted and able to ascend it. As expected, the ladder is more effective in attracting migratory fishes, as demonstrated by their higher relative abundance in the ladder. Nevertheless, the majority of the species of this group are only sporadically present in the ladder, and many were recorded only downriver. This selectivity was also seen along the ladder, as revealed by the strong gradient in abundance inside the ladder (it decreased from the entrance to the top), even among the most abundant species. In addition, the water level below the dam was associated with the species richness and the abundance of fish downriver, affecting the entry of fish into the ladder. Manipulation of the discharge in the ladder played a similar role, where more fish tended to enter under conditions of moderate flow velocities, especially when fish density below the dam was lower.
Finally, it is appropriate to emphasize that evaluation of the efficiency of ladders and other mechanisms for fish transposition in Brazil is an inglorious task, given that these facilities are, in general, constructed with vague or imprecise objectives (passage of fish), under strong legal constraints. Given that any mechanism for moving fish will be highly speciesselective, it is necessary to define the target species to be moved in order to guide the design. Because these structures are only management tools, this must be decided in the context of a general plan that envisions the resolution of known problems, under the penalty of allowing priority access to species that do not need to ascend the upriver stretches, spreading demographic imbalances to both sides of the dam. It is not uncommon that fishmoving mechanisms facilitate the passage of undesirable species, or even remove desirable species from stretches where they might have sustainable stocks, and trapping them in stretches where they cannot succeed in completing their life cycles (Agostinho et al., 2007a; Britto & Sirol, 2005).
Therefore, selectivity is an important bottleneck to initiatives to translocate fish with the aim of conserving their stocks or biodiversity. It is not, however, the only one; the intensification of predation, the delay in moving toward the breeding sites, the return of migrants and the recruitment of new individuals to the stocks downriver from the dam are other impediments that could render the operation of a fish passage inefficient or even environmentally undesirable. It is, therefore, urgent to review the decisionmaking process for construction of these facilities and to evaluate the functioning of those already constructed.
Acknowledgments
We thank our colleagues of the Núcleo de Estudos Ambientais (Neamb), Universidade Federal do Tocantins (UFT), for their assistance in the field and laboratory work, Sidney Lazaro Martins for hydraulic data, and Jaime Luiz Lopes Pereira for drawing the figures. We also thank Investco S.A. for financial support.
Literature Cited
Received January 2007
Accepted May 2007
- Agostinho, A. A., L. C. Gomes, D. R. Fernandez & H. I. Suzuki. 2002. Efficiency of fish ladders for neotropical ichthyofauna. River Research and Applications, 18(3): 299306.
- Agostinho, A. A., L. C. Gomes & F. M. Pelicice. 2007a. Ecologia e manejo de recursos pesqueiros em reservatórios do Brasil. EDUEM, Maringá. 502p.
- Agostinho, A. A., L. C. Gomes, H. I. Suzuki & H. F. Júlio Jr. 2003. Migratory fish from the upper Paraná River basin, Brazil. Pp. 1999. In: Carolsfeld, J., B. Harvey, C. Ross & A. Baer (Eds.). Migratory fishes of South America: biology, social importance and conservation status. World Fisheries. World Fisheries Trust, British Columbia, Canada. 380p.
- Agostinho, A. A., E. E. Marques, C. S. Agostinho, D. A. de Almeida, R. J. de Oliveira & & J. R. B. de Melo. 2007b. Fish ladder of Lajeado Dam: migrations on oneway routes? Neotropical Ichthyology, 5(2): 121130.
- Agostinho, A. A., V. P. Mendes, H. I. Suzuki & C. Canzi. 1993. Avaliação da atividade reprodutiva da comunidade de peixes dos primeiros quilômetros a jusante do reservatório de Itaipu. Revista UNIMAR, Maringá, 15(Supplement): 175189.
- Agostinho, C. S., C. R. Pereira, R. J. Oliveira, I. S. Freitas & E. E. Marques. 2007. Movements through a fish ladder: temporal patterns and motivations to move upstream. Neotropical Ichthyology, 5(2): 161167.
- Britto, S. G. C. & R. N. Sirol. 2005. Transposição de peixes como forma de manejo: as escadas do complexo Canoas, médio Paranapanema, bacia do Alto Paraná. Pp. 285304. In: M. G. Nogueira, R. Henry & A. Jorcin. Ecologia de reservatórios: impactos potenciais, ações de manejo e sistemas em cascata. Rima, São Carlos. 472p.
- Capeleti, A. R. & M. Petrere Jr. 2006. Migration of the curimbatá Prochilodus lineatus (Valenciennes, 1836) (Pisces, Prochilodontidae) at the waterfall "Cachoeira de Emas" of the MogiGuaçu River São Paulo, Brazil. Brazilian Journal of Biology, 66(2B): 651659.
- Carolsfeld, B. Harvey, C. Ross & A. Baer (Eds.). 2003. Migratory fishes of South America: biology, fisheries and conservation status. World Fisheries Trust, British Columbia, Canada. 380p.
- CastroSantos, T. 2004. Quantifying the combined effects of attempt rate and swimming capacity on passage through velocity barriers. Canadian Journal of Fisheries and Aquatic Sciences, 61(9):16021615.
- Clay, C. H. 1995. Design of fishways and other fish facilities. 2nd ed. Boca Raton: Lewis Publishers, 248p.
- Colwell, R. K. 1997. EstimateS 5: statistical estimation of species richness and shared species from samples. Version 5.0.1.
- Fernandez, D. R., A. A. Agostinho & L. M. Bini. 2004. Seletividade em uma escada de peixes experimental na barragem do reservatório de Itaipu. Brazilian Archives of Biology and Technology, 47(4): 579586.
- Godinho, H. P., A. L. Godinho, P. S. Formagio & V. C. Torquato. 1991. Fish ladder efficiency in a southeastern Brazilian river. Ciência e Cultura, 43(1): 6367.
- Krebs, C. J. 1999. Ecological methodology. New York: Harper & Row, 620p.
- Larinier, M. & Murmulla, G. 2003. Fish passes: types, principles and geographical distribution : an overview. Second International Symposium on the Management of Large Rivers for Fisheries Sustaining Livelihoods and Biodiversity in the New Millennium. Phnom Penh, Cambodia. Pp. 183205
- Oldani, N. O., C. R. M. Baigún. 2002. Performance of a fishway system in a major South American dam on the Paraná River (ArgentinaParaguay). River Research and Applications, 18(2): 171183.
- Pompeu, P. dos S. & C. B. Martinez. 2006. Temporal patterns of fish passage in Santa Clara Power Plant's fish lift, Murici River, east Brazil. Revista Brasileira de Zoologia, 23(2): 340349.
- Quiros, R & J. C. Vidal. 2000. Cyclic behaviour of potamodromous fish in large river. Pp. 7186. In: I. G. Cowx (Ed.), Management and ecology of river fisheries. Fishing News Book, Blackwell Science Ltd, Oxford. 446p.
- Rodríguez, T. T., J. P. Agudo, L. P. Mosquera & E. P. González. 2006. Evaluating verticalslot fishway designs in terms of fish swimming capabilities. Ecological Engineering, 27:3748.
- Statsoft, Inc. 2000. Statistica for Windows (Computer manual program). Tulsa, OK.
- Vazzoler, A. E. A. de M. 1996. Biologia da reprodução de peixes teleósteos: teoria e prática. EDUEM, Maringá, Paraná. 169 p.
- Vazzoler, A. E. A. de M. & N. A. Menezes. 1992. Síntese de conhecimentos sobre o comportamento reprodutivo dos Characiformes da América do Sul (Teleostei, Ostariophysi). Revista Brasileira de Biologia, 52(4):627640.
- Vono, V., P. M. Bizzotto, H. P. Godinho, A. L. Godinho & B. Kynard. 2004. Fish passage at the Igarapava fish ladder, Rio Grande, Brazil. Pp. 15. In: Annals of the International Congress on the Biology of Fish, Tropical Hotel Resort, Manaus. American Fisheries Society.
Publication Dates
-
Publication in this collection
24 July 2007 -
Date of issue
2007
History
-
Accepted
May 2007 -
Received
Jan 2007