Abstracts
Ituglanis mambai, new species, is described from a cave in the Mambaí karst area, State of Goiás, Central Brazil. The new species distinguishes from epigean and cave congeners by the combination of the following characteristics: posterior supraoccipital fontanel absent; pectoral-fin rays usually i,7; six pleural ribs; total vertebrae 37-38 behind Weberian apparatus; shorter predorsal length (65.1-70.8% SL); shorter caudal peduncle length (8.4-11.9% SL); shorter dorsal-fin base length (7.7-11.3% SL); wider interobital width (29.2-36.5% HL); larger mouth width (43.4-64.0% HL); intermediate between epigean and other cave Ituglanis species as regards to both eyes (diameter varying from 0.5 to1.0 mm in adults, 7.8-10.1 % HL) and pigmentation, composed by irregular light brown spots along the body. The latter indicate the troglobitic status for I. mambai. In addition, this species has the maxillae with a discrete medial-posterior projection; fronto-lacrimal one half-length of the maxillae and pointed backwards; posterior process of palatine half its length, with a tenuous medial concavity; 14 dorsal and 12 ventral procurrent rays. In the natural habitat, I. mambai displayed cryptobiotic habits, trying to hide in the graveled bottom or under boulders when disturbed, apparently showing a negative response to light. It was observed a preference to slow-moving waters. Recent flood marks were observed in the stream conduit in March/April 2007 (end of the rainy season) when less individuals were observed on 300 m of the subterranean stream compared to September 2004 (end of the dry season).
New cave catfish; Rio Tocantins basin; Taxonomy; Brazil
Ituglanis mambai, nova espécie, é descrita de uma caverna localizada na área cárstica de Mambaí, estado de Goiás, Brasil central. A nova espécie difere dos congêneres epígeos e cavernícolas pela combinação das seguintes características: fontanela posterior do supraoccipital ausente; raios da nadadeira peitoral usualmente i,7; seis costelas; número total de vértebras 37-38; comprimento pré-dorsal menor, 65.1-70.8% do comprimento-padrão; comprimento do pedúnculo caudal menor, 8.4-11.9% do comprimento-padrão; base da nadadeira dorsal menor, 7.7-11.3% do comprimento-padrão; altura maior da cabeça, 46.7-71.6% do comprimento da cabeça; largura da cabeça menor, 89.2-115.7% do comprimento da cabeça; largura interobital maior (29.2-36.5% do comprimento da cabeça); largura da boca maior (43.4-64.0% do comprimento da cabeça); olhos e pigmentação intermediários entre as espécies epígeas e cavernícolas de Ituglanis - diâmetro dos olhos variando entre 0.5 e 1.0 mm nos adultos, 7.8-10.1 % do comprimento da cabeça e pigmentação composta por manchas irregulares marrom-claras ao longo do corpo. Este último caráter indica o status de troglóbio de I. mambai. Adicionalmente, esta espécie apresenta maxila com uma discreta projeção medial-posterior; fronto-lacrimal 1,5 vezes o tamanho da maxilla e projetado posteriormente; comprimento do processo posterior do palatino metade do seu comprimento total, com uma tênue concavidade medial; 14 raios procurrentes dorsais e 12 ventrais. No ambiente natural, I. mambai mostrou hábitos criptobióticos. Quando perturbados, tentavam entocar-se sob os seixos e matacões. Aparentemente mostraram reação negativa à luz. Foi observada uma preferência por remansos. Marcas de enchentes recentes foram observadas no conduto do riacho subterrâneo em março/abril 2007 (fim da estação chuvosa) quando um número menor de indivíduos foi observado, comparando-se a setembro de 2004 (final da estação chuvosa).
Ituglanis mambai, a new subterranean catfish from a karst area of Central Brazil, rio Tocantins basin (Siluriformes: Trichomycteridae)
Maria Elina BichuetteI; Eleonora TrajanoII
IDepartamento de Ecologia e Biologia Evolutiva, Universidade Federal de São Carlos, Via Washington Luis, km 235, C.P. 676, 13565-905 São Carlos, SP, Brazil. bichuette@uol.com.br
IIDepartamento de Zoologia, Instituto de Biociências, Universidade de São Paulo. C.P. 11461, 05422-970 São Paulo, SP, Brazil. etrajano@usp.br
ABSTRACT
Ituglanis mambai, new species, is described from a cave in the Mambaí karst area, State of Goiás, Central Brazil. The new species distinguishes from epigean and cave congeners by the combination of the following characteristics: posterior supraoccipital fontanel absent; pectoral-fin rays usually i,7; six pleural ribs; total vertebrae 37-38 behind Weberian apparatus; shorter predorsal length (65.1-70.8% SL); shorter caudal peduncle length (8.4-11.9% SL); shorter dorsal-fin base length (7.7-11.3% SL); wider interobital width (29.2-36.5% HL); larger mouth width (43.4-64.0% HL); intermediate between epigean and other cave Ituglanis species as regards to both eyes (diameter varying from 0.5 to1.0 mm in adults, 7.8-10.1 % HL) and pigmentation, composed by irregular light brown spots along the body. The latter indicate the troglobitic status for I. mambai. In addition, this species has the maxillae with a discrete medial-posterior projection; fronto-lacrimal one half-length of the maxillae and pointed backwards; posterior process of palatine half its length, with a tenuous medial concavity; 14 dorsal and 12 ventral procurrent rays. In the natural habitat, I. mambai displayed cryptobiotic habits, trying to hide in the graveled bottom or under boulders when disturbed, apparently showing a negative response to light. It was observed a preference to slow-moving waters. Recent flood marks were observed in the stream conduit in March/April 2007 (end of the rainy season) when less individuals were observed on 300 m of the subterranean stream compared to September 2004 (end of the dry season).
Key Words: New cave catfish, Rio Tocantins basin, Taxonomy, Brazil.
RESUMO
Ituglanis mambai, nova espécie, é descrita de uma caverna localizada na área cárstica de Mambaí, estado de Goiás, Brasil central. A nova espécie difere dos congêneres epígeos e cavernícolas pela combinação das seguintes características: fontanela posterior do supraoccipital ausente; raios da nadadeira peitoral usualmente i,7; seis costelas; número total de vértebras 37-38; comprimento pré-dorsal menor, 65.1-70.8% do comprimento-padrão; comprimento do pedúnculo caudal menor, 8.4-11.9% do comprimento-padrão; base da nadadeira dorsal menor, 7.7-11.3% do comprimento-padrão; altura maior da cabeça, 46.7-71.6% do comprimento da cabeça; largura da cabeça menor, 89.2-115.7% do comprimento da cabeça; largura interobital maior (29.2-36.5% do comprimento da cabeça); largura da boca maior (43.4-64.0% do comprimento da cabeça); olhos e pigmentação intermediários entre as espécies epígeas e cavernícolas de Ituglanis - diâmetro dos olhos variando entre 0.5 e 1.0 mm nos adultos, 7.8-10.1 % do comprimento da cabeça e pigmentação composta por manchas irregulares marrom-claras ao longo do corpo. Este último caráter indica o status de troglóbio de I. mambai. Adicionalmente, esta espécie apresenta maxila com uma discreta projeção medial-posterior; fronto-lacrimal 1,5 vezes o tamanho da maxilla e projetado posteriormente; comprimento do processo posterior do palatino metade do seu comprimento total, com uma tênue concavidade medial; 14 raios procurrentes dorsais e 12 ventrais. No ambiente natural, I. mambai mostrou hábitos criptobióticos. Quando perturbados, tentavam entocar-se sob os seixos e matacões. Aparentemente mostraram reação negativa à luz. Foi observada uma preferência por remansos. Marcas de enchentes recentes foram observadas no conduto do riacho subterrâneo em março/abril 2007 (fim da estação chuvosa) quando um número menor de indivíduos foi observado, comparando-se a setembro de 2004 (final da estação chuvosa).
Introduction
Brazil has a remarkable subterranean ichthyofauna, similar to few other countries or geographically comparable karst areas, such as Mexico, China and the southeastern Asia. To date, nearly 21 species (several awaiting formal description) of Brazilian subterranean fishes are known to present reduction of eyes and pigmentation at least at some degree beyond that observed in their epigean congeners, suggesting a troglobitic (exclusively subterranean) status for these fishes, and this number is increasing every year (Trajano, 1997; Bichuette & Trajano, 2003; Trajano & Bichuette, 2004). The majority (all but two) are siluriformes belonging to three out of 12 catfish families found in Brazil. Catfish species are also predominant in groundwaters of other Neotropical countries, whereas troglobitic cypriniforms largely outnumber catfishes in subterranean habitats in China, Thailand and other Asian countries.
The Neotropical Trichomycteridae forms a monophyletic group of small-sized catfishes, with Cis- and Transandine species, representing one of the richest group of siluriforms with at least 200 nominal species in 35 genera (de Pinna, 1998; Wosiacki, 2002; de Pinna & Wosiacki, 2003). The genus Trichomycterus Valenciennes, 1833 is the largest of the family (over 100 species) and one of the catfish genera best represented in the cave environment, with several troglomorphic species. Up to now, at least eight species were reported: T. chaberti Durand, 1968 from Bolivia; T. conradi (Eigenmann, 1912) and T. spelaeus DoNascimento, Villarreal & Provenzano, 2001, both from Venezuela; T. itacarambiensis Trajano & de Pinna, 1996 from eastern Brazil; T. santanderensis Castellanos-Morales, 2007, and three undescribed species, one from Colombian Andes (Sket, 1988) and two respectively from northeastern and southwestern Brazil (unpubl. data).
Costa & Bockmann (1993) erected the genus Ituglanis to include nine species previously placed in Trichomycterus, based on three autapomorphies (supraoccipital fontanel reduced to a small round orifice, a palatine with a deep medial concavity, and the anterior extremity of the sphenotic directed anteriorly). The genus Ituglanis is distributed on the Cisandine South America, encompassing 18 described species (Costa & Bockmann, 1993; Fernández & Bichuette, 2002; de Pinna & Keith, 2003; Bichuette & Trajano, 2004; Datovo & Landim, 2005; Sarmento-Soares et al., 2006; Campos-Paiva & Costa, 2007), of which four are restricted to the subterranean environment in the São Domingos karst area, Central Brazil presenting moderately to highly reduced eyes and melanic pigmentation: I. passensis Fernández & Bichuette, 2002, I. bambui Bichuette & Trajano, 2004, I. epikarsticus Bichuette & Trajano, 2004 and I. ramiroi Bichuette & Trajano, 2004.
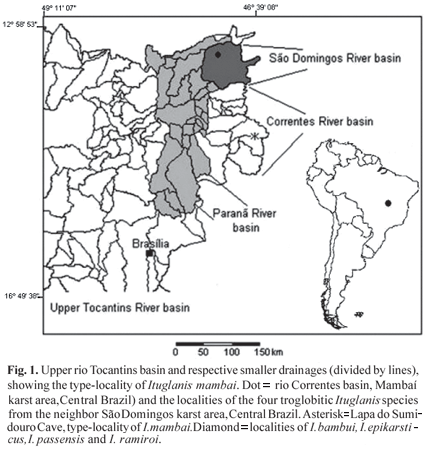
The Mambaí karst area, upper rio Tocantins basin, State of Goiás is located to south of the São Domingos area. The latter is distinguished by its subterranean ichthyofauna, particularly rich in both troglomorphic and non-troglomorphic species. It is the Brazilian region with the highest diversity of subterranean fishes, including seven troglobitic species reported until now (Bichuette & Trajano, 2003; Trajano et al., 2004). During two fieldtrips carried out in 2004 and 2007 to the Mambaí karst area, three populations of cave trichomycterids were found in caves. Differences in the external morphology and anatomy were observed in the large population found in the Lapa do Sumidouro Cave when comparing to other Ituglanis cave species and epigean congeners, justifying the recognition of a distinct species. The new Ituglanis species described herein is characterized by the high individual variability in the cutaneous pigmentation and slightly, but statistically significantly, reduced eyes.
Study area
Mambaí is a carbonate karst area characterized by the presence of continuous limestone outcrops belonging to the Bambuí Group. This area is located in the left margin of the rio Paranã, which is part of upper rio Tocantins basin (Fig. 1). Characteristically, after an epigean reach, each stream enters into a cave through a sinkhole, cross hundreds of meters (the largest cave in the region, Gruna da Tarimba, is approximately 7 km long) through subterranean conduits, and emerges to the surface through a resurgence. This area lies in the Cerrado (the savannah-like Brazilian vegetation) domain and is characterized by a tropical semi-humid climate with 4-5 dry mo/yr (Nimer, 1979).
The Lapa do Sumidouro (14º19'21.0"S 46º14'41.0"W) is a 2,000 m long cave, formed by many dry and higher galleries and crossed by a shallow stream (depth varying from 10 to 90 cm during our visits; mean width = 1.5 m), with a graveled and soft (silt and clay) bottom interspersed with collapsed boulders.
Material and Methods
The study specimens were hand-netted, anesthetized in benzocain solution until death, preserved in formalin and then transferred to alcohol 70%.
All measurements were straight-line, taken under stereomicroscope with a dial caliper, 0.1 mm precision, on the left side of specimens. Measurements follow Tchernavin (1944), de Pinna (1992), and Trajano & de Pinna (1996). Counts of dorsal-, anal-fin rays and vertebrae follow the method proposed by de Pinna (1992). Two specimens (unregistered) were cleared and double-stained for bone and cartilage by the method of Taylor & Van Dyke (1985). Osteological terminology follows de Pinna (1989).
We considered adults those individuals longer than 30.0 mm SL, which is the minimum size of specimens with matured gonads. Minimum population densities were estimated with basis on visual censuses of adult individuals, taking into account the area of the cave stream inspected (300 m long and 1.5 m wide, about 450 m2). The spatial distribution in the natural environment was described after ad libitum observations. Standard length and weight data of 12 adult individuals (paratypes) were used to calculate allometric condition factors (K= 100.W.SL-b; Le Cren 1951). The power coefficient (b) for the growth equation was estimated from the slope of the regression of log weight on log standard length using the pooled data.
Abbreviations: MCP, Museu de Ciências e Tecnologia da PUCRS, Porto Alegre; MZUSP, Museu de Zoologia da Universidade de São Paulo; LISDEBE, Laboratório de Ictiologia Sistemática do Departamento de Ecologia e Biologia Evolutiva da UFSCar; SL, standard length; HD, head depth; HL, head length; HW, head width; IOW, interorbital width; C&S, cleared and stained.
Ituglanis mambai, new species
Holotype. MCP 42538, 53.4 mm SL, Brazil, northeastern State of Goiás, Posse County, Lapa do Sumidouro Cave (14º19'21.0"S 46º14'41.0"W, elevation 792 m), upper rio Tocantins basin; M. E. Bichuette & S. Secutti col., 31 Mar 2007.
Paratypes. LISDEBE 2047, 6, 32.6-65.4 mm SL (C&S); same locality as holotype; M. E. Bichuette, E. Trajano & A. C. Barbosa col., 1 Sep 2004.MCP 42537, 3, 38.7-59.5 mm SL, same locality as holotype; M. E. Bichuette, E. Trajano & A. C. Barbosa col., 1 Sep 2004. MZUSP 94719, 4, 26.7-66.1 mm SL; same locality as holotype; M. E. Bichuette, E. Trajano & S. Secutti col., 31 March 2007.
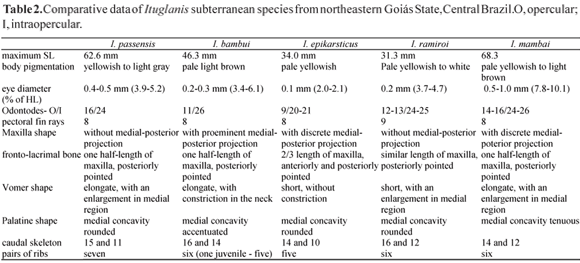
Diagnosis. Ituglanis mambai is distinguished from both epigean (I. cahyensis, I. laticeps, I. macunaima, I. metae, I. nebulosus, I. parahybae and I. proops) and cave congeners (I. bambui, I. epikarsticus, I. passensis and I. ramiroi) by the combination of the following characteristics: posterior fontanel absent (except for I. epikarsticus and I. macunaima); pectoral-fin rays usually i,7 (except for I. bambui, I. passensis and I. epikarsticus); six pleural ribs (except for I. bambui, I. parahybae and I. ramiroi); total vertebrae 37-38 behind Weberian apparatus (except for I. bambui, I. laticeps and I. macunaima,); shorter predorsal length (65.1-70.8% in SL) (except when compared to I. bambui); shorter caudal peduncle (8.4-11.9% in SL); shorter dorsal-fin base (7.7-11.3% in SL); larger interobital width (29.2-36.5% in HL) (except to the cave species); wider mouth (43.4-64.0% in HL) (except for I. bambui); pigmentation intermediary between epigean and cave Ituglanis species, composed by irregular light brown spots along the body; eyes size variable and intermediate between those of epigean and formerly described cave-restricted congeners, with a diameter varying, in adults, from 0.5 to 1.0 mm (7.8-10.1 % in HL) (except for I. cahyensis 7.0-8.1 % in HL). Additional diagnostic character states: the maxillae with a discrete medial-posterior projection; fronto-lacrimal one half-length of the maxillae and pointed backwards; posterior process of palatine half its length, with a tenuous medial concavity; 14 dorsal and 12 ventral procurrent rays.
Description. Morphometric and meristic data of holotype and paratypes given in Table 1. Body elongate, semi-cylindrical, becoming compressed toward caudal fin. Dorsal and ventral profiles of body, and caudal peduncle straight (Fig. 2a). Lips and barbels covered by papillae.
Head relatively wide and depressed, rounded in dorsal view. Eyes reduced in relation to epigean Ituglanis species, visible externally as round black spots, covered by thin skin (Fig. 2b). Anterior nostril transversally ovoid and slightly smaller than posterior one, surrounded laterally by nasal barbels. Posterior nostril rounded, surrounded anteriorly by thin flap of integument. Mouth subterminal, rictus laterally directed. When adpressed to body, maxillary barbel extending to middle of adpressed pectoral fin; submaxillary and nasal barbels extending to origin of pectoral fin.
Pectoral fin i,7, triangular in dorsal view, first ray longer, unbranched and filamentous. Dorsal fin ii,7, rectangular in lateral view. Anal fin i,5, semi-circular in lateral view, distal margin slightly rounded. Pelvic fin i,4, rectangular in ventral view. Margin of caudal fin straight, with 13 principal rays (7 in the upper and 6 in the lower lobe), 14 dorsal and 12 ventral procurrent rays.
General morphology of cranium: main body axis of mesethmoid with anterior width larger than posterior width, width of cornua 3/4 of its lenght, cornua with rounded distal extremity; posttemporosupracleithrum of medium size, with dorsal limb lying on pterotic. Two premaxillary tooth rows. Maxillae with discrete medial-posterior projection; fronto-lacrimal one half-length of maxillae and pointed backwards. Six branchiostegal rays. Base of laminar surface of urohyal four times longer than its distal extremity; urohyal dorsal process short. Length of posterior process of palatine half total palatine length, medial concavity tenuous. Vomer arrow-shaped with long posterior process. Opercle with 14-16 odontodes and interopercle with 24-26 odontodes.
Postcranial skeleton. Total vertebrae 37-38 behind Weberian apparatus, six pairs of ribs. Epural absent. Neural spine (n = 1) of preural centrum with round tip. Upper hypural plate triangular, lower hypural plate trapezoidal. Uroneural with acute distal extremity.
Color in alcohol. Pale yellowish to light brown in adults and pale yellowish in juveniles, with irregular light brown spots in lateral and dorsal views (Figs. 2a and 2b). Eye black. Fins poorly pigmented, showing translucent aspect in those individuals with pale pigmentation. Dorsal portion of head light brown.
Color in life. Body in general pale yellowish, irregular light brown spots along dorsum and flanks of body. Eyes black. Dorsal portion of head darker than rest of body. Juveniles less pigmented than adults, dorsal region of skull darker.
Distribution. Ituglanis mambai is known from a single subterranean stream inside the Lapa do Sumidouro Cave. This stream reach at least 500 m of extension.
Etymology. The specific name makes reference to the karst region , Mambaí, where the species occurs.
Notes on ecology and behaviour. Ituglanis mambai inhabits a subterranean stream inside the Lapa do Sumidouro Cave. The stream reach where the fishes were observed is characterized by slow to fast-moving waters, 10-90 cm deep on average, and bottom formed basically by sand, silt, some gravels and boulders. Visual censuses, carried out in two occasions and covering an area approximately 300 m long and 1.5 m wide, resulted in minimum population densities varying from 0.12 inds.m-2 (September 2004, end of the dry season) to 0.04 inds.m-2 (April 2007, beginning of the dry season). Environmental variables measured in September 2004: water temperature, 23.4°C; pH, 7.9; conductivity, 0.141 mSm.cm-1; dissolved oxygen, 7.4 mg.l-1. All observed individuals were solitary, with swimming activity on the bottom and sometimes in the midwater. In the natural habitat, I. mambai catfish displayed cryptobiotic habits, trying to hide into the gravels and under boulders when disturbed, apparently showing a negative response to carbide and flashlight. It was observed a preference to slow-moving pools. Recent flood marks were observed in the base-level stream conduit in March/April 2007 (end of rainy season), when less individuals were observed along the 300 m reach inspected, when compared to September 2004. The average condition factor for 12 individuals of I. mambai captured at the end of the dry season (September 2004) was 2.1, showing, for two juvenile individuals (33.0 and 35.9 mm SL), values of 0.5, and varying from 0.7 to 3.4, for adults (40.2-72.0 mm SL).
Discussion
Taxonomy. According to Costa & Bockmann (1993), Ituglanis is defined by three autapomorphies: supraoccipital fontanel reduced to a small round orifice, palatine with a deep medial concavity and anterior extremity of the sphenotic directed anteriorly. I. mambai presented some variation as regards to these characters. The supraoccipital fontanel was absent in two claired and stained individual, a condition shared with some individuals of I. epikarsticus and I. macunaima. Datovo & Landim (2005) discussed this character-state and concluded that this tendency (reduction to absence of supraoccipital fontanel) can be considered a derived condition. From this point of view, I. mambai could be a more derived species. In relation to the medial concavity of the palatine, we observed that it is not as deep in I. mambai as in the other species. Therefore, only the third synapomorphy (anterior extremity of the sphenotic directed anteriorly) is explicitly present, showing a mosaic of character-states.
Some hypotheses have been proposed to clarify the relationships within the genus. Pinna & Keith (2003) proposed two putatively monophyletic groups within Ituglanis, commented by Datovo & Landim (2005) and Sarmento-Soares et al. (2006). One monophyletic group includes northern South American species, which are distributed in the rio Amazonas and its left tributaries and in the Guyanas, sharing a a reduced number of pleural ribs (2-3) and, within this clade, a subgroup sharing the widening of the mesethmoid shaft. The second monophyletic group includes southern species, distributed from the basins of the Paraná/Paraguai, Ribeira do Iguape and Paraíba do Sul rivers, southeastern drainages in Brazil and Uruguay, and two southern Amazonian forms, as well as two undescribed forms from Uruguay and Tocantins river basins. These species share a large number of pleural ribs (5-7), and, within this clade,, a subgroup shares the medial process on the fronto-lacrimal tendom bone. In Ituglanis mambai, there is a narrow mesethmoid shaft, no process in the fronto-lacrimal tendom bone and 6 pleural ribs, which approach I. mambai to the second monophyletic group, not surprisingly in view of its occurrence in the upper Tocantins basin.
Ituglanis mambai may be distinguished from the epigean and cave Ituglanis species mainly by the classical troglomorphic character states (Christiansen, 1962): eyes reduced when compared to epigean ones and pigmentation intermediary between epigean and cave Ituglanis species. The troglobitic status of I. mambai is further supported by the fact that no specimen of Ituglanis has been found in the epigean streams in the area, in spite of extensive collecting efforts (Bichuette & Trajano, 2003). The only record of an epigean species near Mambaí and São Domingos karst areas (right margin of Paranã basin) is represented by Ituglanis sp. (MZUSP 53222), from a tributary of the upper rio Tocantins basin (left margin of the rio Paranã). Based on this, we hypothesize that the cave Ituglanis species from São Domingos karst area (I. bambui, I. epikarsticus, I. passensis and I. ramiroi), as well as I. mambai, are geographic relicts (sensu Holsinger, 1988), i.e., the subterranean survivors of one or more lineages previously living in epigean streams in this area. Table 2 summarizes the differences between I. mambai and the other known Ituglanis cave species, which are geographically close, showing a mosaic distribution of character states. Although most character states are shared by two or more species, they may all be distinguished from the others by a particular combination of morphometric and osteological characteristics. For instance, there is some overlapping in the eye diameter (in % of HL) of I. passensis, I. bambui and I. ramiroi but these species differ in the vomer shape.
Several authors argue that there is correlation between the time of isolation in the subterranean habitat and the degree of reduction of eyes and pigmentation, with populations showing less reduced and/or variable eyes and pigmentation isolated for shorter times than those homogeneously anophthalmic and unpigmented (Wilkens, 1982, 1986; Trajano, 1995). From this, it may be concluded that I. mambai, which present a low degree of regression of body pigmentation and eyes, showing variability in these character-state, has been isolated in the subterranean environment for a shorter time than, for example, I. bambui, I. passensis, I. ramiroi and I. epikarsticus. As a matter of fact, I. mambai exhibits cryptobiotic habits, occasionally swimming in the midwater, traits usually also observed in epigean species of Trichomyctyeridae, but not in the most specialized troglobitic ones. Nevertheless, this assumption must be regarded cautiously because many factors are involved in the differentiation rates of each character (Trajano, 2007).
Ecological and conservation remarks. For cavefish standards (Trajano, 2001), the population densities recorded by visual censuses on two occasions, respectively at the beginning and at the end of the dry season, may be considered low (0.04 inds.m-2, beginning of the dry season) to moderate (0.12 inds.m-2, end of the dry season). The lower number of individuals visualized at the beginning of the dry season may be due to floods on the previous rainy season, washing away some fish and/or to a tendency to remain sheltered until heavy rains ceased. Lowered condition factor values at the end of the dry season, similar to the observed for Ituglanis mambai were also reported for Pimelodella spelaea (Trajano et al., 2004), indicating a food-limited regime, intensified along the dry season due to an accentuated shortage of water-carried nutrients. The lack of specializations to cope with the food shortage is another evidence of a relatively short time in isolation in the subterranean habitat for both species. In relation to the power coefficient for the growth equation (b), the value of 3.23 indicates a positive-allometric growth (increase in relative body thickness or plumpness) and is close to those registered for other Brazilian cave catfishes (Trajano et al., 2004, Trajano & Bichuette, 2007).
Troglobitic species are generally fragile, highly vulnerable to environmental disturbance, due to their generally high degree of endemism, low population sizes, high sensitivity to stressors, and a K-selected life style leading to a slow population turnover (Trajano, 2000). As a matter of fact, many hypogean fishes are considered as endangered by different types of threats (Proudlove, 2001), among which human visitation cannot be neglected.
Mambaí is a poorly developed region, relatively well preserved, and there are no short- to medium-term big projects for the area. Ituglanis mambai presents a relatively large population, and the Lapa do Sumidouro Cave is not currently visited by tourists. Nevertheless, the fast growing speleotourism throughout Brazil, the relative proximity to a large city (Brasília, about 330 km far away), and the fact that these caves are not included in any conservation unit imposes some concern about the future of I. mambai in particular, as well as of the subterranean ecosystems in general situated in this region. The creation of a conservation unit encompassing all the Mambaí karst area is a highly recommendable preventive measure.
Comparative material. Ituglanis bambui, MZUSP 79860, holotype, Brazil: Goiás: São Domingos: Terra Ronca State Park: Angélica Cave. Ituglanis epikarsticus, MZUSP 79869, holotype, Brazil: Goiás: São Domingos: Terra Ronca State Park: São Mateus Cave. Ituglanis passensis: MZUSP 80097, 3, Brazil: Goiás: São Domingos, Passa Três Cave. Ituglanis proops: MZUSP 79576, 15, Brazil: Paraná: Cerro Azul, Ribeirão Bonito. Ituglanis ramiroi, MZUSP 79865, holotype, Brazil: Goiás: São Domingos: Terra Ronca State Park: São Bernardo Cave. Ituglanis sp.: MZUSP 53222, 6, Brazil: Goiás: Minaçú, tributary of Rio Tocantinzinho. Trichomycetrus bahianus: MZUSP 74655, 10, Brazil: Bahia: Livramento do Brumado, Rio Brumado. Trichomycterus itacarambiensis: MZUSP 50548, paratypes, 4, Brazil: Minas Gerais: Itacarambi, Olhos D'Água Cave. Trichomycterus sp.: MZUSP 60205, 10, Brazil: São Paulo: Barra do Turvo, Ribeirão Fria. Trichomycterus zonatus: MZUSP 68173, 20, Brazil: São Paulo: Cajati, Rio do Queimado. Due to difficulties to get additional loans from the Museu de Zoologia da USP, the comparison with other Ituglanis species (I. parahybae, I. metae, I. laticeps, I. cahyensis and I. nebulosus) was based on the literature (Costa & Bockmann, 1993; de Pinna & Keith, 2003; Datovo & Landim, 2005; Sarmento-Soares et al., 2006).
Acknowledgements
We are greatly indebted to the Centro de Estudo, Proteção e Manejo de Cavernas in the State of Goiás CECAV/GO: José A. O. Motta, for the information about these new catfishes and support during the fieldwork; Geovanna M.V. Rocha, José E. T. Alarcão, Luis R. da Silva and Marco A. Olive. To colleagues and cavers who helped us in the field: Adriana C. Barbosa, Sandro Secutti, Fernando P. L. Marques, Ricardo Pinto-da-Rocha and Ericson C. Igual. We are grateful to Osvaldo Oyakawa (MZUSP) for providing the loan of comparative material. This paper was benefited with the comments and criticism of two anonymous reviewers. IBAMA granted us permission for collections (license # 02001.005646/98-40). Financial support was given by FAPESP (# 03/00794-5) and partially by All Catfish Species Inventory (ACSI) project (NSF DEB-0315963). The junior author is partially supported by the CNPq (fellowship # 306066/88-2).
Literature Cited
Accepted February 2008
Published 31 March 2008
- Bichuette, M. E. & E. Trajano. 2003. Epigean and subterranean ichthyofauna from São Domingos karst area, upper Tocantins river basin, Central Brazil. Journal of Fish Biology, 63: 1100-1121.
- Bichuette, M. E. & E. Trajano. 2004. Three new subterranean species of Ituglanis from Central Brazil (Siluriformes: Trichomycteridae). Ichthyological Exploration of Freshwasters, 15(3): 243-256.
- Campos-Paiva, R. M. & W. J. E. M. Costa. 2007. Ituglanis paraguassuensis sp. n. (Teleostei: Siluriformes: Trichomycteridae): a new catfish from the rio Paraguaçu, northeastern Brazil. Zootaxa, 1471: 53-59.
- Christiansen, K. 1962. Proposition for the classification of cave animals. Spelunca, 2: 76-78.
- Costa, W J. E. M. & F. A. Bockmann. 1993. Un nouveau genre néotropical de la famille des Trichomycteridae (Siluriformes: Loricarioidei). Revue Française de Aquariologie, 20: 43-46.
- Datovo, A. & M. I. Landim. 2005. Ituglanis macunaima, a new catfish from the rio Araguaia basin, Brazil (Siluriformes: Trichomycteridae). Neotropical Ichythyology, 3(4): 455-464.
- Fernández, L. & M. E. Bichuette. 2002. A new cave dwelling species of Ituglanis from the São Domingos karst, central Brazil (Siluriformes: Trichomycteridae). Ichthyological Exploration of Freshwaters, 13: 273-278.
- Holsinger, J. R. 1988. Troglobites: the evolution of cave-dwelling organisms. American Scientist, 76: 146-153.
- Nimer, E. 1979. Climatologia do Brasil. SUPREN, Rio de Janeiro, Brasil, 421 p.
- de Pinna, M. C. C. 1989. A new sarcoglanidine catfish, phylogeny of its subfamily, and an appraisal of the phyletic status of the Trichomycterinae (Teleostei, Trichomycteridae). American Museum Novitates, 2950: 1-39.
- de Pinna, M. C. C. 1992. A new subfamily of Trichomycteridae (Teleostei, Siluriformes), lower loricarioid relationships and a discussion on the impact of additional taxa for phylogenetic analysis. Zoological Journal of the Linnean Society, 106: 175-229.
- de Pinna, M. C. C. 1998. Phylogenetic relationships of Neotropical Siluriformes (Teleostei: Ostariophysi); historical overview and synthesis of hypothesis. Pp. 279-330. In: L. Malabarba, R. Reis, R. P. Vari, C. Lucena & M. Lucena (Eds.). Phylogeny and classification of neotropical fishes. Edipucrs, Porto Alegre.
- de Pinna, M. C. C. & P. Keith. 2003. A new species of the catfish genus Ituglanis from French Guyana (Osteichthyes: Siluriformes: Trichomycteridae). Proceedings of the Biological Society of Washington, 116(4): 873-882.
- de Pinna, M. C. C. & W. Wosiacki. 2003. Trichomycteridae. Pp. 270-290, in Reis R. E., Kullander, S. O. & Ferraris, Jr. C. J. (eds.), Check list of the freshwater fishes of South and Central America. Edipucrs, Porto Alegre.
- Proudlove, G. S. 2001. The conservation status of hypogean fishes. Environmental Biology of Fishes, 62: 201-213.
- Sarmento-Soares, L. M., R. F. Martins-Pinheiro, A. T. Aranda & C. C. Chamon. 2006. Ituglanis cahyensis, a new catfish from bahia, Brazil (Siluriformes: Trichomycteridae). Neotropical Ichthyology, 4(3): 309-318.
- Sket, B. 1988. Speleobiological investigations in the Colombian Andes. Bioloski Vestnik, 36: 53-62.
- Taylor, W. R. & G. A. Van Dyke. 1985. Revised procedures for staining and clearing small fishes and other vertebrates for bone and cartilage study. Cybium, 9: 107-119.
- Tchernavin, V. 1944. A revision of some Trichomycterinae based on material preserved in the British Museum (Natural History). Proceedings of the Zoological Society of London, 114: 234-275.
- Trajano, E. 1995. Evolution of tropical troglobites: applicability of the model of Quaternary climatic fluctuations. Mémoires de Biospéologie, 22: 203-209.
- Trajano, E. 1997. Synopsis of Brazilian troglomorphic fishes. Mémoires de Biospéologie, 24: 119-126.
- Trajano, E. 2000. Cave faunas in the Atlantic tropical rain forest: Composition, ecology and conservation. Biotropica, 32:882-893.
- Trajano, E. 2001. Ecology of subterranean fishes: an overview. Environmental Biology of Fishes, 62: 133-160.
- Trajano, E. 2007. The challenge of estimating the age of subterranean lineages: examples from Brazil. Acta carsologica, 36(1): 191-198.
- Trajano, E. & M. E. Bichuette. 2004. Diversity of subterranean fishes in Brazil. Proceedings of the Symposium on World Subterranean Biodiversity, Villeurbanne, França. Pp. 65-65.
- Trajano, E. & M. E. Bichuette. 2007. Population ecology of cave armored catfishes, Ancistrus cryptophthalmus Reis, 1987, from Central Brazil (Siluriformes: Loricariidae). Ecology of Freshwater Fish, 16: 105-115.
- Trajano, E. & M. C. C. de Pinna. 1996. A new cave species of Trichomycterus from eastern Brazil (Siluriformes, Trichomycteridae). Revue Française de Aquariologie, 23: 85-90.
- Trajano, E., R. E. Reis & M. E. Bichuette. 2004. Pimelodella spelaea: a new cave catfish from Central Brazil, with data on ecology and evolutionary considerations. Copeia, 2004(2): 315-325.
- Wilkens, H. 1982. Regressive evolution and phylogenetic age: the history of colonization of freshwaters of Yucatan by fish and Crustacea. Texas Memorial Museum Bulletin, 128: 237-243.
- Wosiacki, W. 2002. Estudo das relações filogenéticas de Trichomycterinae (Teleostei, Siluriformes, Trichomycteridae) com uma proposta de classificação. Unpublished Ph. D. Dissertation, Instituto de Biociências, Universidade de São Paulo, São Paulo, Brazil, 324 p.
Publication Dates
-
Publication in this collection
14 Apr 2008 -
Date of issue
Mar 2008
History
-
Received
Feb 2008 -
Accepted
31 Mar 2008