Abstracts
A second inseminating species bearing club cells organized into an anal-fin gland in sexually active males belonging to the genus Bryconadenos was recently discovered in the rio Xingu drainage and is described as new. It is distinguished from B. tanaothoros by the presence of a conspicuous dark blotch at the humeral region, and fewer scale rows between dorsal-fin origin and anal-fin origin. Males of B. weitzmani have longer pelvic fins at sizes beyond 30 mm SL. Bryconadenos weitzmani is further distinguished from B. tanaothoros by the cup shape of the anal-fin gland, its lower part much more developed than the upper, whereas in B. tanaothoros there is just a slit separating the almost equally developed upper and lower parts.
Freshwater fish; South America; Neotropical; Insemination; Systematics; Taxonomy
Uma segunda espécie inseminadora do gênero Bryconadenos, possuindo células "club" organizadas em uma glândula na nadadeira anal de machos sexualmente ativos, foi recentemente descoberta na bacia do rio Xingu, é descrita como nova. Distingue-se da outra espécie do gênero, B. tanaothoros, pela presença de uma conspícua mancha umeral e menor número de escamas entre a origem da nadadeira dorsal e a origem da nadadeira anal. Machos de B. weitzmani têm nadadeiras pélvicas mais longas a partir de 30 mm SL. Bryconadenos weitzmani distingue-se ainda de B. tanaothoros por apresentar a glândula da nadadeira anal em forma de taça, com a porção ventral muito mais desenvolvida que a dorsal, enquanto que em B. tanaothoros as duas porções apresentam-se quase igualmente desenvolvidas e separadas por uma fenda.
A new species of Bryconadenos (Characiformes: Characidae) from the rio Curuá, rio Xingu drainage, Brazil
Naércio A. Menezes; André Luiz Netto-Ferreira; Katiane M. Ferreira
Museu de Zoologia da Universidade de São Paulo, Caixa Postal 42494, 04218-970 São Paulo, SP, Brazil. naercio@usp.br (NAM); alnferreira@gmail.com (ALNF)
ABSTRACT
A second inseminating species bearing club cells organized into an anal-fin gland in sexually active males belonging to the genus Bryconadenos was recently discovered in the rio Xingu drainage and is described as new. It is distinguished from B. tanaothoros by the presence of a conspicuous dark blotch at the humeral region, and fewer scale rows between dorsal-fin origin and anal-fin origin. Males of B. weitzmani have longer pelvic fins at sizes beyond 30 mm SL. Bryconadenos weitzmani is further distinguished from B. tanaothoros by the cup shape of the anal-fin gland, its lower part much more developed than the upper, whereas in B. tanaothoros there is just a slit separating the almost equally developed upper and lower parts.
Key words: Freshwater fish, South America, Neotropical, Insemination, Systematics, Taxonomy.
RESUMO
Uma segunda espécie inseminadora do gênero Bryconadenos, possuindo células "club" organizadas em uma glândula na nadadeira anal de machos sexualmente ativos, foi recentemente descoberta na bacia do rio Xingu, é descrita como nova. Distingue-se da outra espécie do gênero, B. tanaothoros, pela presença de uma conspícua mancha umeral e menor número de escamas entre a origem da nadadeira dorsal e a origem da nadadeira anal. Machos de B. weitzmani têm nadadeiras pélvicas mais longas a partir de 30 mm SL. Bryconadenos weitzmani distingue-se ainda de B. tanaothoros por apresentar a glândula da nadadeira anal em forma de taça, com a porção ventral muito mais desenvolvida que a dorsal, enquanto que em B. tanaothoros as duas porções apresentam-se quase igualmente desenvolvidas e separadas por uma fenda.
Introduction
Specimens of the species herein described, Bryconadenos weitzmani are from two localities in the rio Curuá, a stream flowing into rio Iriri, rio Xingu basin, Pará State, Brazil. Preliminary examination indicated the presence of a thick tissue on the anterior portion of the anal fin in which club cells were later identified, and other color features diagnostic of B. tanaothoros Weitzman, Menezes, Evers & Burns, but further detailed examination revealed significant differences with respect to the only known species of Bryconadenos and therefore a second species should be recognized and described. Histological sections undertaken through female ovaries revealed the presence of spermatozoa indicating that the new species is inseminating.
The discovery of another inseminating species belonging to incertae sedis genera within the Characidae primarily defined by sexual characters (Vari & Ortega, 2000; Malabarba & Weitzman, 2003; Weitzman et al., 2005; Burns & Weitzman, 2006) increases the source of information of such characters for a better understanding of relationships within the family.
Material and Methods
Counts, measurements and presentation of data are described in Fink & Weitzman (1974), Weitzman et al. (2005) and Menezes (2007). In the description the range of counts are presented first, followed by counts of the holotype and the mean. Meristic and morphometric data corresponding to Bryconadenos tanaothoros are from the holotype, MZUSP 85852 (rio Suiá-Miçu, Mato Grosso State at 12º50'90"S 52º07'46"W), and the following paratypes: MZUSP 62102 (tributary of rio Teles Pires, upper rio Tapajós basin, approximately 11º17'S 55º20'W), Mato Grosso State; MZUSP 79752 and LIRP 4087 (confluence of rio Cervo and córrego do Gato, upper rio Xingu basin at 13º03'32"S 55º02'12"W, Mato Grosso State; MCP 29467 (rio Ferro on the road between Novo Mato Grosso and Nova Ubiratã, upper rio Xingu basin at 13º03'32"S 55º2'12"W, Mato Grosso State). In species description the range of meristic characters is presented first, followed in parentheses by counts of the holotype and the mean of the sample. All counts are based on the examination of 31 specimens with exception of number of vertebrae (n = 19), horizontal scale rows between dorsal-fin origin and anal-fin origin (n = 30), predorsal scales (n = 30), lateral line scales (n = 20) and horizontal scale rows around caudal peduncle (n = 18). Vertebrae were counted from radiographs of specimens. All morphometric data expressed as percentages of standard length that showed statistically significant differences between males and females (in bold in Table 1) were treated through regression analysis. The t-test to estimate significance of mean differences between males and females was performed using the R Package (R Development Core Team, 2007). A difference was considered significant when p < 0.05. For histological analyses, tissues of gill, ovaries, testis and the anal-fin gland were removed from specimens initially fixed in 10% formalin and subsequently transferred to 70% ethanol. These tissues were hydrated again for five hours, and post-fixed overnight in 2% glutaraldehyde and 4% paraformoldehyde in 0.1 M Sorensen phosphate buffer, pH 7.2. In the sequence the tissues were dehydrated in an ethanol series, embedded in methacrylate, sectioned at 3 µm and stained with Hematoxylin and Eosin and Toluide Blue pH alkaline.
The specimens studied are deposited at the Museu de Zoologia, Universidade de São Paulo (MZUSP).
Results
Bryconadenos weitzmani , new species
Holotype. MZUSP 98666, 40.7 mm SL, female, Brazil, Pará State: Altamira, rio Curuá tributary of rio Iriri, rio Xingu basin, 08º43'50"S 54º57'49"W, 20 Oct 2007, J. L. O. Birindelli, L. M. Sousa, A. L. Netto-Ferreira, M. H. Sabaj-Perez & N. Lujan.
Paratypes. MZUSP 96558, 10, 32.0-41.3 mm SL, collected with holotype. MZUSP 96559, 20, 18.0-36.5 mm SL, Brazil: Pará State: Altamira, vila de Castelo dos Sonhos, tributary of rio Iriri, rio Xingu basin, 08º19'07"S 55º05'23"W, 22 Oct 2007, same collectors as holotype.
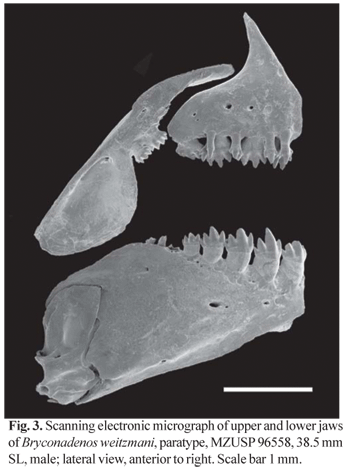
Diagnosis. Bryconadenos weitzmani can be distinguished from B. tanaothoros, the only known species of the genus by the presence of a conspicuous dark blotch at the humeral region (vs. absent in B. tanaothoros), by having fewer scale rows between dorsal-fin origin and anal-fin origin (7-8, mean = 7.03 vs. 8-10, mean = 8.8) and the anal-fin gland cup-shaped in mature males, its lower part much more developed than the upper (Fig. 2). The anal-fin gland in B. tanaothoros (Weitzman et. al. 2005, fig. 7, p. 339) is not cup-shaped, the upper and lower parts being almost equallly developed and sometimes separated by a slit in fully mature males.
Description. Morphometrics of holotype and paratypes are presented in Table 1. Body small reaching at least 41.3 mm SL, elongate and laterally compressed; greatest body depth slightly anterior to dorsal-fin origin at vertical crossing pelvic-fin origin.
Dorsal body profile convex at snout, gently convex from above nostril to dorsal-fin origin, straight or nearly so and posteroventrally inclined from this point to caudal peduncle. Dorsal profile of caudal peduncle slightly concave. Ventral body profile convex at tip of lower jaw, gently convex from below anterior margin of orbit to anal-fin origin, nearly straight and dorsally inclined along anal-fin base and slightly concave along ventral margin of caudal peduncle. Lower jaw somewhat included in upper jaw when mouth closed. Posterior tip of maxilla extending beyond vertical crossing anterior border of orbit.
Dorsal-fin rays ii,8 in all specimens. Posterior most ray split to its base. Adipose fin present. Anal-fin rays iv,17-19 (19), 17.5, posterior most ray split to its base. Strongly developed anterior anal-fin lobe including anterior unbranched rays and first 5-6 branched rays in both sexes. Anal fin of sexually mature males with bilateral bony hooks on anterior five branched rays. Hooks distributed on sides of anterior and posterior primary branches. In a cleared and stained male specimen (MZUSP 96558, 38.5 mm SL) hooks are not always present on both sides of anterior and posterior primary branches. 17 hooks on first branched ray, 9 on second, 8 on third, 5 on fourth, and 2 on fifth. Well-developed cup-shaped gland covering bases of five anterior most anal-fin rays. Pectoral-fin rays i,11-12 (i,11), 11.2. Distal tip of longest pectoral-fin ray not reaching pelvic-fin origin even in mature females, but almost reaching origin of that fin in males, but no differences found in relative lengths of pectoral fins in males and females through regression analysis. Pectoral-fin rays without hooks. Pelvic-fin rays i,7 in all specimens. No hooks on pelvic-fin rays of sexually mature males. Distal tip of longest pelvic-fin rays extending slightly beyond anal-fin origin. Pelvic-fin length of sexually mature specimens sexually dimorphic (Fig. 3). Principal caudal-fin ray count 10/9 in all specimens.
Scales cycloid. Lateral line complete, perforated scales 36-39(38), 37.8. Predorsal scales 10-12(11), 11.1. Scale rows between dorsal-fin origin and anal-fin origin 7-8(7), 7.03. Scale rows around caudal peduncle 13-15(14), 13.8.
Shape, size and arrangement of teeth on premaxilla, maxilla and dentary as described for Bryconadenos tanaothoros (compare Fig. 3, this paper, with fig. 9 in Weitzman et al., 2005, p. 341). Outer row small teeth on premaxilla 2-5(3), 3.3. Inner row premaxillary teeth 4 in all specimens. Maxillary teeth 2 in all specimens. Dentary with 4 large teeth followed more or less abruptly by 3-4(3), 3.3 smaller teeth. Dorsal limb gill rakers 5-7(6), 6.1; ventral limb gill rakers 10-13(11), 11.4 in first gill arch. Branchiostegal rays 4 in one cleared and stained specimen, 3 rays originating on anterior and 1 on posterior ceratohyal. Vertebrae 36-38(37), 37.1.
Color in alcohol. Body color of Bryconadenos weitzmani identical to that of B. tanaothoros as described by Weitzman et al. (2005, p. 341-342) with exception of: dark lateral stripe extending from behind dorsal part of opercle to caudal peduncle where it broadens, its posterior margin covering bases of 1-16 principal caudal-fin rays; medially on caudal fin stripe continues to distal tip of caudal-fin rays 9-12. Stripe widest above pelvic-fin origin, covering at his area three longitudinal scale rows. Conspicuous dark blotch slightly deeper than wide present at humeral region. Longitudinal dark stripe on dorsum extending from occiput to dorsal part of caudal peduncle separated by one and a half longitudinal scale rows from horizontal body stripe, ventrally interrupted on each scale immediately below median dorsal scale row by a small pale area.
Sexual dimorphism. The most obvious sexually dimorphic feature in Bryconadenos weitzmani is the presence of a cup-shaped anal-fin gland in sexually active males, not present in females. Histological sections revealed the presence of glandular club cells in the glandular tissue which apparently undergo holocrine secretion. Gill glands are present in sexually active males but not in females, and develop from anterior gill filaments of the first gill arches, consisting of five gill filaments and four chambers. The anterior anal-fin profile of males and females are slightly different (Fig. 1).
Estimation of slope differences between males and females indicated that only the relationship between pelvic-fin length and standard length was significant (difference between slopes = 0.115; standard error = 0.021; t = 5.121; p = 0.00002). Figure 4 suggests that significant differences between males and females start to appear at about the standard length of 30 mm.
Etymology. Bryconadenos weitzmani is named in honor of Stanley H. Weitzman for his outstanding contribution to the knowledge of neotropical freshwater fishes.
Distribution. This species is known from the rio Curuá, a tributary of rio Iriri, rio Xingu basin in Altamira, Pará State, Brazil (Fig. 5).
Ecological notes. Specimens of Bryconadenos weitzmani were collected from two stretches of the rio Curuá. Both localities were clear water rocky rapids, with thick sand on the bottom (Fig. 6). They were observed by one of us (ALN-F) forming small schools of about 10-15 individuals actively swimming against fast flowing waters below a one meter high fall. The new species seems to be very rheophylic, living in streams with fast flowing waters. Specimens of Baryancistrus sp. 1, Baryancistrus sp. 2, Caiapobrycon sp., Crenicichla of the group saxatilis, and Teleocichla sp. were collected below the same waterfall where part of the type specimens (MZUSP 96559) were found. Other 91 species were collected in rio Curuá at the collecting sites near the villages of Castelo dos Sonhos and Cachoeira da Serra, below large waterfalls near the latter which seem to represent an effective barrier isolating the headwater fish fauna from the species living below it (Birindelli et al., in press).
Discussion
The discovery of Bryconadenos weitzmani brings up to two the number of species known in this genus. It is very similar to B. tanaothoros, sharing with it characters 1 to 7 listed and described by Weitzman et al. (2005, p. 35), leaving no doubts about their close relationship. In Bryconadenos weitzmani the pterygiophores and respective medial radials of anterior five branched and last unbranched rays are fused, a condition absolutely identical to that in B. tanaothoros (Weitzman et al., 2005, fig. 5, p. 337). This and the presence of an anal-fin gland in males are exclusive features of the two Bryconadenos species as emphasized by Weitzman et al. (2005: 335-336) and distinguish them from all characids. In addition to the meristic difference pointed out above, there are clear differences in color patterns between the two species, the most conspicuous represented by the presence of a dark humeral blotch in B. weitzmani. In the color description of B. tanaothoros it is emphasized that no obvious humeral mark or blotch is present. The difference in size and shape of the anal-fin gland between the two species is also remarkable. In B. weitzmani the club cells are found in a cup-shaped gland involving at least five anterior branched anal-fin rays and having its lower part much more developed than the upper (Fig. 2). The anal-fin gland in B. tanaothoros is not cup-shaped, and just a slit can be sometimes distinguished separating the upper and lower parts of the gland in fully mature males (Weitzman et al., 2005, fig. 7, p. 339).
As in Bryconadenos tanaothoros (Weitzman et al., 2005: 348, fig. 15), males of B. weitzmani present spermatozoa with elongated nuclei, denominated intrasperm, and packs of spermatozoa were observed within the ovaries of females. The new species is therefore among the inseminating members of the Clade A of Malabarba & Weitzman.
Acknowledgements
All the specimens examined were collected during the PIPE Expedition to Serra do Cachimbo in Brazil in October 2007 funded by the U.S.A. National Science Foundation (NSF DEB 0315963) through the All Catfish Species Inventory Project. Collecting permits were issued by the Instituto Brasileiro do Meio Ambiente e dos Recursos Naturais Renováveis (processo IBAMA 02001.002040/99-61). The authors thank Irani Quagio-Grassioto (UNESP - Botucatu) for making available the facilities of her laboratory to carry out light and electron microscopy on Bryconadenos weitzmani. Stanley H. Weitzman read the manuscript and made useful comments. Eduardo G. Baena (MZUSP) took radiographs of specimens and prepared all the illustrations used in this paper. Assistance in the use of computer programs was provided by Rogério R. Silva (MZUSP) and in the use of the scanning electron microscope by Lara Maria Guimarães (MZUSP).
Literature Cited
Accepted April 2009
Published June 17, 2009
- Birindelli, J. L. O., A. M. Zanata, L. M. Sousa & A .L. Netto-Ferreira. 2009. New species of Jupiaba Zanata (Characiformes: Characidae) from Serra do Cachimbo, with comments on the endemism of upper Curuá, rio Xingu basin, Brazil. Neotropical Ichthyology, 7(1): 11-18.
- Burns, J. R. & S. H. Weitzman. 2006. Intromittent organ in the genus Monotocheirodon (Characiformes: Characidae). Copeia, 2006(3): 529-534.
- Fink, W. L. & S. H. Weitzman. 1974. The so-called cheirodontin fishes of Central America with descriptions of two new species (Pisces: Characidae). Smithsonian Contributions to Zoology, 172: 1-46.
- Malabarba, L. R. & S. H. Weitzman. 2003. Description of a new genus and six new species from Southern Brazil, Uruguay and Argentina with a discussion of a putative characid clade (Teleostei: Characiformes: Characidae). Comunicações do Museu de Ciências e Tecnologia, PUCRS, Série Zoologia, 16(1): 67-151.
- Menezes, N. A. 2007. A new species of Cynopotamus Valenciennes, 1849 (Characiformes: Characidae) with a key to species of the genus. Zootaxa, 1635: 55-61.
- Vari, R. P. & H. Ortega. 2000. Attonitus a new genus of sexually dimorphic characiforms (Ostariophysi: Characidae) from western Amazonia; a phylogenetic definition and description of three new species. Ichthyological Exploration of Freshwaters, 11(2): 113-141.
- Weitzman, S. H., N. A. Menezes, H-G. Evers & J. R. Burns. 2005. Putative relationships among inseminating and externally fertilizing characids, with a description of a new genus and species of Brazilian inseminating fish bearing an anal-fin gland in males (Characiformes: Characidae). Neotropical Ichthyology, 3(3): 329-360.
Publication Dates
-
Publication in this collection
24 July 2009 -
Date of issue
June 2009
History
-
Accepted
17 June 2009 -
Received
Apr 2009