Abstracts
A new species of Tatia is described from the lower rio Iguaçu basin, Paraná State, Brazil. The new species is distinguishable from congeners by having large, irregular pale blotches over dark brown base coloration, absent longitudinal stripe, caudal fin with round to striate dark brown spots over light base in adults or entirely dark in juveniles, and by having a long humeral process. Sexual dimorphism of the new species is marked by differences among genital and urinary apertures, size proportions between upper and lower lobe of caudal fin, and size, width and presence of antrorse and retrorse spines on anal-fin rays.
Neotropical; Freshwater fish; Sexual dimorphism; Catfish
Uma nova espécie de Tatia é descrita da bacia do baixo rio Iguaçu, Estado do Paraná, Brasil. A nova espécie é diagnosticada de suas congêneres por apresentar grandes manchas claras e irregulares sobre um fundo marrom escuro, faixa longitudinal ausente, nadadeira caudal com manchas arredondadas a estrias marrons escuras sobre uma base clara nos adultos ou completamente negra nos jovens, e por apresentar o processo umeral longo. O dimorfismo sexual da nova espécie é marcado por diferenças entre as aberturas genitais e urinárias, proporções de tamanho entre o lobo superior e inferior da nadadeira caudal e tamanho, largura e presença de espinhos antrorsos e retrorsos nos raios da nadadeira anal.
A new Tatia (Ostariophysi: Siluriformes: Auchenipteridae) from the rio Iguaçu basin, Paraná State, Brazil
Carla Simone PavanelliI; Alessandro Gasparetto BifiII
INupélia, Universidade Estadual de Maringá, Av. Colombo, 5790, 87020-900 Maringá, PR, Brazil. carlasp@nupelia.uem.br
IIPrograma de Pós-Graduação em Ecologia de Ambientes Aquáticos Continentais, Universidade Estadual de Maringá, Av. Colombo, 5790, 87020-900 Maringá, PR, Brazil. agbifi@hotmail.com
ABSTRACT
A new species of Tatia is described from the lower rio Iguaçu basin, Paraná State, Brazil. The new species is distinguishable from congeners by having large, irregular pale blotches over dark brown base coloration, absent longitudinal stripe, caudal fin with round to striate dark brown spots over light base in adults or entirely dark in juveniles, and by having a long humeral process. Sexual dimorphism of the new species is marked by differences among genital and urinary apertures, size proportions between upper and lower lobe of caudal fin, and size, width and presence of antrorse and retrorse spines on anal-fin rays.
Key words: Neotropical, Freshwater fish, Sexual dimorphism, Catfish.
RESUMO
Uma nova espécie de Tatia é descrita da bacia do baixo rio Iguaçu, Estado do Paraná, Brasil. A nova espécie é diagnosticada de suas congêneres por apresentar grandes manchas claras e irregulares sobre um fundo marrom escuro, faixa longitudinal ausente, nadadeira caudal com manchas arredondadas a estrias marrons escuras sobre uma base clara nos adultos ou completamente negra nos jovens, e por apresentar o processo umeral longo. O dimorfismo sexual da nova espécie é marcado por diferenças entre as aberturas genitais e urinárias, proporções de tamanho entre o lobo superior e inferior da nadadeira caudal e tamanho, largura e presença de espinhos antrorsos e retrorsos nos raios da nadadeira anal.
Introduction
The genus Tatia was proposed by Miranda-Ribeiro (1911) who included two species: T. intermedia (Steindachner) and T. aulopygia (Kner). Recently Sarmento-Soares & Martins-Pinheiro (2008) diagnosed Tatia among the Centromochlinae by having the hyomandibula anterodorsally elongated, not contacting the narrow metapterygoid and, instead, connected to the trapezoidal quadrate, reduced anal-fin base in adult males, and compressed and deep caudal peduncle, with a middorsal keel posterior to adipose fin. Those authors also recognized 12 valid species widespread in cis-Andean South American basins.
Previous inventories in the rio Iguaçu basin (e. g. Garavello et al., 1997; Ingenito et al., 2004; Baumgartner et al., 2006) have recorded only two species of Centromochlinae, Glanidium ribeiroi Haseman, and a new species of Tatia. Many Tatia specimens also have been collected during frequent samples in the lower rio Iguaçu basin over the last few decades by the Núcleo de Pesquisas em Limnologia, Ictiologia e Aqüicultura (Nupélia), of the Universidade Estadual de Maringá. Tatia neivai has been recorded from the whole Paraná-Paraguay system, except the Iguaçu basin, where occurs a different and undescribed species referable to the genus Tatia (sensu Sarmento-Soares & Martins-Pinheiro, 2008). This species exhibits a unique color pattern among congeners and it is described herein as a new Tatia.
Material and Methods
Counts follow Soares-Porto (1995) and measurements Sarmento-Soares & Buckup (2005) with the latter made on the left side of the specimens whenever possible, with calipers to the nearest 0.1 mm. Measurements of the subunits of the head are presented as percent of head length (HL) and those related to the body, including HL and subunits of the body as percent of standard length (SL). Meristic data included vertebrae, caudal-fin and procurrent rays which were taken from six male and six female specimens cleared and stained (c&s) according to procedures of Taylor & Van Dyke (1985). Five males and two females were also radiographed for helping osteological analyses. Vertebrae counts follow Sarmento-Soares & Martins-Pinheiro (2008). Counts from the holotype are indicated by an asterisk. Material is listed informing, in parentheses, the number of specimens in the lot from which counts and measurements were taken if that number is less than the total number of specimens in the lot, and their range of SL. Institutional abbreviations are CZCEN, Colección Zoológica de la Facultad de Ciencias Exactas y Naturales, Universidad Nacional de Asunción, DZSJRP, Departamento de Zoologia e Botânica, São José do Rio Preto, MNRJ, Museu Nacional, Rio de Janeiro, MZUEL, Museu de Zoologia da Universidade Estadual de Londrina, MZUSP, Museu de Zoologia da Universidade de São Paulo, NUP, Coleção Ictiológica do Nupélia, Maringá, besides collectors Copel, Companhia Paranense de Energia, and Gerpel, Grupo de Pesquisas em Recursos Pesqueiros e Limnologia.
Tatia jaracatia, new species
Tatia sp. Luiz et al., 2005: 183 (checklist, Paraná State, Brazil).Baumgartner et al., 2006: 2 (checklist, Salto Osório Reservoir, lower rio Iguaçu basin, Brazil).
Holotype. MZUSP 98248, male, 65.7 mm SL, Brazil, Paraná State, Municípios de Salto do Lontra and Dois Vizinhos, lower rio Iguaçu basin, rio Jaracatiá, 25º38'17"S 53º17'04"W, 25 Nov 1998, Nupélia staff.
Paratypes. All lots from Brazil, Paraná State, lower rio Iguaçu basin, collector Nupélia staff (unless otherwise stated). Municípios de Boa Esperança do Iguaçu and Nova Prata do Iguaçu, rio Jaracatiá, 25º37'37"S 53º16'52"W: MZUSP 98249, 2, 46.5-48.0 mm SL, 8 Aug 1997; NUP 4936, 4, 38.1-46.0 mm SL (1, 46.0 mm SL), 24 Nov 1998; NUP 4937, 1, 50.8 mm SL, 25 Jan 1999; NUP 5327, 1 c&s, 46.9 mm SL, 25 Jan 1999; NUP 5329, 3 c&s, 44.3-51.8 mm SL, 24 Nov 1998. Municípios de Boa Vista da Aparecida and Três Barras do Paraná, rio Tormenta, 25º25'23"S 53º21'09"W: NUP 4958, 3, 31.4-58.5 mm SL (1, 51.6 mm SL), 19 Jan 1999. Municípios de Capitão Leônidas Marques and Realeza (Marmelândia), Salto Caxias Reservoir, upstream from the dam, 25º31'55"S 53º29'01"W: MNRJ 31909, 1, 42.9 mm SL, 21 Jan 1999; MZUSP 98250, 2, 51.5-55.6 mm SL, 5 and 6 Feb 1998; MZUSP 98251, 1, 52.2 mm SL, 21 Jan 1999; MZUSP 98252, 1, 45.4 mm SL, 3 Feb 2001; MZUSP 98253, 1, 63.0 mm SL, 6 Mar 2000; NUP 4947, 2, 44.0-66.3 mm SL (1, 66.3 mm SL), 5 and 6 Jun 1997; NUP 4948, 2, 47.5-49.4 mm SL (1, 49.4 mm SL), 28 Jun 2000; NUP 4950, 2, 60.3-72.2 mm SL, 5 and 6 Feb 1998; NUP 4951, 1, 49.4 mm SL, 21 Jan 1999; NUP 4959, 2, 62.8-64.4 mm SL, 4 Apr 1997; NUP 5330, 6 c&s, 40.0-52.2 mm SL, 21 Jan 1999. Municípios de Capitão Leônidas Marques and Realeza (Marmelândia), Salto Caxias Reservoir, downstream from the dam, 25º31'55"S 53º29'01"W: NUP 4957, 1, 63.5 mm SL, 17 Dec 1998. Município de Cruzeiro do Iguaçu, rio Chopim, 25º34'27"S 53º05'49"W: MZUSP 98254, 1, 27.4 mm SL, 27 Nov 1999. Município de Quedas do Iguaçu, Salto Osório Reservoir, 25º30'-25º37'S 52º43'-53º01'W: NUP 2654, 4, 49.3-53.1 mm SL, Jun 2001, Copel staff; NUP 3243, 5, 42.7-56.8 mm SL, 7 Nov 2003, Gerpel staff; NUP 4175, 1, 54.0 mm SL, 9 Dec 2003, Gerpel staff; NUP 4176, 1, 48.9 mm SL, 25º30'49"S 53º00'049"W, 5 Dec 2004, Gerpel. Município de Realeza, rio Cotejipe, 25º34'35"S 53º30'06"W: NUP 4956, 1, 55.3 mm SL, 16 Dec 1998. Municípios de Salto do Lontra and Dois Vizinhos, rio Jaracatiá, 25º38'17"S 53º17'04"W: MNRJ 31910, 1, 57.0 mm SL, 13 Feb 1997; NUP 4931, 1, 57.5 mm SL, 11 Mar 1997; NUP 4932, 1, 56.2 mm SL, 8 Oct 1997; NUP 4933, 2, 51.8-53.9 mm SL (1, 53.9 mm SL), 25 Nov 1998. Município de Três Barras do Paraná, rio Adelaide, 25º27'18"S 53º18'26"W: MNRJ 31912, 1, 54.1 mm SL, 20 Nov 1998; NUP 4954, 4, 54.8-58.7 mm SL (1, 57.0 mm SL), 20 Nov 1998; NUP 4955, 1, 52.2 mm SL, 13 Dec 1998; NUP 5328, 1 c&s, 49.7 mm SL, 20 Nov 1998. Municípios de Três Barras do Paraná and Nova Prata do Iguaçu, rio Iguaçu, close to Porto Vorá, 25º32'24"S 53º18'08"W: MNRJ 31511, 2 ex., 51.2-54.2 mm SL, 10 Jan 1998; NUP 4953,1, 45.9, 6 Mar 2000; NUP 5326, 1 c&s, 43.2 mm SL, 10 Jan 1998.
Non-measured paratypes. All lots from Brazil, Paraná State, lower rio Iguaçu basin, collector Nupélia staff. Municípios de Boa Esperança do Iguaçu and Nova Prata do Iguaçu, rio Jaracatiá, 25º37'37"S 53º16'52"W: NUP 1630, 2, 41.8-52.1 mm SL, 4 Oct 2000; NUP 4935, 1, 42.8 mm SL, 7 Jan 2001. Municípios de Capitão Leônidas Marques and Realeza, Salto Caxias Reservoir, 25º31'55"S 53º29'01"W: NUP 4986, 1, 52.2 mm SL, 1 Nov 2002, Copel staff; NUP 4987, 3, 48.8-56.1 mm SL, Feb 1997 to Jan 2000. Municípios de Capitão Leônidas Marques and Realeza (Marmelândia), Salto Caxias Reservoir, upstream from the dam, 25º31'55"S 53º29'01"W: NUP 4946, 1, 38.0 mm SL, 3 Oct 2000. Município de Cruzeiro do Iguaçu, rio Chopim, 25º34'27"S 53º05'49"W: NUP 1743, 1, 37.9 mm SL, 25 Nov 2000; NUP 1799, 6, 28.2-31.5 mm SL, 27 Nov 1999. Municípios de Três Barras do Paraná and Nova Prata do Iguaçu, rio Iguaçu, close to Porto Vorá, 25º32'24"S 53º18'08"W: NUP 707, 10, 35.2-48.0 mm SL, 10 Jan 1998; NUP 4952,1, 47.3 mm SL, 3 Nov 2000. Município de São Jorge do Oeste, Salto Osório Reservoir, 25º36'28"S 52º52'45"W: NUP 4988, 1, 55.1 mm SL, 22 Nov 2006, Gerpel staff.
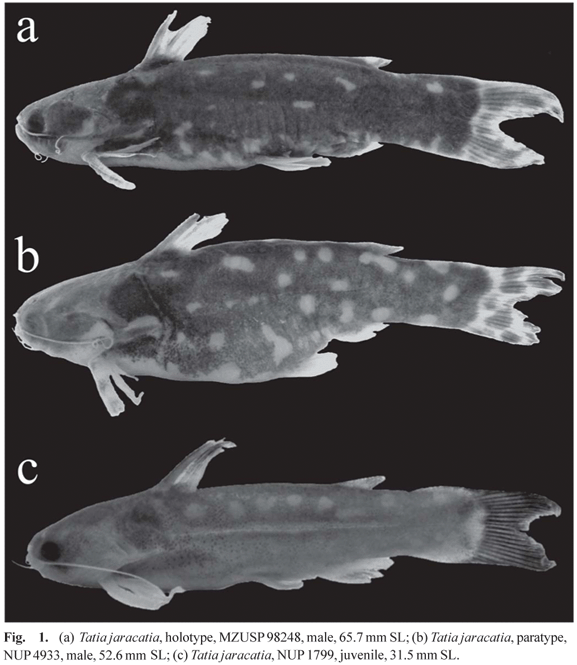
Diagnosis. Tatia jaracatia can be distinguished from its congeners, except T. aulopygia, T. brunnea, T. dunni, T. galaxias, T. intermedia, and T. neivai, by having large pale blotches over a dark brown base coloration (vs. body with dark brown blotches or more or less uniformly dark in T. boemia, T. caxiuanensis, T. meesi, T. nigra, dark brown base with pale stripes in T. strigata, and body with dark brown midlateral stripe in T. gyrina). Tatia jaracatia can be distinguished from T. aulopygia by having anal-fin rays of mature males uniformly decreasing in size from third unbranched ray (vs. anal fin of mature males distinctly notched). Tatia jaracatia have large and irregular pale dots over a dark brown base (vs. regular, longitudinally elongated or roundish, and very small in T. galaxias and T. neivai, and longitudinally more elongate pale blotches in T. dunni). Moreover, juvenile individuals of T. jaracatia possess uniformly dark brown caudal fin (vs. caudal fin uniformly pale, dotted or striped in T. aulopygia, T. intermedia and T. neivai). Furthermore T. jaracatia have 31 or 32 post-Weberian vertebrae (vs. more than 35 in T. aulopygia, T. dunni and T. intermedia) and 9 or 10 ribs (vs. 7 in T. galaxias). Some specimens of T. jaracatia have brown caudal-fin dots arranged as stripes, which might be confused with pale striae to roundish dots on caudal fins of some T. neivai. However, T. jaracatia has three to five (mode = 4, n = 11) hemal spines with thicker distal portion, while T. neivai has two or three (mode = 2, n = 3), in addition to different color pattern of body above mentioned. Tatia jaracatia can be diagnosed from T. intermedia by presenting caudal fin uniformly dark brown or with several dark brown chromatophores irregularly widespread, forming roundish blotches, and rarely forming stripes, and the humeral process long, reaching or surpassing the vertical line through origin of dorsal fin (vs. caudal fin with the same color pattern of the body, and humeral process shorter, not reaching that line).
Description. Morphometric data are presented in Table 1. Body elongated and head slightly depressed. Dorsal profile of body slightly convex from snout tip to adipose fin and somewhat concave on the anterior half of the caudal peduncle, then convex to the caudal-fin rays. Ventral profile of body faintly convex from mandibulary symphysis to pelvic fin and slightly concave from this point to caudal peduncle.
Thin integument over dorsal portion of head, allowing discernible skull roof. Fontanel elliptical and narrow, surrounded by frontals and entering mesethmoid. Third nuchal plate projecting laterally. Mouth terminal, slightly prognathous, possessing three irregular rows of conic premaxillary and dentary teeth. Maxillary barbel occasionally surpassing tip of humeral process. Inner mental barbel on the average approximately 70% of the length of outer mental barbel. Lateral line complete and inconspicuous, more perceptible in juveniles and c&s specimens.
Dorsal-fin rays I,5* (n = 40); dorsal-fin spine with 11 to 18 (15*, mode = 16, n = 33) serrations distributed along anterior edge. Pectoral-fin rays I,4* (n = 41), pectoral-fin spine with 13 to 27 (23*, mode = 22, n = 36) antrorse serrations along anterior edge and 9 to 18 (18*, mode = 16, n = 36) retrorse serrations along posterior edge. Pelvic-fin rays i,5* (n = 43), with second branched ray longest. In males, anal-fin rays iii,6* (n = 18); females ii,7 (n = 23). Forked caudal-fin rays i,7/8,i* (n = 16) and 16 to 20 (mode = 17, n = 14) upper procurrent rays and 15 to 19 (mode = 16, n = 14) lower. Post-Weberian vertebrae 31 (n = 7) or 32 (n = 9), and nine (n = 16) or ten (n = 1) pairs of ribs; last vertebra with vestigial ribs preceded by one without ribs.
Color in alcohol. Base color of dorsal and lateral surfaces of head and trunk ranging from light to dark brown; base color of ventral region of head and trunk white, lacking dark chromatophores. Lateral surface of body having pattern of pale blotches ranging from large to small, round to oval, and arranged longitudinally, transversally, or non-uniformly (Fig. 1). Dorsal fin generally hyaline, with scattered dark chromatophores on central or distal portion of first and second branched rays. Pectoral fin hyaline. Pelvic and anal fins hyaline, darkly colored only close to the base. Caudal fin ranging from uniformly dark brown, chiefly in juveniles (Fig. 1c), to having brown asymmetrical or oval blotches, or also with diffuse dark chromatophores irregularly arranged, occasionally forming stripes.
Sexual dimorphism. Adult females of Tatia jaracatia have genital and urinary apertures separate, but very close together; and proximal radials of anal fin not fused. Adult males have genital and urinary openings separate as well, but the former is close to the first anal-fin ray origin, and the latter is at vertical line through pelvic-fin tip; upper lobe of caudal fin longer than inferior; three to five* hemal spines above proximal radials distally thicker; first unbranched anal-fin ray reaching about a half length of the second, third unbranched anal-fin ray longer and thicker than second and with two or three* antrorse spines along anterior edge of distal segments, and first branched anal-fin ray with two retrorse spines along anterior portion of median segments (Fig. 2). Proportions between branched and unbranched fin rays differ among sexes as well, as mentioned above.
Distribution. The new species is known from the lower rio Iguaçu basin, upstream from the Iguaçu falls, Southern Brazil (Fig. 3).
Etymology. The species name, jaracatia, refers to the type-locality, rio Jaracatiá, lower rio Iguaçu basin. Jaracatiá is an indigenous name given to a fructiferous tree common in the region. A noun in apposition.
Discussion
Tatia jaracatia fits all given characters for the genus stated by Sarmento-Soares & Martins-Pinheiro (2008), but exhibits a peculiar trunk color pattern composed of conspicuous, irregular, pale blotches over a dark background.
Most species of Tatia (sensu Sarmento-Soares & Martins-Pinheiro, 2008) are distributed across the Amazon, Tocantins, and Orinoco rivers, and coastal drainages of the Guiana Shield. The only two more southerly distributed species are T. boemia and T. neivai from the Uruguay and Paraná rivers, respectively. These last two species share with T. jaracatia the absence of ribs on vertebra preceding the last rib-bearing vertebrae (Soares-Porto, 1998). This putative osteological synapomorphy suggests that all three Tatia species in the La Plata basin are monophyletic.
Given its apparent absence from the entire rio Paraná basin below Iguaçu Falls, T. jaracatia appears to be endemic to the rio Iguaçu above the falls. The rio Iguaçu above Iguaçu Falls has been described by several authors as relatively depauperate in overall fish diversity, but high in fish endemism, presumably due to isolating effects of the falls (Garavello et al., 1997; Júlio Jr. et al., 1997; Baumgartner et al., 2006). Several other new species putatively endemic to the region above the falls are currently in press or in preparation by us and other researchers, and we predict that further collecting effort in the headwaters of rio Iguaçu will likely yield additional new, endemic taxa.
Material examined (range of SL is given in millimeters). Brazil. Centromochlus altae. MZUEL 1749, 6, 28.7-35.2, córrego Salobro, Serra das Araras, Mato Grosso State; NUP 3113, 3, 21.9-30.0, Manso Reservoir, rio Cuiabá, rio Paraguai basin, Mato Grosso State. C. perugiae. MZUSP 31880, 1, 28.1, rio Tarauacá, affluent of rio Juruá, rio Solimões basin, Acre State. C. schultzi. MZUSP 49189, 1, 46.6, rio Fontoura, rio Xingu basin, Pará State; MZUSP 86997, 1, 52.3, rio Culuene, affluent of rio Xingu, Mato Grosso State. Glanidium cesarpintoi. DZSJRP 4570,1, 90.6, Salto Grande Reservoir, upper rio Paraná basin, São Paulo State. G. melanopterum. MZUSP 51275, 10 of 24, 82.0-100.8, rio Ribeira de Iguape, São Paulo State. G. ribeiroi. NUP 5467, 127, 49.6-77.8, Salto Caxias Reservoir, lower rio Iguaçu basin, Paraná State. Tatia aulopygia. MZUSP 37764, 1, 61.3, igarapé affluent of rio Aripuanã, Aripuanã, Mato Grosso State. T. cf. brunnea. MZUSP 84977, 5 of 35, 52.5-60.0, rio Tiquié, Amazonas State. T. caxiuanensis. MZUSP 47503, 1 c&s, 28.6, igarapé affuent of rio Capim, Pará State. T. cf. dunni. MZUSP 6643, 1, 65.6, igarapé of lago Manacapuru, Manacapuru, Amazonas State. T. galaxias. MZUSP 57269, 1, 45.4, rio Jutuaí, Amazonas State. T. gyrina. MZUSP 84979, 1, 30.5, rio Tiquié, Amazonas State. T. intermedia. MZUSP 30584, 3 of 5, 69.1-76.1, rio Uraricuera, Roraima State; MZUSP 31881, 2, 35.7-37.7, igarapé do Cujobim, Rio Branco, Roraima State. MZUSP 44127, 1, 86.8, rio Pauini, Amazonas State. T. neivai. NUP 363, 2, 40.0, ribeirão São Pedro, upper rio Paraná basin, Paraná State; NUP 971, 17, 19.7-51.9, rio Palmeiras, rio Paraguai basin, Mato Grosso State; NUP 1106, 48, 28.6-72.2, rio do Peixe, affluent of rio Corumbá, upper rio Paraná basin, Goiás State; NUP 2077, 2, 55.0-52.0, rio Piquiri, upper rio Paraná basin, Paraná State; NUP 2080, 2, 45.4-63.5, Itaipu Reservoir, upper rio Paraná basin, Paraná State; NUP 2687, 4, 60.0-61.8, Chavantes Reservoir (rio Paranapanema), upper rio Paraná basin, Paraná State; NUP 2696, 4, 50.8-58.6, Canoas II Reservoir (rio Paranapanema), upper rio Paraná basin, Paraná State; NUP 2706, 1, 67.5, Salto Grande Reservoir (rio Paranapanema), upper rio Paraná basin, Paraná State; NUP 3799, 2, 47.5-63.3, rio Pirapó, upper rio Paraná basin, Paraná State; NUP 4225, 3, 59.8-68.4, rio Goioerê, upper rio Paraná basin, Paraná State; NUP 4260, 1, 42.0, rio Cuiabá, rio Paraguai basin, Mato Grosso State; NUP 4422, 4, 41.3-49.6, Rosana Reservoir (rio Paranapanema), upper rio Paraná basin, São Paulo State; NUP 5331, 1 c&s, 60.0, rio do Peixe, affluent of rio Corumbá, upper rio Paraná basin, Goiás State; NUP 5332, 1 c&s, 43.1, rio Palmeiras, rio Paraguai basin, Mato Grosso State; NUP 5949, 2, 57.6-63.0, rio Laranjinha, affluent of rio Tibaji, upper rio Paraná basin, Paraná State. Paraguay. T. neivai. CZCEN 314, 6, 59.6-65.2, arroyo Ytú, affluent of río Piribebuy, río Paraguay basin, Cordillera. Venezuela. T. aulopygia. MZUSP 44072, 2, 68.1-70.3, Caño Cocuiza, affluent of río Orinoco, Bolívar.
Acknowledgements
We thank Weferson J. da Graça (UEM) for clearing and staining specimens besides valuable discussions, Mário de Pinna and Osvaldo Oyakawa (MZUSP) for loaning comparative material and offering radiographic facilities, José L. Birindelli (MZUSP) for providing some important bibliographic references, and Francisco Langeani (DZSJRP) and Oscar Shibatta (MZUEL) for loaning specimens. Collections were made by the Gerpel (Grupo de Pesquisas em Recursos Pesqueiros e Limnologia), and the Nupélia (Núcleo de Pesquisas em Limnologia, Ictiologia e Aqüicultura), which also provided logistical support. Research associated with this study was supported by the Copel (Companhia Paranense de Energia) and UEM (Universidade Estadual de Maringá)/Nupélia. Both authors have been supported by grants from CNPq (Conselho Nacional de Desenvolvimento Científico e Tecnológico), and are participants of the ACSI (All Catfish Species Inventory) Project, financed by the National Science Foundation-USA.
Literature Cited
Accepted March 2009
Published June 17, 2009
- Baumgartner, D., G. Baumgartner, C. S. Pavanelli, P. R. L. Silva, V. A. Frana, L. C. Oliveira & M. R. Michelon. 2006. Fish, Salto Osório Reservoir, rio Iguaçu basin, Paraná State, Brazil. Check List, 2(1): 1-4.
- Garavello, J. C., C. S. Pavanelli & H. I. Suzuki. 1997. Caracterização da ictiofauna do rio Iguaçu. Pp. 61-84. In: Agostinho, A. A. & L. C. Gomes (Eds.). Reservatório de Segredo: bases ecológicas para o manejo. Maringá, Eduem, 387p.
- Ingenito, L. F. S., L. F. Duboc & V. Abilhoa. 2004. Contribuição ao conhecimento da ictiofauna da bacia do alto rio Iguaçu, Paraná, Brasil. Arquivos de Ciências Veterinárias e Zoologia da Unipar, 7(1): 23-36.
- Júlio Jr., H. F., C. C. Bonecker & A. A. Agostinho. 1997. Reservatório de Segredo e sua inserção na bacia do rio Iguaçu. Pp. 1-17. In: Agostinho, A. A. & L. C. Gomes (Eds.). Reservatório de Segredo: bases ecológicas para o manejo. Maringá, Eduem, 387p.
- Luiz, E. A., A. C. Petry, C. S. Pavanelli, H. F. Júlio Jr., J. D. Latini & W. M. Domingues. 2005. As assembléias de peixes de reservatórios hidrelétricos do Estado do Paraná e bacias limítrofes. Pp. 169-184. In: Rodrigues, L., S. M. Thomaz, A. A. Agostinho & L. C. Gomes (Eds.). Biocenoses e reservatórios. Padrões espaciais e temporais. São Carlos, Rima, 321p.
- Miranda-Ribeiro, A. 1911. Fauna brasiliense, Peixes, Tomo IV (A) [Eleutherobranchios, Aspirophoros]. Arquivos do Museu Nacional, 16: 1-504.
- Sarmento-Soares, L. M. & P. A. Buckup. 2005. A new Glanidium from the rio São Francisco basin, Brazil (Siluriformes: Auchenipteridae: Centromochlinae). Copeia, 2005(4): 846-853.
- Sarmento-Soares, L. M. & R. F. Martins-Pinheiro, 2008. A systematic revision of Tatia (Siluriformes: Auchenipteridae: Centromochlinae). Neotropical Ichthyology, 6(3): 495-542.
- Soares-Porto, L. M. 1995. A new species of Tatia from the Amazon basin (Siluriformes: Auchenipteridae). Ichthyological Exploration of Freshwaters, 6(3): 201-206.
- Soares-Porto, L. M. 1998. Monophyly and interrelationships of the Centromochlinae (Siluriformes, Aucheniperidae). Pp. 331-350. In: Phylogeny and classification of Neotropical fishes. L. R. Malabarba, R. E. Reis, R. P. Vari, Z. M. S. Lucena & C. A. S. Lucena (Eds.). Porto Alegre, Edipucrs, 603p.
- Taylor, W. R. & G. C. Van Dyke. 1985. Revised procedures for staining and clearing small fishes and other vertebrates for bone and cartilage study. Cybium, 9(2): 107-119.
Publication Dates
-
Publication in this collection
15 July 2009 -
Date of issue
June 2009
History
-
Accepted
17 June 2009 -
Received
Mar 2009