Abstracts
This study investigated the variation in abundance, distribution and composition of ichthyoplankton in a lagoon in southern Brazil through the year and at different stations within the lagoon. Ibiraquera Lagoon is a shallow coastal lagoon connected to the sea by a semi-permanent sandbar. Ichthyoplankton samples were collected monthly from December 2003 to December 2004, with a 200 µm mesh net using horizontal surface hauls. A colder, drier period was detected, contrasting with warm months during which salinity varied depending on rainfall and whether the sandbar was open or closed. The mean abundance of ichthyoplankton over the study period was 459.6 ± 76.2 (SE) eggs · 100 m-3 and 63.6 ± 7.9 (SE) larvae · 100 m-3, and they were present during all months. Significantly higher abundances of eggs and larvae were observed during warm months. The community was dominated by the family Engraulidae followed by Clupeidae, Gobiidae and Mugilidae. Engraulidae and Clupeidae were present throughout the year, with greater abundances in months with higher temperatures. Some coastal fish species spawn in Ibiraquera Lagoon, mainly near to the sandbar, demonstrating that the lagoon is a spawning area for coastal stock. We recommend the establishment of environmental procedures to promote the conservation of Ibiraquera Lagoon and its ichthyoplankton community.
Ichthyoplankton; Estuaries; Spawning and Nursery places
A abundância, distribuição e composição do ictioplâncton foram investigadas em uma lagoa no sul do Brasil durante um ano e em diferentes estações dentro da lagoa. A lagoa de Ibiraquera é uma lagoa costeira rasa, conectada ao mar através de uma barra semi-permanente. As amostras de ictioplâncton foram coletadas mensalmente entre dezembro de 2003 e dezembro de 2004, com uma rede cônico-cilíndrica de 200 µm de malha em arrastos horizontais e de superfície. Foi detectado um período frio e seco em contraste aos meses quentes e com variação na salinidade em função das chuvas e da barra estar aberta ou fechada. A abundância média do ictioplâncton, durante o período de estudo, foi de 459,6 ± 76,2 (SE) ovos · 100 m-3 e 63,6 ± 7,9 (SE) larvas · 100 m-3, e ocorreram em todos os meses. As abundâncias de ovos e larvas foram significativamente maiores nos meses quentes. A comunidade foi dominada pela família Engraulidae seguida pelas famílias Clupeidae, Gobiidae e Mugilidae. As famílias Engraulidae e Clupeidae ocorreram principalmente nos meses com maiores temperaturas. Alguns peixes costeiros desovam dentro da lagoa, principalmente próximo a barra, evidenciando que a lagoa é uma área de desova para o estoque costeiro. Recomenda-se o estabelecimento de ações ambientais que promovam a conservação da lagoa de Ibiraquera e da comunidade ictioplanctônica.
Spatial and temporal distribution of fish eggs and larvae in a subtropical coastal lagoon, Santa Catarina State, Brazil
Luis C. P. Macedo-Soares; Alesandra B. Birolo; Andrea S. Freire
Laboratório de Crustáceos/Plâncton, ECZ, Universidade Federal de Santa Catarina, 88040-970 Florianópolis, SC, Brazil. luiscpms@yahoo.com.br, andreasfreire@yahoo.com.br
ABSTRACT
This study investigated the variation in abundance, distribution and composition of ichthyoplankton in a lagoon in southern Brazil through the year and at different stations within the lagoon. Ibiraquera Lagoon is a shallow coastal lagoon connected to the sea by a semi-permanent sandbar. Ichthyoplankton samples were collected monthly from December 2003 to December 2004, with a 200 µm mesh net using horizontal surface hauls. A colder, drier period was detected, contrasting with warm months during which salinity varied depending on rainfall and whether the sandbar was open or closed. The mean abundance of ichthyoplankton over the study period was 459.6 ± 76.2 (SE) eggs · 100 m-3 and 63.6 ± 7.9 (SE) larvae · 100 m-3, and they were present during all months. Significantly higher abundances of eggs and larvae were observed during warm months. The community was dominated by the family Engraulidae followed by Clupeidae, Gobiidae and Mugilidae. Engraulidae and Clupeidae were present throughout the year, with greater abundances in months with higher temperatures. Some coastal fish species spawn in Ibiraquera Lagoon, mainly near to the sandbar, demonstrating that the lagoon is a spawning area for coastal stock. We recommend the establishment of environmental procedures to promote the conservation of Ibiraquera Lagoon and its ichthyoplankton community.
Key words: Ichthyoplankton, Estuaries, Spawning and Nursery places.
RESUMO
A abundância, distribuição e composição do ictioplâncton foram investigadas em uma lagoa no sul do Brasil durante um ano e em diferentes estações dentro da lagoa. A lagoa de Ibiraquera é uma lagoa costeira rasa, conectada ao mar através de uma barra semi-permanente. As amostras de ictioplâncton foram coletadas mensalmente entre dezembro de 2003 e dezembro de 2004, com uma rede cônico-cilíndrica de 200 µm de malha em arrastos horizontais e de superfície. Foi detectado um período frio e seco em contraste aos meses quentes e com variação na salinidade em função das chuvas e da barra estar aberta ou fechada. A abundância média do ictioplâncton, durante o período de estudo, foi de 459,6 ± 76,2 (SE) ovos · 100 m-3 e 63,6 ± 7,9 (SE) larvas · 100 m-3, e ocorreram em todos os meses. As abundâncias de ovos e larvas foram significativamente maiores nos meses quentes. A comunidade foi dominada pela família Engraulidae seguida pelas famílias Clupeidae, Gobiidae e Mugilidae. As famílias Engraulidae e Clupeidae ocorreram principalmente nos meses com maiores temperaturas. Alguns peixes costeiros desovam dentro da lagoa, principalmente próximo a barra, evidenciando que a lagoa é uma área de desova para o estoque costeiro. Recomenda-se o estabelecimento de ações ambientais que promovam a conservação da lagoa de Ibiraquera e da comunidade ictioplanctônica.
Introduction
Temporal and spatial variations of fish abundances in estuaries can be the result of changes in physical conditions that influence species' seasonal movements (Vieira, 1991; Barletta et al., 2003; Spach et al., 2003). Indeed, studies of ichthyoplankton carried out in Brazil have shown that fish larvae abundance, distribution and composition may have temporal and spatial patterns that are caused by physical factors such as rainfall and salinity (Barletta-Bergan et al., 2002a, 2002b) and temperature (Muelbert & Weiss, 1991; Castro et al., 2005).
Those studies of ichthyoplankton have focused on mangroves, bays and continental shelves rather than coastal lagoons (e. g. Ekau et al., 1999; Tovar-Faro & Bonecker, 2000; Barletta-Bergan et al., 2002a; Franco & Muelbert, 2003; Freitas & Muelbert, 2004; Castro et al., 2005). However, coastal lagoons are also considered nursery areas for coastal fish (Sánchez-Velasco et al., 1996; Avendaño-Ibarra et al., 2004).
Life cycles of many commercial species are related to estuaries (Castro & Bonecker, 1996; Esper et al., 2001; Franco & Muelbert, 2003; Freitas & Muelbert, 2004). The southern Brazilian continental shelf is one of the most productive fishing areas of Brazil's 8,500 km coast. In Santa Catarina State, artisanal mullet fishing is an important economic and cultural activity among local communities of fishermen living in estuarine areas (Peterson et al., 2008). In the same way, mullet, pink shrimp and blue crab are the most important artisanal fishery resource in Ibiraquera Lagoon (Seixas & Berkes, 2003; Bonetti et al., 2005). Despite its natural characteristics and tourist importance, Ibiraquera Lagoon has been poorly studied, therefore there are no specific biological data to subsidize management plans for the area (Bonetti et al., 2005).
This aim of this study is to test the hypothesis that the variability in abundance and composition of fish eggs and larvae is homogeneous throughout the year and in a spatial gradient in the lagoon. Our findings were also reported at the Local Agenda 21 Forum to help improve the management initiatives in the area.
Material and Methods
Study area
The Ibiraquera Lagoon is located on the southern Brazilian coast between 28°06'1" and 28°10'1" S and between 48°37'44" and 48°41'53" W (Fig. 1). It is about 9 km long with a total area of 8,700 km2, and the local depth ranges from 0.2 m to 2.0 m with a sandy bottom (Seixas & Berkes, 2003). According to Kjerfve (1994) it is a "shallow shocked" lagoon, with a high surface/volume ratio and restricted water exchange with the adjacent ocean. It is geomorphologically segmented into four sectors known locally as lagoa de Cima, lagoa do Meio, lagoa de Baixo and Lagoa do Saco (Bonetti et al., 2005). These four sectors are interconnected by small channels and the largest of them (Cima) has an area of 4,000 km2. Baixo and Saco are the sectors closest to the adjacent coastal region, and they are separated from the sea by a semi-permanent sandbar that is 150 m wide.
When the sandbar is closed long enough to rise the water level due to freshwater inputs and rainfall events, water pressure opens the sandbar naturally (Seixas & Berkes, 2003). Nowadays, the sandbar is also opened artificially twice a year and the time of opening is dependent on the influence of local political groups. This opening causes significant changes in all areas of the Ibiraquera Lagoon, with increased salinity, even in the innermost sector (Bonetti et al., 2005). In this study, the four sectors of Ibiraquera Lagoon will be referred to as Cima, Meio, Baixo and Saco, or simply as the stations.
Freshwater input is restricted to small streams at Baixo and Saco, and conditioned mainly by rainfall. The input of salty water and the reduction of the water level during the opening of the sandbar, associated with the local wind, improve the water mixture and therefore the nutrients and oxygen distribution in the Lagoon (Bonetti et al., 2005). The area is under the intermediary subtropical zone (humid mesothermic group).
Sampling methods
Monthly ichthyoplankton samples were collected from December 2003 to December 2004, except for May and September 2004. The sandbar was opened from December 2003 to February 2004 and from June to August 2004. Three replicates were taken at each station (Cima, Meio, Baixo and Saco), with the exception of the January and February 2004 samples at Saco, and the June and July 2004 samples at Cima, due to shallow depth, comprising a total of 124 samples. Samples were collected with a 200 µm mesh net equipped with a General Oceanics flowmeter, during surface horizontal hauls lasting 5 minutes each.
The mean volume of water filtered was 46.79 m3. Samples were fixed in 4% buffered formaldehyde and ichthyoplankton were sorted from the entire sample using Olympus SZ40 and Carl Zeiss ® Stemi DV4 stereomicroscopes. Egg and larva abundances were standardized according to the number collected per 100 m3 of water filtered. Larvae were identified according to Lippson & Moran (1974), Moser et al. (1983), Leis & Trnski (1989), Neira et al. (1998), Ré (1999) and Richards (2001).
Water temperature was measured using a thermometer (precise to 0.5 ºC) and salinity (in 0.5 increments) was measured with a refractometer at the surface, at the same time as the plankton was sampled. For months without ichthyoplankton samples, temperature and salinity were obtained from the team conducting the main project in the area. Data on local rainfall were obtained from EPAGRI (Empresa de Pesquisa Agropecuária e Extensão Rural de Santa Catarina S.A) station, located in Urussanga (28º31'S 49º19'W), Santa Catarina State.
Data analysis
Dominance and frequency of occurrence were determined in order to identify the most important fish larvae families in the Ibiraquera Lagoon. Dominance was calculated for each family using the mean abundance of the family in relation to the total mean larvae abundance. The frequency of occurrence was calculated from the number of samples in which each family was caught as a proportion of the total number of samples.
Two-way ANOVA was used to determine whether there were any significant differences in temperature, salinity or ichthyoplankton density for different months and stations. Missing data were estimated from the mean of other replicates. Ichthyoplankton data were log(x+1) transformed. The Bartlett test was applied in order to verify the homogeneity of variances before each analysis and Tukey's multiple comparison test was used whenever significant differences were detected (Zar, 1996). The tests were performed on Statistica© 5.0 (Statsoft Inc., 1984-1995). Graphs were plotted to illustrate the ANOVA interactions for families with high dominance and/or with commercial value.
Cluster analysis was conducted to evaluate similarities between samples in terms of the distribution of families. Samples were clustered using the ranked Bray Curtis index according to mean fish larvae densities at each station (n = 42). Hierarchical group-average linking was carried out using PRIMER© 5.1 routines (PRIMER-E). All data were log (x+1) transformed.
Results
Hydrographic conditions
The water temperature characterized two periods during the year (Fig. 2a): warm months, with mean temperatures greater than 25 °C (January, February, March, November, and December 2004), and cold months with mean temperatures below 20 °C (June and July 2004). The interaction between months and stations can be explained by the significantly higher (p < 0.01) mean temperature at Saco in October, November and December 2004, and at Cima in June and July 2004.

When the sandbar was opened in December 2003, the salinity increased at all stations until March (Fig. 2b). After September 2004, when the bar was closed, the salinity decreased as the months passed. The results observed between April and August 2004 can be explained with reference to the ANOVA interactions. In April, July and August 2004 the salinity was significantly higher (p < 0.01) at Baixo, while in May and June the salinity was significantly higher at Saco.
The variation of rainfall, during the study period, characterized months with rainfall above 200 mm (December 2003, May and September 2004), and months with rainfall less than 100 mm (February, June, July, August, and October 2004) (Fig. 2c).
Analysis of the hydrographic conditions characterized a cold and dry period (June to August 2004), with high salinity at stations near the sandbar. During the warm months, salinity varied depending on rainfall and whether the sandbar was open or closed.
Ichthyoplankton distribution
During the entire study a total of 3,741 fish larvae were identified, while 25,702 eggs were sorted from the samples. Mean ichthyoplankton abundances in the Ibiraquera Lagoon were 459.6 ± 76.2 (SE) eggs · 100 m-3 and 63.6 ± 7.9 (SE) larvae · 100 m-3. The highest mean egg abundance (5,784.1 eggs · 100 m-3) was obtained in January and the highest mean larval abundance (512.9 larvae · 100 m-3), in November 2004 (Fig. 3). These values were obtained respectively at Cima and Meio Lagoon. High mean abundances of fish eggs were observed during all months, especially December 2003 and January 2004. The lowest mean abundance of eggs was during June 2004. Fish larvae were also observed every month, with lower mean abundance between June and August 2004 (Fig. 3).
The ANOVA showed the interaction between months and stations in relation to the distribution of eggs (Table 1). The mean abundance of eggs in Meio, Baixo and Saco followed the general pattern seen in Fig. 3. The ANOVA's interaction was explained by the reverse pattern of Cima, with significantly lower (p < 0.01) mean abundances in April, July, November and December 2004.
Composition of fish larvae
Engraulid larvae had the highest mean abundance, dominance and frequency of occurrence (Table 2 and Fig. 4). The second most abundant family was Clupeidae followed by Gobiidae, Mugilidae and Sciaenidae. Other families, such as Syngnathidae, Atherinopsidae and Achiridae had frequencies of occurrence greater than 10%, although with low abundance and dominance. Only engraulid, clupeid and syngnathid larvae were found in samples taken during all months of the year.
The dominance of the main families in the ichthyoplankton community changed over different months (Fig. 5). The Engraulidae family was dominant in December 2003 and August 2004 and shared dominance with Gobiidae in January 2004 and with Clupeidae in June 2004. Clupeid larvae were also dominant in November and December 2004, while the larvae of Gobiidae dominated in March and July 2004. In February 2004, Sciaenidae was the dominant family, while in October 2004 Mugilidae was dominant.
Clupeid larvae were present every month and at significantly higher abundances in December 2004 and at Baixo. The abundance of blenniid larvae was also significantly higher (p < 0.01) in December 2004, while syngnathid larvae had a significantly higher (p < 0.05) abundance at Saco (Table 2).
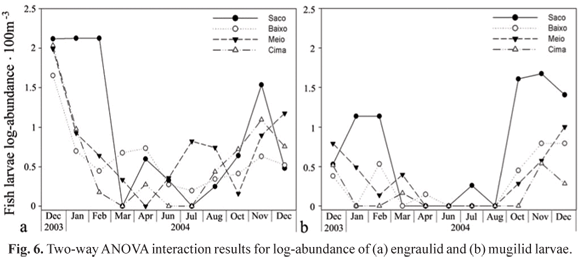
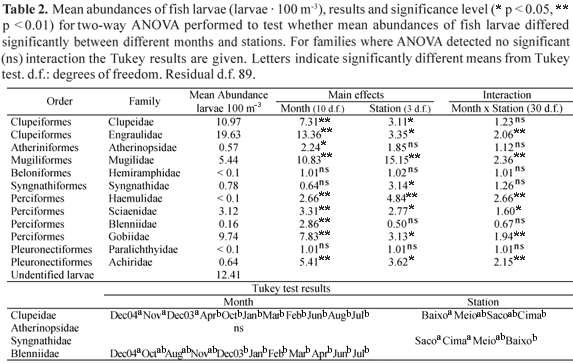
The ANOVA results demonstrated a significantly higher (p < 0.01) abundance of engraulid larvae at Saco from December 2003 to February 2004, and at Meio during the cold and dry period. There was also a significantly higher abundance at Baixo during March and April 2004 (Fig. 6a). There were significantly higher abundances of mugilid larvae during October and November 2004 at Saco, and in December 2003 and March 2004 at Meio (Fig. 6b). In December 2003, gobiid larvae were significantly more abundant distributed at Saco and Meio, while their abundance was significantly higher at Baixo and Cima during March 2004. In February 2004, sciaenid and achirid larvae were significantly more abundant at Meio. Moreover, in March 2004 haemulid larvae had significantly higher (p < 0.01) abundance at Baixo.
The cluster analysis identified two groups (Fig. 7) distinguished by hydrographic conditions and seasonal abundance of fish larvae. In both groups, engraulid and clupeid larvae were the most abundant. Group A was characterized by the cold and dry season, with low larvae abundance (mean of 8.21 larvae · 100 m-3), and syngnathid, atherinopsid, sciaenid, gobiid and blenniid larvae were also registered. The warm months were clustered in group B, where high larvae abundances were obtained (mean of 71.44 larvae · 100 m-3), with gobiid and mugilid larvae also being abundant.
Discussion
Salinity is strongly influenced by rainfall and dynamics of the sand bar, and characterizes Ibiraquera Laggon as a mixohaline laggon (Bonetti et al., 2005). Temperature variation follows the pattern found in subtropical latitudes (Muelbert & Weiss, 1991; Lüchmann et al., 2008), and the rainfall pattern is similar to that recorded at Conceição Lagoon (Lüchmann et al., 2008). Freshwater input was mainly influenced by rainfall, since Ibiraquera Lagoon has few small freshwater sources (Bonetti et al., 2005).
Mean eggs abundance ranges in Ibiraquera Lagoon were comparable with coastal areas from Bahia to Santa Catarina State (Franco & Muelbert, 2003; Freitas & Muelbert, 2004; Mafalda Jr. et al., 2004) and the rio de la Plata estuary (Berasategui et al., 2004). The mean abundance of fish larvae was higher than along the adjacent coast (Franco & Muelbert, 2003; Freitas & Muelbert, 2004) and than in the rio de la Plata estuary (Berasategui et al., 2004), but was lower than the channel area of laguna dos Patos (Muelbert & Weiss, 1991). Due to the small dimensions of Ibiraquera lagoon in relation to the systems cited above and its abundance of ichthyoplankton, we can consider it a spawning and nursery area.
Ichthyoplankton composition was similar to other Brazilian estuarine and coastal systems (Muelbert & Weiss, 1991; Castro & Bonecker, 1996; Ekau et al., 1999; Tovar-Faro & Bonecker, 2000; Barletta-Bergan et al., 2002a; Barletta-Bergan et al., 2002b; Joyeux et al., 2004; Mafalda Jr. et al., 2004; Castro et al., 2005; Chagas et al., 2006). The number of ichthyoplankton families (12) found in the area was similar to the number of adult fish families sampled, although Haemulidae, Blenniidae and Syngnathidae were not captured as adults (Ferreira & Freire, unpublished data).
Like in other estuarine systems (Muelbert & Weiss, 1991; Sánchez-Velasco et al., 1996; Tovar-Faro & Bonecker, 2000; Barletta-Bergan et al., 2002a; Joyeux et al., 2004; Castro et al., 2005) the community was dominated by engraulids and clupeids. Gobiid larvae, the third most abundant family in Ibiraquera Lagoon, were also important in the ichthyoplankton community in the southern Gulf of Mexico (Sánchez-Velasco et al., 1996) and in Vitória Bay (Joyeux et al., 2004). In the channel area of laguna dos Patos, Caeté estuary and Guanabara Bay, ichthyoplankton communities were also dominated by Sciaenidae (Muelbert & Weiss, 1991; Barletta-Bergan et al., 2002a; Barletta-Bergan et al., 2002b; Castro et al., 2005), which was the fifth most abundant family in the present study. All those examples corroborated the fact that these families use inner coastal systems during their life cycle, despite of its dimension and water exchange mechanism with the sea.
In coastal areas, down to 100 m depth, engraulid and clupeid larvae make up 60% of the total catch (Katsuragawa et al., 2006). These families are known to be estuarine residents, estuarine dependents or marine visitors (Garcia & Vieira, 2001) and can be found in the lagoon due to the dynamics of the sandbar. Engraulidae and Clupeidae exhibited higher abundance in the months when the sandbar was closed. Since their life cycle is short, up to 45 days for Sardinella brasiliensis reared in the laboratory (Silva et al., 2003; Katsuragawa et al., 2006), they could be breeding and spawning in the lagoon.
In the present study, Mugilidae abundance was higher than in the channel area of laguna dos Patos, Vitória Bay and Guanabara Bay (Muelbert & Weiss, 1991; Joyeux et al., 2004; Castro et al., 2005). Mugilids occur along the whole south Brazilian coast, Mugil curema and Mugil platanus being dominant in the southern lagoons (Figueiredo & Menezes, 1985; Garcia & Vieira, 2001; Ferreira & Freire, pers. comm.). Spawning migrations of mugilids usually refer to the movement of ripe fish from their feeding areas (generally estuaries) to the open sea (Ibáñez & Benítez, 2004). Nevertheless, mugilid larvae enter the estuary to complete their development and they are considered estuarine dependent (Figueiredo & Menezes, 1985; Garcia & Vieira, 2001). Early mugilid larvae, produced from recent spawn in the coast, could enter the lagoon during the months when the sandbar is open. On the other hand, larval mugilid stages reared in the laboratory completed their development between 29 and 89 days, depending on species (Kuo et al., 1973; Boglione et al., 1992; Yoshimatsu et al., 1992; Ismail et al., 1998; Monteiro-Ribas & Bonecker, 2001). Therefore, abundance of early mugilid larvae in the lagoon after 3 and 4 months without the bar being open, could only be explained by spawning inside the lagoon.
In Ibiraquera Lagoon, warm months had higher egg abundances, in common with the northeast coast of Brazil (Mafalda et al., 2004). The highest egg abundances were found at the stations with the highest abundance of brachyuran zoeae (Freire, pers. comm.). According to Freitas & Muelbert (2004) a partial overlap between zooplankton and fish egg distribution suggests a synchrony between spawning and the availability of food for the future larvae.
Abundance and richness of fish larvae were also higher in warm months in Ibiraquera lagoon, which is comparable with the channel area of laguna dos Patos (Muelbert & Weiss, 1991), Guanabara Bay (Tovar-Faro & Bonecker, 2000; Castro et al., 2005), the Magdalena-Almejas Bay lagoon system (Avendaño-Ibarra et al., 2004) and Lima estuary (Ramos et al., 2006). In those estuaries, temperature increases are a controlling factor for spawning and recruitment periods (Phonlor, 1984), determining temporal patterns of ichthyoplankton composition, distribution and abundance. In contrast with this pattern, in estuaries located in lower latitudes (Sánchez-Velasco et al., 1996; Barletta-Bergan et al., 2002a, 2002b), it is salinity and freshwater inputs that induce increases in fish larvae abundance (Barletta-Bergan et al., 2002b). Fluvial discharges related to the rainfall increase, nutrient concentration and plankton biomass increase, results in the spawning of many fishes during this period (Sánchez-Velasco et al., 1996).
The most dominant families, Engraulidae and Clupeidae, were present throughout the year (Muelbert & Weiss, 1991), mainly in the warm months (Fig. 5). Clupeids can spawn all year round, mostly in the late spring and summer, as recorded for Sardinella brasiliensis which mainly spawn in December 2003 and January 2004 along the south-eastern coast (Magro et al., 2000; Katsuragawa et al., 2006). In Guanabara Bay, Clupeidae dominance was also associated with the warm and rainy months, while Engraulidae were present during the greater part of the sampling period (Tovar-Faro & Bonecker, 2000). Gobiidae larvae were also present in Ibiraquera Lagoon most of the year, and peaked during months when rainfall increased. The same pattern was detected in the southern Gulf of Mexico (Sánchez-Velasco et al., 1996), where gobiid species spawn in the rainy period, and the opposite was observed in the channel area of laguna dos Patos (Muelbert & Weiss, 1991), where gobiid larvae were only present during early spring and late summer.
The spatial distribution of fish larvae demonstrates that they were well distributed across all stations. In Patos Lagoon, the densities of larvae from the Engraulidae, Clupeidae, Sciaenidae, Gobiidae and other families found in the Ibiraquera Lagoon were uniformly distributed throughout the channel area (Muelbert & Weiss, 1991). Similarly, in Vitória Bay, spatial variation in ichthyoplankton assemblage was not very pronounced, because some species occupy all places at all times (Chagas et al., 2006). However, Saco and Baixo exhibited a high abundance of some important fisheries and ecological groups. Generally, an area has been called a nursery if a juvenile fish or invertebrate species occurs at higher abundances, avoids predation more successfully, or grows faster there than in a different habitat (Beck et al., 2001). Since Saco and Baixo are the closest stations to the sea and have the highest salinity, the larvae of coastal and estuarine dependent fish could be arriving from the sea while the sandbar is open.
Ichthyoplankton distribution and composition were strongly influenced by environmental conditions, mainly the temperature pattern, which is to be expected at subtropical latitudes (Muelbert & Weiss, 1991; Barletta-Bergan et al., 2002b). Some families, such as Engraulidae, occurred throughout the year with variations in abundance. However, important fish such as mugilids peaked in October 2004, due to their reproductive period (Esper et al., 2001). The fact that economically and ecologically important families like Engraulidae, Clupeidae and Mugilidae spawn in the lagoon emphasizes the importance of the Ibiraquera Lagoon as a nursery area for coastal fish. The depletion of coastal stocks is not only being caused by over fishing, but also by destruction of nurseries areas (Palumbi, 2001; Watson & Pauly, 2001; Myers & Worm, 2003; Palumbi, 2003). High numbers of ichthyoplankton were registered during months of increased tourist activity (late spring and summer) in Ibiraquera Lagoon. Therefore, we strongly recommend environmental procedures, as preservation of the lagoon vegetation and maintenance of the water quality, during these periods, especially in the area near the sandbar. Further studies should be conducted to understand the circulation of water throughout the lagoon and to investigate the ichthyoplankton community in the adjacent coastal area.
Acknowledgements
This research was supported by a grant from Fundo Nacional do Meio Ambiente (FNMA). We are grateful to Paulo Freire Vieira NMD (Núcleo Interdisciplinar de Meio Ambiente e Desenvolvimento/UFSC) coordinator of the project "Fisheries Management in the Ibiraquera Lagoon, SC". We would also like to thank Eduardo Sampaio, a local fisherman, for his help during field work, IBAMA/CEPESUL for providing the boat facilities, all the biologists and aquaculture engineers who helped during the sampling trials, Juliana S. Oliveira (UFPR) for initial larval identification and Ana Cristina T. Bonecker (UFRJ) for her help with further larval identification. We are also thankful to Ibiraquera's Agenda 21 Forum and the local traditional leaders.
Literature Cited
Accepted March 2009
Published June 17, 2009
- Avendaño-Ibarra, R., R. Funes-Rodríguez, A. Hinojosa-Medina, R. González-Armas & G. Aceves-Medina. 2004. Seasonal abundance of fish larvae in a subtropical lagoon in the west coast of the Baja California Peninsula. Estuarine, Coastal and Shelf Science, 61(1): 125-135.
- Barletta-Bergan, A., M. Barletta & U. Saint-Paul. 2002a. Community structure and temporal variability of ichthyoplankton in North Brazilian mangrove creeks. Journal of Fish Biology, 61(sA): 33-51.
- Barletta-Bergan, A., M. Barletta & U. Saint-Paul. 2002b. Structure and seasonal dynamics of larval fish in the Caeté River estuary in North Brazil. Estuarine, Coastal and Shelf Science, 54(2): 193-206.
- Barletta, M., A. Barletta-Bergan, U. Saint-Paul & G. Hubold. 2003. Seasonal changes in density, biomass and diversity of estuarine fishes in tidal mangrove creeks of the lower Caeté Estuary (northern Brazilian coast, east Amazon). Marine Ecology Progress Series, 256: 217-228.
- Beck, M. W., K. L. Heck, K. W. Able, D. L. Childers, D. B. Eggleston, B. M. Gillanders, B. Halpern, C. G. Hays, K. Hoshino, T. J. Minello, R. J. Orth, P. F. Sheridan & M. P. Weinstein. 2001. The identification, conservation, and management of estuarine and marine nurseries for fish and invertebrates. BioScience, 51(8): 633-641.
- Berasategui, A. D., E. M. Acha & N. C. F Araoz. 2004. Spatial patterns of ichthyoplankton assemblages in the Río de la Plata Estuary (Argentina-Uruguay). Estuarine, Coastal and Shelf Science, 60(4): 599-610.
- Boglione, C., B. Bertolini, M. Russiello & S. Cataudella. 1992. Embryonic and larval development of the thick-lipped mullet (Chelon labrosus) under controlled reproduction conditions. Aquaculture, 101(3-4): 349-359.
- Bonetti, C., J. Bonetti & E. Beltrame. 2005. Mudanças nas características composicionais das águas da Lagoa de Ibiraquera (SC) em resposta a dinâmica de abertura e fechamento de sua desembocadura. Brazilian Journal of Aquatic Science and Technology, 9(2): 39-47.
- Castro, M. S. & A. C. T. Bonecker. 1996. Ocorrência de larvas de peixe no sistema estuarino do rio Mucuri. Arquivos de Biologia e Tecnologia, 39(1): 171-185.
- Castro, M. S., A. C. T. Bonecker & J. L. Valentin. 2005. Seasonal variation in fish larvae at the entrance of Guanabara Bay, Brazil. Brazilian Archives of Biology and Technology, 48(1): 121-128.
- Chagas, L. P., J. C. Joyeux & F. R. Fonseca. 2006. Small-scale spatial changes in estuarine fish: subtidal assemblages in tropical Brazil. Journal of Marine Biological Association of the United Kingdom, 86: 861-875.
- Ekau, W., P. Westhaus-Ekau & C. Medeiros. 1999. Large scale distribution of fish larvae in the continental shelf waters off North-East Brazil. Archive of Fishery Marine Research, 47(2/3): 183-200.
- Esper, M. L. P., M. S. Menezes & W. Esper. 2001. Época reprodutiva de Mugil platanus (Günther, 1880), Pisces Mugilidae da Baia de Paranaguá (Paraná, Brasil). Acta Biológica Paranaense, 30: 5-17.
- Figueiredo, J. L. & N. A. Menezes. 1985. Manual de peixes marinhos do Sudeste do Brasil, V Teleostei (4). São Paulo, Museu de Zoologia, Universidade de São Paulo, 105p.
- Franco, B. C. & J. H. Muelbert. 2003. Distribuição e composição do ictioplâncton na quebra de plataforma do Sul do Brasil. Atlântica, 25(1): 75-86.
- Freitas, D. M. & J. H. Muelbert. 2004. Ichthyoplankton distribution and abundance off Southeastern and Southern Brazil. Brazilian Archives of Biology and Technology, 47(4): 601-612.
- Garcia, A. M. & J. P. Vieira. 2001. O aumento da diversidade de peixes no estuário da Lagoa dos Patos durante o episódio El Niño 1997-1998. Atlântica, 23: 133-152.
- Ibáñez, A. L. & O. G. Benítez. 2004. Climate variables and spawning migrations of the striped mullet and white mullet in the north-western area of the Gulf of Mexico. Journal of Fish Biology, 65(3): 822-831.
- Ismail, W. A., K. Al-Abdul-Elah & F. Al-Yamani. 1998. Larval development of the back keeled Mullet Liza carinata Hydrobiologia, 385(1-3): 87-105.
- Joyeux, J. C., B. B. Pereira & H. G. Almeida. 2004. The flood-tide ichthyoplanktonic community at the entrance into a Brazilian tropical estuary. Journal of Plankton Research, 26(11): 1277-1287.
- Katsuragawa, M., J. H. Muelbert & J. F. Dias. 2006. O ictioplâncton na região entre o Cabo de São Tomé (RJ) e o Chuí (RS). Pp. 359-446. In: Rossi-Wongschowski, C. L. D. B. & L. S. P. Madureira (Eds.). O Ambiente Oceanográfico da Plataforma Continental e do Talude na Região Sudeste-Sul do Brasil. São Paulo, Edusp, 472p.
- Kjerfve, B. 1994. Coastal lagoon processes. Pp. 1-8. In: Kjerfve, B. (Ed.). Coastal lagoon processes. Amsterdam, Elsevier Oceanography Series, 577p.
- Kuo, C. M., Z. H. Shehadeh & K. K. Milken. 1973. A preliminary report on the development, growth and survival of laboratory reared larvae of the grey mullet, Mugil cephalus L. Journal of Fish Biology, 5(4): 459-470.
- Leis, J. M. & T. Trnski. 1989. The larvae of Indo-Pacific shorefishes. Hawaii and Australia, University of Hawaii Press and New South Wales University Press, 370p.
- Lippson, A. J. & R. L. Moran. 1974. Manual for identification of early development stages of fishes of the Potomac River Estuary. Baltimore, Environmental Technology Center Martin Marietta Corporation, 282p.
- Lüchmann, K. H., A. S. Freire, N. C. Ferreira, F. G. Daura-Jorge & M. R. F. Marques. 2008. Spatial and temporal variations in abundance and biomass of penaeid shrimps in the subtropical Conceição Lagoon, southern Brazil. Journal of the Marine Biological Association of the United Kingdom, 88(2): 293-299.
- Mafalda Jr., P. O., C. Sinque, J. H. Muelbert & C. S. Souza. 2004. Distribuição e abundância do ictioplâncton na costa Norte da Bahia, Brasil. Tropical Oceanography, 32(1): 69-88.
- Magro, M., M. C. Cergole & C. L. D. B. Rossi-Wongstschowski. 2000. Síntese de conhecimentos dos principais recursos pesqueiros costeiros potencialmente explotáveis na costa Sudeste-Sul do Brasil: Peixes. Brasil, Ministério do Meio Ambiente, dos Recursos Hídricos e da Amazônia Legal, 109p.
- Monteiro-Ribas, W. M. & A. C. T. Bonecker. 2001. Artificial fertilization and development in laboratory of Mugil liza (Valenciennes, 1836) (Osteichthyes, Mugilidae). Bulletin of Marine Science, 68(3): 427-433.
- Moser, H. G., W. J. Richards, D. M. Cohen, M. P. Fahay, A. W. Kendall Jr. & S. L. Richardson. 1983. Ontogeny and systematics of fishes. Lawrence, American Society of Ichthyologists and Herpetologists, 760p.
- Muelbert, J. H. & G. Weiss. 1991. Abundance and distribution of fish larvae in the channel area of the Patos Lagoon estuary, Brazil. Pp. 43-54. In: Hoyt, R. D. (Ed.). Larval fish recruitment in the Americas. Proceedings of the thirteenth annual fish conference. Merida, Mexico, NOAA Technical Report NMFS, 95, 137p.
- Myers, R. A. & B. Worm. 2003. Rapid worldwide depletion of predatory fish communities. Nature, 423(6937): 280-283.
- Neira, F. J., A. G. Miskiewicz & T. Trnski. 1998. Larvae of temperate Australian fishes: laboratory guide for larval fish identification. Nedlands, University of Western Australian Press, 474p.
- Palumbi, S. R. 2001. The ecology of marine protected areas. Pp. 509-530. In: Bertness, M. D., S. D. Gaines & M. Hay (Eds.). Marine Community Ecology. Sunderland, Sinauer, 550p.
- Palumbi, S. R. 2003. Populations genetics, demographic connectivity, and the design of marine reserves. Ecological Applications, 13(1): 146-158.
- Peterson, D., N. Hanazaki & P. C. Simões-Lopes. 2008. Natural resource appropriation in cooperative artisanal fishing between fishermen and dolphins (Tursiops truncatus) in Laguna, Brazil. Ocean & Coastal Management, 51: 469-475.
- Phonlor, G. 1984. Morfologia e biologia dos ovos de Engraulidae do sul do Brasil (Teleostei, Clupeiformes). Revista Brasileira de Biologia, 44: 467-487.
- Ramos, S., R. K. Cowen, P. Ré & A. A. Bordalo. 2006. Temporal and spatial distributions of larval fish assemblages in the Lima estuary (Portugal). Estuarine, Coastal and Shelf Science, 66(1-2): 303-314.
- Ré, P. 1999. Ictioplâncton estuarino da Península Ibérica: guia de identificação dos ovos e estados larvares planctônicos. Lisboa, Faculdade de Ciências da Universidade de Lisboa.
- Richards, W. J. 2001. Preliminary Guide to the identification of the Early Life History Stages of Ichthyoplankton of the Western Central Atlantic (Draft Edition). Southeast Fisheries Science Center, NOAA.
- Sánchez-Velasco, L., C. Flores-Coto & B. Shirasago. 1996. Fish larvae abundance and distribution in the coastal zone off Terminos Lagoon, Campeche (Southern Gulf of México). Estuarine, Coastal and Shelf Science, 43(6): 707-721.
- Seixas C. S. & F. Berkes. 2003. Learning from fishers: local knowledge for management design and assessment. Pp. 333-371. In: Vieira, P. F. (Ed.). Conservação da diversidade biológica e cultural em zonas costeiras: enfoques e experiências na América Latina e no Caribe. Florianópolis, APED, 528p.
- Silva, M. A., F. G. Araújo, M. C. C. Azevedo & P. Mendonça. 2003. Distribuição espacial e temporal de Cetengraulis edentulus (Cuvier) (Actinopterygii, Engraulidae) na Baía de Sepetiba, Rio de Janeiro, Brasil. Revista Brasileira de Zoologia, 20(4): 577-581.
- Spach, H. L., C. Santos & R. S. Godefroid. 2003. Padrões temporais na assembléia de peixes na gamboa do Sucuriú, Baía de Paranaguá, Brasil. Revista Brasileira de Zoologia, 20(4): 591-600.
- Tovar-Faro, B. C. M. & A. C. T. Bonecker. 2000. Composição das larvas de peixes no setor mais interno da Baía da Guanabara (RJ Brasil). In: V Simpósio de Ecossistemas Brasileiros: Conservação, v. 2, p. 73-84.
- Vieira, J. P. 1991. Juvenile Mullets (Pisces: Mugilidae) in the Estuary of Lagoa dos Patos, RS, Brazil. Copeia, 1991(2): 409-418.
- Watson, R. & D. Pauly. 2001. Systematic distortions in world fisheries catch trends. Nature, 414(6863): 534-536.
- Yoshimatsu, T., S. Matsui & C. Kitajima. 1992. Early development of laboratory-reared redlip mullet, Liza haematocheila Aquaculture, 105(3-4): 379-390.
- Zar, J. H. 1996. Biostatistical analysis. 3 ed. New Jersey, Prentice Hall, 662p.
Publication Dates
-
Publication in this collection
15 July 2009 -
Date of issue
June 2009
History
-
Accepted
17 June 2009 -
Received
Mar 2009