Abstracts
The structure and composition of a fish community in a tributary stream of the Aquidauana River, located in the plateau region of the Paraguay River basin, central west Brazil are described, with special attention to the differential effects of the phytophysiognomies of the Cerrado (a tropical savanna), which are predominant in riparian zones all along their longitudinal gradient. A total of 1,166 individuals belonging to 44 species were captured. Similarity analyses of composition and abundance showed two groups: upstream reaches (R1 and R2) and downstream reaches (R3, R4 and R5), which presented 85.8% of the average dissimilarity. Although physical habitat and physicochemical descriptors varied along the longitudinal gradient of the Correntes stream, shoreline vegetation was the most important environmental feature predicting fish structure and composition. The Mantel test revealed a correlation between shoreline vegetation structure and fish composition and quantitative structure of the fish community (R > 0.65; p < 0.04). This relationship is driven by the prevalence of species occupying microhabitats associated with shoreline vegetation in contact with water in upstream reaches. Structural differences in shoreline vegetation along the longitudinal gradient correspond to the phytophysiognomic dichotomy observed in the Correntes stream, where riparian vegetation is made up of wet grassland upstream and of gallery forest downstream; this reinforces the importance of the phytophysiognomic heterogeneity of the Cerrado in maintaining ichthyofauna diversity.
Paraguay basin; Cerrado; Longitudinal gradient; River continuum concept; Riparian forest
São descritas a estrutura e composição da comunidade de peixes em um riacho afluente do rio Aquidauana, localizado na região planáltica da bacia do rio Paraguai, centro-oeste do Brasil, com atenção especial aos efeitos diferenciais das fitofisionomias de Cerrado predominantes nas zonas ripárias ao longo do seu gradiente longitudinal. Foram capturados 1.166 indivíduos pertencentes a 44 espécies. As análises de similaridade com a composição e abundância evidenciaram dois grupos: os trechos a montante (R1 e R2) e trechos a jusante (R3, R4 e R5), os quais apresentaram 85,8% de dissimilaridade média. Embora os descritores fisico-químicos e do habitat físico tenham variado ao longo do gradiente longitudinal do córrego Correntes, a vegetação marginal foi o componente ambiental mais importante predizendo a estrutura e composição da ictiofauna. O teste de Mantel revelou correlação entre a estrutura da vegetação marginal com a composição e estrutura quantitativa da comunidade de peixes (R > 0.65; p < 0.04). Essa relação é representada pela prevalência de espécies que exploram micro-habitats associados à vegetação marginal em contato com a água nos trechos a montante. As diferenças na estrutura da vegetação marginal ao longo do gradiente longitudinal correspondem à dicotomia fitofisionômica observada no córrego Correntes, cuja vegetação marginal é constituída de campos úmidos à montante e por fisionomias florestais nos trechos à jusante, reforçando a importância da heterogeneidade fitofisionomica do Cerrado brasileiro para a manutenção da diversidade ictiofaunística.
Influence of the riparian zone phytophysiognomies on the longitudinal distribution of fishes: evidence from a Brazilian savanna stream
Fabrício Barreto Teresa; Renato de Mei Romero
UNESP, Universidade Estadual Paulista, Instituto de Biociências, Letras e Ciências Exatas - IBILCE, Laboratório de Ictiologia, Departamento de Zoologia e Botânica. Rua Cristóvão Colombo, 2265, 15.054-000 São José do Rio Preto, SP, Brazil. fabricioteresa@yahoo.com.br, romerobio@yahoo.com.br
ABSTRACT
The structure and composition of a fish community in a tributary stream of the Aquidauana River, located in the plateau region of the Paraguay River basin, central west Brazil are described, with special attention to the differential effects of the phytophysiognomies of the Cerrado (a tropical savanna), which are predominant in riparian zones all along their longitudinal gradient. A total of 1,166 individuals belonging to 44 species were captured. Similarity analyses of composition and abundance showed two groups: upstream reaches (R1 and R2) and downstream reaches (R3, R4 and R5), which presented 85.8% of the average dissimilarity. Although physical habitat and physicochemical descriptors varied along the longitudinal gradient of the Correntes stream, shoreline vegetation was the most important environmental feature predicting fish structure and composition. The Mantel test revealed a correlation between shoreline vegetation structure and fish composition and quantitative structure of the fish community (R > 0.65; p < 0.04). This relationship is driven by the prevalence of species occupying microhabitats associated with shoreline vegetation in contact with water in upstream reaches. Structural differences in shoreline vegetation along the longitudinal gradient correspond to the phytophysiognomic dichotomy observed in the Correntes stream, where riparian vegetation is made up of wet grassland upstream and of gallery forest downstream; this reinforces the importance of the phytophysiognomic heterogeneity of the Cerrado in maintaining ichthyofauna diversity.
Key words: Paraguay basin, Cerrado, Longitudinal gradient, River continuum concept, Riparian forest.
RESUMO
São descritas a estrutura e composição da comunidade de peixes em um riacho afluente do rio Aquidauana, localizado na região planáltica da bacia do rio Paraguai, centro-oeste do Brasil, com atenção especial aos efeitos diferenciais das fitofisionomias de Cerrado predominantes nas zonas ripárias ao longo do seu gradiente longitudinal. Foram capturados 1.166 indivíduos pertencentes a 44 espécies. As análises de similaridade com a composição e abundância evidenciaram dois grupos: os trechos a montante (R1 e R2) e trechos a jusante (R3, R4 e R5), os quais apresentaram 85,8% de dissimilaridade média. Embora os descritores fisico-químicos e do habitat físico tenham variado ao longo do gradiente longitudinal do córrego Correntes, a vegetação marginal foi o componente ambiental mais importante predizendo a estrutura e composição da ictiofauna. O teste de Mantel revelou correlação entre a estrutura da vegetação marginal com a composição e estrutura quantitativa da comunidade de peixes (R > 0.65; p < 0.04). Essa relação é representada pela prevalência de espécies que exploram micro-habitats associados à vegetação marginal em contato com a água nos trechos a montante. As diferenças na estrutura da vegetação marginal ao longo do gradiente longitudinal correspondem à dicotomia fitofisionômica observada no córrego Correntes, cuja vegetação marginal é constituída de campos úmidos à montante e por fisionomias florestais nos trechos à jusante, reforçando a importância da heterogeneidade fitofisionomica do Cerrado brasileiro para a manutenção da diversidade ictiofaunística.
Introduction
The upper Paraguay River basin is one of the most important basins in the Neotropics, occupying an area of approximately 363,442 km² in Brazilian territory. It can be divided into two distinct regions: a floodplain called the Pantanal which is one of largest wetland areas in the world and a plateau around the Pantanal, including areas with an altitude of over 200 m. These two regions are distinguished by geologic and geomorphologic characteristics, but they are interdependent in ecological terms since headwaters of the most important rivers that supply the Pantanal are situated in the plateau region, providing high hydrologic connectivity (Willink et al., 2000).
The plateau region is biologically distinct from the floodplain (Chernoff & Willink, 2000), and its ichthyofauna, as well as the factors that modulate it, is poorly known when compared to the accumulated scientific knowledge about the Pantanal region (Catella et al., 1996; Súarez et al., 2004; Britski et al., 2007). Knowledge is especially scarce for small fishes from streams, which make up the greatest proportion of fish species richness of the Neotropics (Castro, 1999).
Most of the Paraguay basin plateau streams flow into the Brazilian Cerrado (savanna) biome, which is a world biodiversity hotspot due to its richness, high degree of endemism and anthropogenic threat (Ratter et al., 1997; Myers et al., 2001). Vegetation in these areas is very heterogeneous, and riparian vegetation is one example of this variation, showing phytophysiognomies ranging from grassland to closed canopy (Ratter et al., 1997; Ribeiro & Walter, 1998). Considering the strong predictive power of riparian vegetation in structuring stream fish communities (Wichert & Rapport, 1998; Growns et al., 2003), it is plausible that the spatial distribution of fish species can be affected by alterations in the longitudinal variation of riparian vegetation structure in Cerrado streams.
The purpose of this study was to describe fish community structure and composition in a stream located in the plateau region of the Paraguay basin, central west Brazil, with special attention to the differential effects of the predominant Cerrado phytophysiognomy in riparian zones. In this stream, shoreline vegetation of the upstream reaches is formed by wet grassland, a phytophysiognomy characterized by the dominance of herbaceous species, while in downstream reaches gallery forest predominates.
Material and Methods
This study was carried out in the Correntes stream, a tributary on the left bank of the Aquidauana River (Fig. 1), located in Dois Irmãos do Buriti municipality, Mato Grosso do Sul State, central west Brazil, Paraguay basin. This stream is roughly 18.5 km in length and its catchment encompasses approximately 11,690 ha, where cattle pastures and eucalyptus plantations predominate. The region is located in the São Bento formation, represented by the Botucatu group. As to geomorphology, it belongs to the Maracaju plateau, a region characterized by plains topography and wide hills. The climate is wet mesothermal with a rainy summer (October to March) and a dry winter (April to September); annual precipitation ranges from 1,300 to 1,400 mm.
Five 100-m reaches were selected (R1-R5) along the longitudinal gradient of the Correntes stream (Table 1). The distance between each reach was 2.5 km, except between R4 and R5, where the distance was 5.2 km (Fig. 1). One sampling of the ichthyofauna was conducted in both the wet (February 2008) and dry seasons (September 2008). Fish were collected by use of a hand seine (2.5 m in width x 1.5 m in height, 3 mm mesh) and a dip net (0.9 m in width x 0.45 m in height, 3 mm mesh) during a period of 60 min. Fish were fixed in 10% formalin and afterward transferred to 70% EtOH. Voucher specimens were deposited in the fish collection at the Departamento de Zoologia e Botânica da Universidade Estadual Paulista (DZSJRP), São José do Rio Preto, São Paulo State, Brazil.
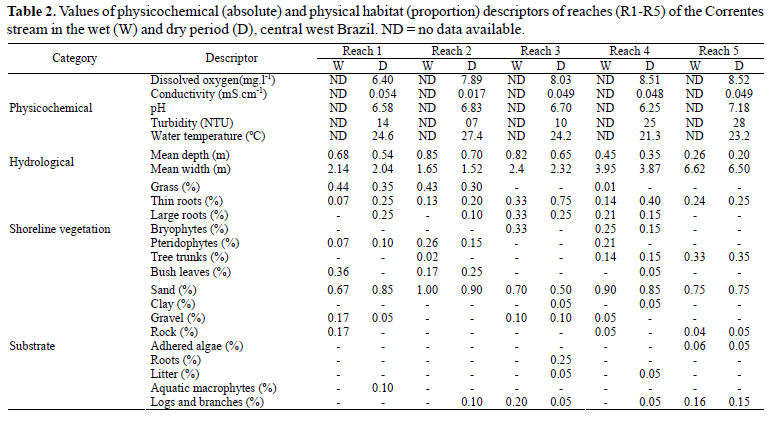
Eighteen habitat structure descriptors related to in-stream and shoreline physical structures were estimated visually along each reach in the wet and dry periods. These descriptors were divided into three categories related to shoreline vegetation structure, substrate composition and hydrological measurements. Descriptors related to shoreline vegetation comprise characteristics of shoreline vegetation in contact with the water as follows: proportion of shoreline occupied by grass, thin roots, large roots, bryophytes, pteridophytes, tree trunks, and bush leaves. Substrate composition was evaluated by estimating the proportion of bottom occupied by clay, sand, gravel, rock, algae, roots, litter, aquatic macrophytes and logs and branches. Hydrological descriptors included depth and width measurements obtained by means of transect measured along each reach. Additionally, a physicochemical analysis including measurement of dissolved oxygen, conductivity, pH, turbidity and temperature (all measured using electronic equipment, Horiba®, model U-10) was carried out. These analyses were only performed in the dry period due to operational problems with the equipment in the wet period.
Species diversity was calculated using the Shannon-Wiener index (H') as described by Magurran (2004), and species dominance was obtained using the Berger-Parker dominance index (where abundance of the dominant species is divided by the total abundance of the species, multiplied by 100). The spatial-temporal ordination of the reaches was evaluated by non-metric multidimensional scaling (NMDS) analysis, considering ichthyofauna composition and the quantitative structure of the community. This analysis resulted in stress values of 0.08 and 0.05, respectively, indicating good ordination and reliability of resulting interpretation (Clarke & Warwick, 2001). To complement the ordination analysis, a two-way analysis of similarity (routine ANOSIM) was used to verify differences in the fish composition and quantitative structure of communities between rainy and dry seasons and between groupings determined by ordination analysis (NMDS). In order to investigate if spatial-temporal variation in the environmental descriptors had a pattern similar to fish abundance data, the same analysis was performed for shoreline vegetation structure, substrate composition and hydrological measurements. Additionally, a similarity percentage analysis (routine SIMPER) was applied to determine which species mainly explained differences in community composition between groupings established by NMDS.
The relative importance of environmental descriptors (shoreline vegetation, substrate and hydrological measurements) structuring ichthyofauna in the Correntes stream was evaluated by 2Stage analysis. This analysis indicates which categories of the environmental descriptors are more correlated to biotic similarity matrices, calculating the rank correlation between all pairs of matrices. Afterward, the significance of the correlation between biotic and abiotic matrices was tested using the Mantel test in the Past software, version 1.68 (Hammer et al., 2001). In this case, data were pooled from the wet and dry seasons by summing species abundance (biotic data) and by means (abiotic data), except for physicochemical data which were obtained just in the dry period. In this case, abiotic data was correlated with biotic ones of the same period. Data were transformed according to the arcsin procedure for proportions (shoreline and substrate descriptors), log (x+1) procedure for hydrological measurements (Zar, 1999) and fourth root for abundance matrix. Similarity matrices were constructed using the Euclidean distance similarity measure for abiotic data (Clarke & Ainsworth, 1993), Jaccard similarity coefficient for ichthyofauna composition and Bray-Curtis similarity coefficient for quantitative structure of the communities. All analyses (except the Mantel test) were performed using Primer 6 (2006) software.
Results
Spatial variability on the environmental descriptors was greater than temporal variability (Table 2). Reaches 1, 2 and 3 were narrower and deeper than downstream reaches (Table 2). Grass and bush leaves were the most representative shoreline vegetation in contact with water in reaches 1 and 2, whereas in the downstream reaches, tree trunks and roots were the vegetation most representative (Table 2). Substrate was uniform along the longitudinal gradient of Correntes stream with a predominance of sand in all reaches (Table 2). A total of 1,166 individuals were sampled, belonging to six orders, 18 families and 44 species (Table 3). Characiformes and Siluriformes represented 85.3% of the species recorded and approximately 97.2% of total abundance. The most abundant species were Odontostilbe pequira (24.3%), Jupiaba acanthogaster (14.0%), Bryconamericus exodon (9.0%), Characidium aff. zebra (7.9%), Moenkhausia bonita (6.4%), Hemiloricaria lanceolata (4.6%), Hypostomus sp. (4.6%) and Astyanax abramis (4.6%) (Table 3). Five species (11.4%) were recorded in all reaches in at least one sampling: Astyanax abramis, Bryconamericus exodon, Jupiaba acanthogaster, Hypostomus sp. and Hemiloricaria lanceolata.
Total abundance obtained in the rainy season was greater than in the dry, except for reach 2 (Table 3). Reach 5 exhibited the greatest species richness, followed by reaches 1 and 4 which had greater species diversity but less dominance. Reach 2 had low species richness and diversity and high Jupiaba acanthogaster dominance. Reach 3 had the lowest species richness of all and low dominance. Although having the greatest species richness, reach 5 had low diversity and higher dominance due to the high relative abundance of Odontostilbe pequira.
The ordination generated by NMDS, considering the fish composition and quantitative structure of the communities, revealed two subsets formed by upstream reaches (sites 1 and 2) and downstream reaches (sites 3, 4 and 5), notwithstanding seasonality (Fig. 2). This was confirmed by two-way ANOSIM which indicated significant differences of ichthyofauna composition and quantitative structure between upstream and downstream reaches (R = 1.00; p = 0.01 for both), but not between rainy and dry seasons (p > 0.40).
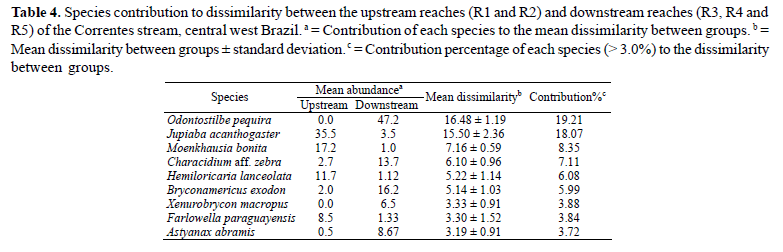
The SIMPER procedure revealed that the two sub-sets indicated in the NMDS, i.e., the upstream reaches (R1 and R2) and the downstream reaches (R3, R4 and R5), had an average dissimilarity of 85.8%. This pattern was principally obtained because of the differential contribution of Jupiaba acanthogaster, Odontostilbe pequira, Moenkhausia bonita, Characidium aff. zebra, Hemiloricaria lanceolata and Bryconamericus exodon along the longitudinal gradient of the Correntes stream (Table 4).
In a similar way, the shoreline vegetation structure differed significantly between upstream and downstream reaches (R = 0.79; p = 0.01), but not between wet and dry periods (R = 0.06; p = 0.37). Substrate composition and hydrological descriptors did not differ significantly between upstream and downstream reaches (p > 0.46) not between seasons (p > 0.30). Shoreline vegetation structure was more related to fish composition and quantitative structure ofcommunity (2Stage analysis: R = 0.63 and 0.62, respectively)than substrate composition and hydrological measurements(0.28 > R > -0.32). This was confirmed by Mantel tests whichrevealed a significant correlation of fish composition andquantitative structure of the fish communities with theshoreline vegetation (R > 0.65; p < 0.04), but not with substratecomposition, width and depth and physicochemicalmeasurements (p > 0.12). Qualitatively, it was shown that Gymnotiformes species such as Apteronotus albifrons,Brachyhypopomus sp., Eigenmannia trilineata andGymnotus sp. and the loricariids Farlowella paraguayensis,Hemiloricaria lanceolata and Rineloricaria cacerensis werealways captured associated with thin roots, shrub leaves andgrass in contact with the water.
Discussion
The two most abundant species, Odontostilbe pequiraand Jupiaba acanthogaster, were also the ones that mostcontributed to the dissimilarity between up- and downstreamreaches in the Correntes stream. Jupiaba acanthogaster wasmore restricted to upstream reaches, while Odontostilbepequira was exclusive to downstream reaches and waspredominant in R5. Marginal areas with moderate water flowseem to favor O. pequira in this reach. The occurrence ofJupiaba acanthogaster in upstream reaches of the Correntesstream is compatible with findings of their preferentialoccurrence in headwater streams of the Serra da Bodoquena(L. Casatti & R. M. Romero, pers. comm.).
The greater species abundance recorded during the rainyseason can be explained by the increased volume of the habitat, which provides a higher availability of microhabitats and increases the amount of allochthonous material in the stream, increasing the availability of food and substrates that can be used as shelter and as reproductive sites (Schlosser, 1982; Angermeier & Karr, 1983; Casatti, 2005). Moreover, the rainy season is also the reproduction period for many tropical species when there is an increase in abundance, especially of young in the streams (Casatti, 2005; Súarez et al., 2007). Reach 2 with its lower abundance in the rainy season may be due to a sampling artifact, since there are limitations to the collecting methodology used in deeper localities, as is the case with this reach in rainy periods. Despite potential limitation of the sampling techniques used in this study (Wiley & Tsai, 1983; Mercado-Silva & Escandón-Sandoval, 2008), there was no evidence of differential catchability between reaches along
the longitudinal gradient of the Correntes stream, except between dry and wet period in reach 2. In this case, abundance rather than richness seems to have been underestimated, but this did not affect the results because relations obtained from fish composition and quantitative structure of communities were congruent in all analyses.
Despite the tendency for an increased abundance during the rainy season, similarity analyses results indicate that such differences were not enough to alter the structure of the communities. Studying the fish communities in headwater streams of Paraguay and Paraná basins, Valério et al. (2007) found little seasonal influence on the organization of communities, a pattern comparable to that seen in this study. The increase in richness and diversity along the longitudinal gradient predicted by the River Continuum Concept (Vannote et al., 1980) has been confirmed in rivers and streams in different zoogeographic regions (Sheldon, 1968; Evans & Noble, 1979; Garutti, 1988; Oberdorff et al., 1993; Casatti, 2005; Petry & Schulz, 2006). In the Correntes stream, this pattern was especially evident from the intermediate reach (R3) to the extreme downstream reach (R5) (14 to 25 species), apparently following the increase in habitat volume (Angermeier & Schlosser, 1989; Casatti, 2005). Moreover, fish migration from bigger rivers to tributaries is also a factor that contributes toward an increase in species richness in reaches of the stream near the mainstem (Gorman, 1986; Fernandes et al., 2004). Therefore, the proximity of R5 to the Aquidauana River (less than 600 m) could also have contributed to the maintenance of richer communities in the downstream reaches.
Surprisingly, headwater reaches, normally with smaller species richness (Whiteside & McNatt, 1972; Garutti, 1988; Casatti, 2005), had high species richness and diversity in the Correntes stream. This happens when the headwaters have singular characteristics such as a high structural heterogeneity, making the occurrence of richer communities feasible (McNeely, 1986; Ferreira & Casatti, 2006). In fact, upstream reaches of the Correntes stream maintain high structural diversity, especially the abundance of shoreline vegetation in contact with the water; notably grasses, bushes and roots. The high structural diversity of the banks was a marked characteristic of upstream reaches, allowing the occurrence of diverse species, notably fishes of the orders Gymnotiformes and Siluriformes, which were abundant at these sites. These results reinforce the predictive role of the structural composition of the habitat for the structuring of fish communities (Gorman & Karr, 1978; Schneider & Winemiller, 2008).
The addition of species was the most important process in the downstream reaches (R3 to R5), which is generally the result of increased habitat volume and complexity along the longitudinal gradient of the streams, providing favorable conditions for the occurrence of a greater number of species (Sheldon, 1968; Angermeier & Schlosser, 1989). Nevertheless, the high species richness in upstream reaches, among which 12 were exclusive, and its high dissimilarity in relation to downstream reaches, evidenced by NMDS and SIMPER, also suggests species substitution as an important process in the structuring of fish communities in the Correntes stream. Species substitution is predominant when environmental changes along the longitudinal gradient are sudden, causing physical or biological impediments to ichthyofauna longitudinal colonization (Burton & Odum, 1945; Tramer & Rogers, 1973). In fact, the structural composition of habitats in upstream areas is quite distinct from that of downstream reaches, apparently limiting the occurrence of some species while favoring others.
Although physical habitat and physicochemical descriptors varied along the longitudinal gradient of the Correntes stream, the shoreline vegetation was the most important environmental feature predicting fish structure and composition as evidenced by 2Stage analysis and Mantel tests. Structural differences between upstream and downstream reaches correspond to the phytophysiognomy dichotomy observed in the Correntes stream whose shoreline vegetation consists of wet grassland upstream and gallery forest downstream. Wet grasslands are characterized by the dominance of herbaceous and shrubby vegetation adapted to hydromorphic soil (Ribeiro & Water, 1998). In upstream reaches, the small size of the abundantly present shoreline vegetation on the low banks favors their contact with the stream water. Foliage leaning over the channel and roots extending internally increase the spatial heterogeneity of in-stream habitats, creating specific microhabitats (Collier et al., 1999; Rocha et al., 2009) usually explored by species with specialized needs. In fact, some species such as Apteronotus albifrons, Brachyhypopomus sp., Eigenmannia trilineata and Gymnotus sp. were captured exclusively in upstream reaches (Table 3) and always associated with shoreline vegetation in contact with the water. This is probably associated with preferential use of submerged vegetation by Gymnotiformes species for diurnal shelter and feeding (Ferreira & Casatti, 2006; Ferreira, 2007; Rocha et al., 2009). Two other species, Farlowella paraguayensis and Hemiloricaria lanceolata, also occurred associated with marginal areas with dense vegetation and were important for the dissimilarity between upstream and downstream reaches according to the similarity percentage analysis. These data are in agreement with the Mantel test result which indicated an association between shoreline vegetation in contact with water and the composition and quantitative structure of the fish community. On the other hand, most of the species associated with downstream areas that contributed toward dissimilarity between upstream and downstream reaches were water column swimmers such as Bryconamericus exodon and Astyanax abramis. Their preference for downstream reaches was probably related to the increased habitat volume.
The Correntes stream ichthyofauna is structured into two biologically distinct zones, with upstream reaches having particular structural and biological conditions. This reinforces the importance of preserving headwaters to maintain regional biodiversity (Paller, 1994). The composition of ichthyocenosis and structure differences along the longitudinal gradient are influenced by differential contributions of phytophysiognomies in determining specific structural patterns on stream banks. This provides support for the generalization that phytophysiognomic diversity is fundamental for the maintenance of biodiversity in the Brazilian Cerrado (Mares et al., 1986; Ratter et al., 1997; Oliveira & Marquis, 2002; Almeida & Louzada, 2009).
Acknowledgements
This study is part of the Reflorestar Legal project, a partnership comprising MMX Metálicos Corumbá Ltda and Fundação Neotrópica do Brasil. The authors thank the following: MMX Metálicos Corumbá Ltda. - Unidade Florestal and the Fundação Neotrópica do Brasil for financial and logistic support; Vivian R. B. Maria for the in loco floral analysis; Francisco Langeani for helping with fish identification; José L.Veronezzi-Junior for helping with the fish collecting; Lilian Casatti and Fernando R. Carvalho for critical reading of and suggestions to the early version of the manuscript; and two anonymous reviewers for helpful suggestions and comments that improved the manuscript. FBT and RMR are recipients of a doctoral fellowship from Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP: 2008/03583-9, 2007/05400-6, respectively).
Literature Cited
Accepted December 22, 2009
Published March 31, 2010
- Almeida, S. S. P. & J. N. C. Louzada. 2009. Estrutura da comunidade de Scarabaeinae (Scarabaeidae: Coleoptera) em fitofisionomias do Cerrado e sua importância para a conservação. Neotropical Entomology, 38(1): 32-43.
- Angermeier, P. L. & J. R. Karr. 1983. Fish communities along environmental gradients in a system of tropical streams. Environmental Biology of Fishes, 9: 117-135.
- Angermeier, P. L. & I. J. Schlosser. 1989. Species-area relationship for stream fishes. Ecology, 70(5): 1450-1462.
- Britski, H. A., K. Z. S. Silimon & B. S. Lopes. 2007. Peixes do Pantanal: manual de identificação. Brasília, Embrapa, 184p.
- Burton, G. W. & E. P. Odum. 1945. The distribution of stream fish in the vicinity of Mountain Lake, Virginia. Ecology, 26(2): 182-194.
- Casatti, L. 2005. Fish assemblage structure in a first order stream, southeastern Brazil: longitudinal distribution, seasonality, and microhabitat diversity. Biota Neotropica, 5(1): 1-9.
- Castro, R. M. C. 1999. Evolução da ictiofauna de riachos sul-americanos: padrões gerais e possíveis processos causais. Oecologia Brasiliensis, 6: 139-155.
- Catella, A. C. & M. Petrere-Jr. 1996. Feeding patterns in a fish community of Baía da Onça, a floodplain lake of the Aquidauana river, Pantanal, Brazil. Fisheries Management and Ecology, 3: 229-237.
- Chernoff, B. & P. W. Willink. 2000. Ecological and geographical structure in assemblages of freshwater fishes in the Pantanal, Mato Grosso do Sul, Brasil. Pp. 183-201. In: Willink, P. W., B. Chernoff, L. E. Alonso, J. R. Montambault & R. Lourival (Eds.). A Biological Assessment of the Aquatic Ecosystems of the Pantanal, Mato Grosso do Sul, Brazil. Washington, Bulletin of Biological Assessment 18, Conservation International, 307p.
- Clarke, K. R. & R. M. Warwick. 2001. Change in marine communities: an approach to statistical analysis and interpretation. Plymouth, Plymouth Marine Laboratory, 144p.
- Collier, K. J., P. D. Champion & G. F. Crocker. 1999. Patch-and reach-scale dynamics of a macrophyte-invertebrate system in a New Zealand lowland stream. Hydrobiologia, 392: 89-97.
- Evans, J. W. & R. L. Noble. 1979. The longitudinal distribution of fishes in an east Texas stream. American Midland Naturalist, 101(2): 333-343.
- Fernandes, C. C., J. Podos & J. G. Lundberg. 2004. Amazonian ecology: tributaries enhance the diversity of electric fishes. Science, 305: 1960-1962.
- Ferreira, K. M. 2007. Biology and ecomorphology of stream fishes from the rio Mogi-Guaçu basin, Southeastern Brazil. Neotropical Ichthyology, 5(3): 311-326.
- Ferreira, C. P. & L. Casatti. 2006. Influência da estrutura do hábitat sobre a ictiofauna de um riacho em uma micro-bacia de pastagem, São Paulo, Brasil. Revista Brasileira de Zoologia, 23(3): 642-651.
- Garutti, V. 1988. Distribuição longitudinal da ictiofauna de um Córrego na região noroeste do Estado de São Paulo, Bacia do rio Paraná. Revista Brasileira de Biologia, 48: 747-759.
- Gorman, O. T. 1986. Assemblage organization of stream fishes: the effect of rivers on adventitious streams. The American Naturalist, 128(4): 611-616.
- Gorman, O. T. & J. R. Karr. 1978. Habitat structure and stream fish communities. Ecology, 59: 507-515.
- Growns, I., P. C. Gehrke, K. L. Astlea & D. A. Pollard. 2003. A comparison of fish assemblages associated with different riparian vegetation types in the Hawkesbury-Nepean River system. Fisheries Management and Ecology, 10: 209-220.
- Hammer, Ø., D. A. T. Harper & P. D. Ryan. 2001. PAST: paleontological statistics software package for education and data analysis. Palaeontologia Electronica, 4(1): 1-9.
- Magurran, A. E. 2004. Measuring Biological Diversity. United Kingdom, Blackwell Publishing, 256p.
- Mares, M. A., K. A. Ernest & D. D. Gettinger. 1986. Small mammal community structure and composition in the Cerrado Province of central Brazil. Journal of Tropical Ecology, 2: 289-300.
- McNeely, D. L. 1986. Longitudinal patterns in the fish assemblages of an Ozark stream. Southwestern Naturalist, 31: 375-380.
- Mercado-Silva, N. & D. S. Escandón-Sandoval. 2008. A comparison of seining and electrofishing for fish community bioassessment in a Mexican Atlantic slope montane river. North American Journal of Fisheries Management, 28: 1725-1732.
- Myers, N., R. Mittermeier, C. Mittermeier, G. Fonseca & J. Kent. 2000. Biodiversity hotspots for conservation priorities. Nature, 403(6772): 853-858.
- Oberdorff, T., E. Guilbert & J. C. Lucchetta. 1993. Patterns of fish species richness in the Seine River basin, France. Hydrobiologia, 259: 157-167.
- Oliveira, P. S. & R. J. Marquis. 2002. The cerrados of Brazil: ecology and natural history of a Neotropical savanna. Irvington, Columbia University Press, 398p.
- Paller, M. H. 1994. Relationships between fish assemblage structure and stream order in South Carolina coastal plain streams. Transactions of the American Fisheries Society, 123: 150-161.
- Petry, A. C. & U. H. Schulz. 2006. Longitudinal changes and indicator species of the fish fauna in the subtropical Sinos river, Brazil. Journal of Fish Biology, 69: 272-290.
- Ratter, J. A., J. F. Ribeiro & S. Bridgewater. 1997. The Brazilian cerrado vegetation and threats to its biodiversity. Annals of Botany, 80: 223-230.
- Ribeiro, J. F. & B. M. T. Walter. 1998. Fitofisionomias do Bioma Cerrado. Pp. 89-166. In: Sano, S. M. & S. P. Almeida (Eds.). Cerrado: ambiente e flora. Planaltina, Embrapa/CPAC, 556p.
- Rocha, F. C., L. Casatti, F. R. Carvalho & A. M. Silva. 2009. Fish assemblages in stream stretches occupied by cattail (Typhaceae, Angiospermae) stands in Southeast Brazil. Neotropical Ichthyology, 7(2): 241-250.
- Schlosser, I. J. 1982. Fish Community structure and function along two habitat gradients in a headwater stream. Ecological Monographs, 52(4): 395-414.
- Schneider, K. N. & K. O. Winemiller. 2008. Structural complexity of woody debris patches influences fish and macroinvertebrate species richness in a temperate floodplain-river system. Hydrobiologia, 610: 235-244.
- Sheldon, A. L. 1968. Species diversity and longitudinal succession in stream fishes. Ecology, 49(2): 194-198.
- Súarez, Y. R., M. Petrere-Jr. & A. C. Catella. 2004. Factors regulating diversity and abundance of fish communities in Pantanal lagoons, Brazil. Fisheries Management and Ecology, 11: 45-50.
- Súarez, Y. R., S. B. Valério, K. K. Tondado, A. C. Florentino, T. R. A. Felipe, L. Q. L. Ximenes & L. S. Lourenço. 2007. Fish species diversity in headwaters streams of Paraguay and Paraná basins. Brazilian Archives of Biology and Technology, 50(6): 1033-1042.
- Tramer, E. J. & P. M. Rogers. 1973. Diversity and longitudinal zonation in fish populations of two streams entering a metropolitan area. American Midland Naturalist, 90(2): 366-374.
- Valério, S. B., Y. R. Súarez, T. R. A. Felipe, K. K. Tondato & L. Q. L. Ximenes. 2007. Organization patterns of headwater-stream fish communities in the Upper Paraguay-Paraná basins. Hydrobiologia, 583:241-250.
- Vannote, R. L.,G. W. Minshall, K. W. Cummins, J. R. Sedel & C. E. Cushing. 1980. The river continuum concept. Canadian Journal of Fisheries and Aquatic Science, 37: 130-137.
- Whiteside, B. G. & R. M. McNatt. 1972. Fish species diversity in relation to stream order and physicochemical conditions in the Plum creek drainage basin. American Midland Naturalist, 88(1): 90-101.
- Wichert, G. A. & D. J. Rapport. 1998. Fish community structure as a measure of degradation and rehabilitation of riparian systems in an agricultural drainage basin. Environmental Management, 22(3): 425-443.
- Wiley, M. L. & C. F. Tsai. 1983. The relative efficiencies of electrofishing vs seines in Piedmont streams of Maryland. North American Journal of Fisheries Management, 3: 243-253.
- Willink, P. W., O. Froehlich, A. Machado-Alisson, N. Menezes, O. Oyakawa, A. Catella, B. Chernoff, F. C. T. Lima, M. Toledo-Piza, H. Hortega, A. M. Zanata & R. Barriga. 2000. Fishes of the Rio Negro, Negrinho, Taboco, Aquidauana, Taquari and Miranda, Pantanal, Brasil: diversity, distribution, critical habitats and value. Pp. 63-81. In: Willink, P. W., B. Chernoff, L. E. Alonso, J. R. Montambault & R. Lourival (Eds.). A Biological Assessment of the Aquatic Ecosystems of the Pantanal, Mato Grosso do Sul, Brazil. Washington, Bulletin of Biological Assessment 18, Conservation International, 307p.
Publication Dates
-
Publication in this collection
19 Apr 2010 -
Date of issue
Mar 2010
History
-
Received
22 Dec 2009 -
Accepted
31 Mar 2010