Abstracts
A new species of Moenkhausia from the upper rio Xingu basin is described. Moenkhausia eurystaenia is distinguished from all congeners, except M. heikoi and M. phaeonota, by the presence of a dark, broad longitudinal stripe from the opercle to the end of caudal peduncle. The new species is distinguished from M. heikoi mainly by having 18-21 branched anal-fin rays (vs. 24-26) and the absence of a round blotch at the end of the caudal peduncle (vs. present). Moenkhausia eurystaenia can be distinguished from M. phaeonota mainly by the anterior portion of longitudinal band darker at horizontal septum (vs. darker below horizontal septum), the deep dark pigmentation along horizontal septum extending anteriorly up to humeral region (vs. extending up to vertical through pelvic-fin origin), number of vertebrae 34 (vs. 36-37), and others additional measurements.
Systematics; Tetra; Amazon basin; Taxonomy
Uma espécie nova de Moenkhausia da bacia do alto rio Xingu é descrita. Moenkhausia eurystaenia distingue-se de todas as congêneres, exceto M. heikoi e M. phaeonota, pela presença de uma faixa longitudinal larga e escura que se estende do opérculo ao pedúnculo caudal. A espécie nova distingue-se de Moenkhausia heikoi principalmente por apresentar 18-21 raios ramificados na nadadeira anal (vs. 24-26), e ausência de uma mácula circular no final do pedúnculo caudal (vs. presença). Moenkhausia eurystaenia distingue-se de M. phaeonota principalmente pela porção anterior da faixa longitudinal ser mais escura no septo horizontal (vs. mais escura abaixo do septo horizontal), pigmentação profunda e escura ao longo do septo horizontal se estendendo até a região umeral (vs. se estendendo até a vertical que passa pela origem da nadadeira pélvica), 34 vértebras (vs. 36-37), e outras medidas adicionais.
A new species of Moenkhausia Eigenmann (Characiformes: Characidae) from the rio Xingu basin, Brazil
Manoela M. F. Marinho
Museu de Zoologia da Universidade de São Paulo, Cx. Postal 42494, 04299-970 São Paulo, SP, Brazil. manumfm@yahoo.com.br
ABSTRACT
A new species of Moenkhausia from the upper rio Xingu basin is described. Moenkhausia eurystaenia is distinguished from all congeners, except M. heikoi and M. phaeonota, by the presence of a dark, broad longitudinal stripe from the opercle to the end of caudal peduncle. The new species is distinguished from M. heikoi mainly by having 18-21 branched anal-fin rays (vs. 24-26) and the absence of a round blotch at the end of the caudal peduncle (vs. present). Moenkhausia eurystaenia can be distinguished from M. phaeonota mainly by the anterior portion of longitudinal band darker at horizontal septum (vs. darker below horizontal septum), the deep dark pigmentation along horizontal septum extending anteriorly up to humeral region (vs. extending up to vertical through pelvic-fin origin), number of vertebrae 34 (vs. 36-37), and others additional measurements.
Key words: Systematics, Tetra, Amazon basin, Taxonomy.
RESUMO
Uma espécie nova de Moenkhausia da bacia do alto rio Xingu é descrita. Moenkhausia eurystaenia distingue-se de todas as congêneres, exceto M. heikoi e M. phaeonota, pela presença de uma faixa longitudinal larga e escura que se estende do opérculo ao pedúnculo caudal. A espécie nova distingue-se de Moenkhausia heikoi principalmente por apresentar 18-21 raios ramificados na nadadeira anal (vs. 24-26), e ausência de uma mácula circular no final do pedúnculo caudal (vs. presença). Moenkhausia eurystaenia distingue-se de M. phaeonota principalmente pela porção anterior da faixa longitudinal ser mais escura no septo horizontal (vs. mais escura abaixo do septo horizontal), pigmentação profunda e escura ao longo do septo horizontal se estendendo até a região umeral (vs. se estendendo até a vertical que passa pela origem da nadadeira pélvica), 34 vértebras (vs. 36-37), e outras medidas adicionais.
Introduction
Moenkhausia Eigenmann, 1903 was redefined by Eigenmann (1917) to include species with a combination of premaxillary teeth in two rows, the inner with five teeth, complete lateral line, and partially scaled caudal fin. At the present time, Moenkhausia is represented by 70 valid species occurring in South American drainages (Lima et al., 2003; Eschmeyer, 2010).
The new Moenkhausia species described below was collected sympatrically with a similar congener, M. phaeonota Fink, in a clear water stream in the upper rio Xingu basin, Brazil. Both species share a dark broad and straight longitudinal band on body, extending to middle caudal-fin rays, with a pale longitudinal line immediately above, and concentration of dark chromatophores behind the opercle. Similar color pattern is found in other small characids such as Moenkhausia heikoi Géry & Zarske, Tucanoichthys tucano Géry & Römer, and some Hyphessobrycon Durbin in Eigenmann species.
Material and Methods
Counts and measurements were taken with digital calipers under a stereo microscope, according to Fink & Weitzman (1974), with the addition of head depth, measured at vertical through the posteriormost tip of bony opercle. Horizontal scale rows below lateral line were counted to the pelvic-fin insertion. Counts are followed by their occurrence in parentheses; asterisks indicate the count of the holotype. Measurements are given as percents of standard length (SL), except subunits of the head, which are given as percents of head length (HL). Supraneurals, gill-rakers of first arch, tooth cusps, unbranched anal-fin rays, procurrent caudal-fin ray counts and position of pterygiophores were taken from cleared and stained (c&s) specimens, prepared following Taylor & van Dyke (1985). Vertebrae count was taken from c&s and radiographs from alcohol preserved specimens. Vertebrae of the Weberian apparatus were counted as four elements and the fused PU1+U1 of the caudal region as a single element.
Jaws for Scanning Electron Micrographs (SEM) images were removed from c&s specimens, immersed in weak (less than 1%) sodium hypochlorite solution and dried by a successively more concentrated solutions of alcohol, and then acetone. Catalog numbers are followed by the number of specimens in alcohol and the SL range, the number and SL range of measured specimens in parenthesis, when different, and if any, the number of c&s specimens and their SL range. Measured specimens of Moenkhausia phaeonota comes from the lots MZUSP, 13793, 13794, 13795, 45301, 61043, and 91403. Institutional abbreviations used are: CPUFMT, Coleção de Peixes da Universidade Federal do Mato Grosso; DZSJRP, Departamento de Zoologia e Botânica da Universidade Estadual Paulista, São José do Rio Preto; INPA, Instituto Nacional de Pesquisas da Amazônia, Manaus; MCP, Museu de Ciências e Tecnologia, Pontifícia Universidade Católica do Rio Grande do Sul, Porto Alegre; MPEG, Museu Paraense Emílio Goeldi, Belém; MZUSP, Museu de Zoologia da Universidade de São Paulo, São Paulo; and USNM, National Museum of Natural History, Smithsonian Institution, Washington, DC.
Moenkhausia eurystaenia, new species
Holotype. MZUSP 106058, 36.8 mm SL, rio Von Den Stein, on the road to Água Limpa, 13°14'19"S 54°53'01"W, rio Ronuro drainage, rio Xingu basin, Nova Ubiratã, Mato Grosso, Brazil, 24 Oct 2004, J. L. Birindelli, O. T. Oyakawa, C. R. Moreira, M. I. Landim, A. K. Oliveira, A. Datovo & J. C. Nolasco.
Paratypes. CPUFMT 610, 3, 25.2-28.6 mm SL; DZSJRP 12547, 3, 24.4-24.6 mm SL; INPA 34095, 3, 23.3-26.1 mm SL, MCP 45547, 3, 24.9-26.1 mm SL, MPEG 18924, 3, 25.5-27.2 mm SL, MZUSP 91404, 109, 18.5-36.8 mm SL (17, 25.6-36.8 mm SL), 3 c&s, 23.3-26.1 mm SL, USNM 398648, 3, 23.2-27.5 mm SL, all with same data of holotype.
Diagnosis.Moenkhausia eurystaenia is distinguished from all congeners, except M. heikoi and M. phaeonota, by the presence of a dark, broad stripe, from the opercle to the end of the caudal peduncle (vs. stripe, when present, narrow and frequently originating approximately at vertical through dorsal-fin origin). The new species is distinguished from M. heikoi by having 18-21 branched anal-fin rays (vs. 24-26), by the absence of a round blotch at the end of the caudal peduncle (vs. blotch present), and by the dark stripe formed by relatively small chromatophores (vs. large chromatophores). Moenkhausia eurystaenia is distinguished from M. phaeonota (Fig. 3) by its more dispersed dark stripe, not well delimited (vs. less disperse, relatively well delimited), the anterior portion of the stripe darker at horizontal septum (vs. darker below horizontal septum), the deep dark pigmentation along horizontal septum extending anteriorly up to umeral region (vs. extending up to vertical through pelvic-fin origin), the melanophores of interradial membranae between first and fifth branched dorsal-fin rays restricted to the edge of lepidotrichia (vs. spread over interradial membranae), the distance between snout to pelvic-fin origin 46.9-50% SL, n = 35 (vs. 43.2-46.6, n = 30), the distance from snout to anal-fin origin 62.1-65.4% SL, n = 35 (vs. 55.9-61.2, n = 30), the anal-fin base length 25.1-28.4% SL, n = 35 (vs. 28.5-33.8, n = 30), and the number of vertebrae 34, n = 13 (vs. 36-37, n = 12).
Description. Morphometrics in Table 1. Largest specimen examined 36.8 mm SL. Body compressed, greatest body depth slightly ahead of vertical through dorsal-fin origin. Dorsal profile of body convex from upper lip to vertical through anterior edge of nostril; straight from latter point to tip of supraoccipital spine; convex from supraoccipital spine to dorsal-fin origin; straight to slightly convex and posteroventrally inclined along dorsal-fin base; straight to slightly convex from posterior terminus of dorsal-fin base to adipose-fin origin; slightly concave along caudal peduncle. Ventral profile of body convex from tip of lower jaw to pelvic-fin origin; straight from pelvic-fin origin to anal-fin origin; straight and posterodorsally inclined along anal-fin base and slightly concave along caudal peduncle.
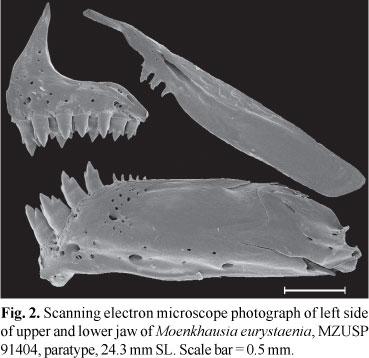
Jaws vertically aligned, mouth terminal. Premaxillary teeth in two rows, outer with 3*(13) or 4(22) tricuspid teeth, inner with five tri- to pentacuspid teeth. Maxilla reaching vertical through anterior fourth of orbit, with 2(14), 3*(17), or 4(1) uni- to tricuspid teeth. Dentary with three pentacuspid teeth, followed by smaller tricuspid and series of 5-10 small conical teeth decreasing gradually in size (Fig. 2).
Pectoral-fin rays i,10*(30), 11(4), or 12(1), reaching pelvic-fin origin in adults, not reaching in juveniles, when adpressed. Pelvic-fin rays i,7, reaching pelvic-fin origin in adults, not reaching in juveniles, when adpressed. Anal-fin rays iv(2) or v(1), and 18(6), 19(13), 20*(13), or 21(3); last unbranched and first four or five branched anal-fin rays longer then remaining rays. Supraneurals 4, all rod-shaped, with bony lamellae in upper portion. Dorsal-fin rays ii,9, first unbranched dorsal-fin ray almost half length of second unbranched ray. Dorsal-fin origin slightly ahead midbody, located at vertical through anterior third of pelvic fin; base of last dorsal-fin ray at vertical through anal-fin origin. First dorsal-fin pterygiophore behind neural spine of 9th vertebra. Adipose-fin origin situated approximately at vertical through 14th to 16th branched anal-fin rays. Caudal-fin rays i,17,i*(32) or i,16,i(2). Caudal fin forked, with lobes of similar size. Dorsal procurrent caudal-fin rays 11, ventral procurrent caudal-fin rays 10(1) or 11(2).
Lateral line completely pored, slightly curved ventrally, with 32(3), 33(9), 34*(8), or 35(4), perforated scales. Longitudinal scale rows between dorsal-fin origin and lateral line 5; longitudinal scale rows between lateral line and pelvic-fin origin 3. Single row of 9*(15), 10(19) or 11(1) predorsal scales. Single row of 3-5 scales overlying base of anteriormost anal-fin rays. Scale rows around caudal peduncle 14. Small scales along first third of upper caudal-fin lobe and first half of lower lobe.
Vertebrae 34(13). Branchiostegal rays 4. First gill arch with 17(2) or 19(1) gill rakers: six on epibranchial, one on intermediate cartilage, eight(2) or 10(1) on ceratobranchial, and two on hypobranchial. Gill rakers without small denticles.
Color in alcohol. Overall ground color yellowish. Infraorbital, opercular and gular areas silvery. Dorsal portion of head and body dark. Dark chromatophores scattered at infraorbital and opercular areas, upper and lower jaws. First three dorsal horizontal scale rows on body with slightly reticulated pattern, formed by scales bordered by dark pigment. Faint pale stripe posterodorsally from eye to caudal peduncle. A broad stripe on body, from opercle to end of caudal-peduncle, darker over horizontal septum, fading ventrally, and extending over middle caudal-fin rays as faint stripe. Smaller specimens with clearer stripe. Narrow midlateral dark stripe at horizontal septum conspicuous, formed by deeper dark chromatophores, extending from humeral region to end of caudal peduncle. Concentration of dark chromatophores immediately posterior to opercle, on first to fourth lateral scales, forming inconspicuous triangular humeral blotch. Scattered dark chromatophores above anal-fin base. All fins with dark chromatophores along edge of lepidotrichia.
Distribution.Moenkhausia eurystaenia is known from the type-locality in the rio Ronuro drainage, upper rio Xingu basin, Brazil (Figs. 4-5).
Etymology. The specific name eurystaenia comes from the Greek eurys, broad, and the Latin taenia, band, a reference to the broad dark stripe.
Discussion
Several authors agree that the species currently included in the genus Moenkhausia most likely do not form a monophyletic group (Fink, 1979; Costa, 1994; Lucena & Lucena, 1999; Lima et al., 2007; Benine et al., 2009). According the most comprehensive phylogenetic analysis of Characidae to date (Mirande, 2009), the analyzed species of Moenkhausia [M. dichroura (Kner), Moenkhausia cf. intermedia Eigenmann, M. sanctaefilomenae (Steindachner), and M. xinguensis (Steindachner)], compose a monophyletic group with Bario steindachneri (Eigenmann). Nonetheless, the synapomorphies of most characid clades, including the synapomorphies of Moenkhausia, are not shown in the appendix 3.
The species of Moenkhausia have a great variety of body shapes, including deep bodied forms as Moenkhausia chrysargyrea (Günther), M. comma Eigenmann and M. doceana (Steindachner), and shallow bodied as M. collettii (Steindachner), M. lopesi Britski & Silimon and the new species. They also exhibit distinct color patterns, such as an overall dark reticulated pattern [i.e. Moenkhausia oligolepis (Günther) and M. sanctaefilomenae]; presence of a dark blotch on the upper caudal-fin lobe [i.e. Moenkhausia lepidura (Kner) and Moenkhausia gracilima Eigenmann]; and dark zigzag lines on body [i.e. Moenkhausia rara Zarske, Géry & Isbrücker and M. simulata (Eigenmann)]. Moenkhausia eurystaenia shares with Moenkhausia phaeonota and M. heikoi, both also occuring in the rio Xingu basin, a dark broad stripe on body, unique in the genus, but also found in other Tetragonopterinae (sensu Géry, 1977), i.e. Tucanoichthys tucano, and some Hyphessobrycon species.
Lima & Zuanon (2004) characterized three types of dark longitudinal bands on body of small characids, without purporting homology: narrow and straight such as in Hyphessobrycon borealis Zarske, Le Bail & Géry, H. heterorhabdus (Ulrey), and H. stegemanni Géry, broad and curve as in Hemigrammus skolioplatus Bertaco & Carvalho and Nematobrycon palmeri Eigenmann, and broad and straight as presented by Moenkhausia eurystaenia and also found in Hyphessobrycon loretoensis Ladiges, H. metae Eigenmann & Henn, H. peruvianus Ladiges, H. notidanos Carvalho & Bertaco, H. herbertaxelrodi Géry, Tucanoichthys tucano, Moenkhausia phaeonota, and M. heikoi Fink (1979) suggested a close relationship between Moenkhausia phaeonota and H. loretoensis, H. metae, H. peruvianus, and perhaps H. agulha and H. herbertaxelrodi. These species share a dark broad and straight stripe, which generally extends from the eye to the caudal-fin base, usually more conspicuous above the anal-fin base. These species also present a pale longitudinal line immediately above the dark stripe, and a concentration of dark chromatophores near the opercle. This color pattern is similar to the one found in Moenkhausia eurystaenia, except by the fainter overall black pigmentation, presented by the later.
Comparative material.Hyphessobrycon cf. agulha: MZUSP 81299, 5, 22.2-30.3 mm SL, rio Negro basin. Hyphessobrycon herbertaxelrodi: MZUSP 96656, 72, 15.1-20.5 mm SL, rio Paraguay basin. Hyphessobrycon loretoensis: MZUSP 85591, 3, 22.0-24.4 mm SL, rio Ucayali basin. Hyphessobrycon metae: MZUSP 50095, 27, 20.5-31.5 mm SL, rio Tapajós basin. Hyphessobrycon notidanos: MZUSP 89645, 15 paratypes, 12.6-35.1 mm SL, rio Tapajós basin. Hyphessobrycon peruvianus: MZUSP 85593, 1, 40.1 mm SL, rio Ucayali basin. Moenkhausia heikoi: MZUSP 82460, 3, 29.5-33.2 mm SL, paratypes of Astyanax dnophos, rio Xingu basin. Moenkhausia phaeonota: MZUSP 78732, 34, 16.0-28.6 mm SL, MZUSP 82292, 1, 33.5 mm SL, rio Paraguay basin; MZUSP 13793, holotype, 32.3 mm SL, MZUSP 13794, 1 paratype, 19.3 mm SL, MZUSP 13795, paratype, 20.7 mm SL, MZUSP 45301, 193, 19.1-43.7 mm SL (7, 28.9-42.9 mm SL), 2 c&s, 29.1-36.2 mm SL, MZUSP 61043, 10, 18.8-37.6 mm SL (5, 33.8-37.9 mm SL), MZUSP 98941, 20, 19.4-36.1 mm SL, rio Tapajós basin; MZUSP 91403, 197, 17.3-31.3 mm SL (15, 24.1-31.0 mm SL), MZUSP 91094, 6, 32.0-39.3 mm SL, MZUSP 98988, 4, 18.7-34.3 mm SL, rio Xingu basin. Nematobrycon palmeri: MZUSP 62432, 1, 36.7 mm SL, aquarium specimen. Tucanoichthys tucano: MZUSP 51322, 10 paratypes, 12.6-15.4 mm SL, rio Negro basin.
Acknowledgements
Naércio A. Menezes and José L. Birindelli (MZUSP) for comments on the manuscript; José L. Birindelli, Osvaldo T. Oyakawa (MZUSP), Cristiano R. Moreira (UNIFESP), Maria I. Landim (MZUSP), Alexandre K. Oliveira (UFSCAR), Aléssio Datovo (LIRP), and José C. Nolasco (MZUSP) for collecting the new species; Leandro M. Sousa (MZUSP) for helping on the preparation of fig. 4; and José L. Birindelli for providing the fig. 5. FAPESP for financial support (09/15075-0).
Literature Cited
Accepted August 20, 2010
Published September 24, 2010
- Benine, R. C., T. C. Mariguela & C. Oliveira. 2009. New species of Moenkhausia Eigenmann, 1903 (Characiformes: Characidae) with comments on the Moenkhausia oligolepis species complex. Neotropical Ichthyology, 7(2): 161-168.
- Costa, W. J. E. M. 1994. Description of two new species of the genus Moenkhausia (Characiformes: Characidae) from the central Brazil. Zoologischer Anzeiger, 232: 21-29.
- Eigenmann, C. H. 1903. New genera of South American fresh-water fishes and new names for some old genera. Smithsonian Miscellaneous Collections, 45: 144-148.
- Eigenmann, C. H. 1917. The American Characidae Part I. Memoirs of the Museum of Comparative Zoology, 43: 1-102.
- Eschmeyer, W. N. (Ed.). 2010. Catalog of Fishes. Electronic version - 12 July 2010. California Academy of Sciences. Avalaible at: http://research.calacademy.org/ichthyology/catalog/fishcatmain.asp Accessed August 19, 2010.
- Fink, W. L. 1979. A new species of Moenkhausia from the Mato Grosso region of Brazil (Pisces: Characidae). Breviora, 450: 1-12.
- Fink, W. L. & S. H. Weitzman. 1974. The so-called Cheirodontin fishes of Central America with description of two new species (Pisces, Characidae). Smithsonian Contributions to Zoology, 172: 1-46.
- Géry, J. 1977. Characoids of the World. T. F. H. Publications, Neptune City, NJ, 672p.
- Lima, F. C. T., L. R. Malabarba, P. A. Buckup, J. F. P. Silva, R. P. Vari, A. Harold, R. C. Benine, O. Oyakawa, C. S. Pavanelli, N. A. Menezes, C. A. S. Lucena, M. C. S. L. Malabarba, Z. M. S. Lucena, R. E. Reis, F. Langeani, L. Casatti, V. A. Bertaco, C. Moreira & P. H. F. Lucinda. 2003. Genera incertae sedis in Characidae. Pp. 106-169. In: Reis, R. E., S. O. Kullander & C. J. Ferraris Jr (Eds.). Check List of Freshwater Fishes of South and Central America. Porto Alegre, Edipucrs, 729p.
- Lima, F. C. T. & J. Zuanon. 2004. A new species of Astyanax (Characiformes: Characidae) from the rapids of the lower rio Xingu, Brazil. Neotropical Ichthyology, 2(3): 117-122.
- Lima, F. C. T., H. A. Britski & F. A. Machado. 2007. A new Moenkhausia (Characiformes: Characidae) from central Brazil, with comments on the area relationship between the upper rio Tapajós and upper rio Paraguay systems. Aqua, International Journal of Ichthyology, 13(2): 45-54.
- Lucena, Z. M. S. & C. A. S. Lucena. 1999. Moenkhausia tergimacula, a new species from the upper rio Tocantis, Brazil (Osteichthyes: Characidae). Ichthyological Exploration of Freshwaters, 10(3): 231-236.
- Marinho, M. M. F. & F. Langeani. 2010. A new species of Moenkhausia from the rio Amazonas and rio Orinoco basins (Characiformes: Characidae). Zootaxa, 2577: 57-68.
- Mirande, J. M. 2009. Weighted parsimony phylogeny of the family Characidae (Teleostei: Characiformes). Cladistics, 25: 1-40.
- Taylor, W. R. & G. C. van Dyke. 1985. Revised procedures for staining and clearing small fishes and other vertebrates for bone and cartilage study. Cybium, 9(2): 107-119.
Publication Dates
-
Publication in this collection
04 Nov 2010 -
Date of issue
2010