
Abstracts
Floating litter banks are an ephemeral habitat consisting of branches, twigs, flowers, seeds, and fruits that are trapped on the stream water surface by a variety of retention mechanisms. These heterogeneous materials form a deep layer of dead plant matter that is colonized by a variety of organisms, including fish that forage on the aquatic macroinvertebrates found in this unique habitat. In this study, we aimed to characterize which fish species occupy the floating litter banks and their trophic characteristics, as well as determine if fish assemblage composition and species richness can be predicted by the size of the floating litter banks. Fish sampling was conducted in five rivers located in the Amazon basin. Of the 31 floating litter banks sampled that contained fish, 455 individuals were recorded and were distributed within 40 species, 15 families and five orders. Siluriformes were the most representative order among the samples and contained the largest number of families and species. The fish fauna sampled was mainly composed of carnivorous species that are typically found in submerged litter banks of Amazonian streams. The fish assemblage composition in the kinon can be predicted by the volume of the floating litter banks using both presence/absence and abundance data, but not its species richness. In conclusion, kinon banks harbor a rich fish assemblage that utilizes this habitat for shelter and feeding, and may function as a refuge for the fishes during the peak of the flooding season.
Ephemeral habitats; Fish assemblages; Insectivores; Lotic systems; Trophic structure
Bancos de folhiço flutuantes são habitats efêmeros constituídos de troncos e galhos de árvores, flores, sementes e frutos retidos por uma variedade de mecanismos. Estes materiais formam uma camada espessa de matéria vegetal morta, que é colonizada por diferentes organismos, incluindo peixes que se alimentam de macroinvertebrados aquáticos presentes neste habitat. Nosso objetivo foi caracterizar as espécies e características tróficas dos peixes que ocupam esses bancos de folhiço flutuantes, bem como determinar se a composição da assembleia de peixes e a riqueza de espécies podem ser previstas pelo tamanho dos bancos. A amostragem dos peixes foi realizada em cinco rios da bacia amazônica. Dos 31 bancos de folhiço flutuantes amostrados que continham peixes, foram registrados 455 indivíduos distribuídos em 40 espécies, 15 famílias e cinco ordens. Siluriformes foi a ordem mais representativa, com o maior número de espécies e famílias. A fauna de peixes coletados foi composta principalmente por espécies carnívoras, que geralmente são encontrados em bancos de folhiço submersos de igarapés amazônicos. A composição das assembleias de peixes do 'kinon' pode ser prevista pelo volume desses bancos de liteira flutuante, tanto utilizando dados de presença/ausência quanto de abundância, mas não de riqueza de espécies. Em conclusão, os bancos de folhiço flutuante abrigam uma rica assembleia de peixes, que utilizam esse habitat como local de abrigo e de alimentação, e podem funcionar como refúgio para a ictiofauna dos bancos de folhiço submerso durante a estação chuvosa.
Introduction
Lotic environments in the Amazon harbor a high diversity of aquatic habitats and allow for the coexistence of a large number of fish species (e.g., Lowe McConnell, 1987; Sabino & Zuanon, 1998; Mendonça et al., 2005). The dense and rich riparian forests that surround the streams and rivers provide a continuous input of organic material to the aquatic systems and form a key source of structure and food in these habitats (e.g. Goulding, 1980). Material inputs from the riparian forests include tree trunks, branches, twigs, roots, leaves, flowers, seeds and fruits, as well as many kinds of invertebrates that fall from the trees or are swept from the stream margins during rain events (Angermeier & Karr, 1984; Lowe McConnell, 1987; Nessimian et al., 1998; Sabino & Zuanon, 1998). Despite the considerable amount of food material that is consumed by the aquatic fauna, most of the organic debris is deposited on the stream bottom or drifts downstream until it becomes trapped by a variety of retention mechanisms (Fig. 1). Such floating or partially submerged litter banks either accumulate upstream on tree branches hanging from the riparian vegetation or stay anchored by fallen tree trunks near the water surface, where they may form a deep layer of dead plant material (Fittkau, 1977).
Floating litter banks of different dimensions and means of accumulation (i.e., diverse retention mechanisms): (a) floating litter banks retained by branches of riparian vegetation; (b) a large floating litter bank retained in a "ria lake" condition at the confluence of the Aliança Stream with the Branco River; (c) a small floating litter bank close to the margin of a 4th-order stream in the Urubu River basin; and (d) a large floating litter bank almost completely out of the water in a stream in the Urubu River basin during the dry season.
Despite their putative temporal instability, such habitats are colonized by a variety of benthic, pelagic and nektonic organisms and were given the name "kinon" by Fittkau (1977). This author described the fauna of a kinon to be mostly aquatic macroinvertebrates and fungi and microorganisms that inhabit its superficial layers. Fittkau (1977) also mentioned the occurrence of fish species that could occupy the kinon banks and subsist on their macroinvertebrate fauna. However, he did not present details on fish species composition or provide empirical evidence of trophic relationships. "Kinon banks" are floating litter banks, which suggest that their fish assemblages should be very similar to those found in the submerged marginal litter banks of streams (e.g., Henderson & Walker, 1986, 1990). Nevertheless, the fish species composition and ecological characteristics of floating litter banks remain poorly understood.
Given that floating litter banks tend to increase in size (surface cover) over time, their areas may correlate with species richness of associated fish assemblages. Considering the dynamics of litter accumulation associated with the retention mechanisms, one may suppose that the size of a floating litter bank may be used as a surrogate for its age. Therefore, the area of floating litter banks may affect the species composition of their fish assemblages. Known species-area relationships allow for the prediction of higher species richness in larger floating litter banks (see Drakare et al., 2006, for a recent review on this subject).
During recent fieldwork to study fish assemblages along longitudinal gradients in Amazonian streams, we observed several floating litter banks, mainly in larger (4th and 5th order) streams. Initial sampling suggested that a characteristic fish assemblage could be associated with these microhabitats. In this study, we described the taxonomic and trophic composition of fish assemblages occupying the floating litter banks, as well as tested if fish assemblage composition and species richness can be predicted by the size of the floating litter banks.
Material and Methods
Study area and general sampling procedures. Between August 2006 and May 2007, 46 spatially discrete floating litter banks were randomly selected and sampled in the following 4th and 5th order rivers of the Amazon basin: Copacá River (N = 10), Aliança Stream, a tributary of Branco River (N = 2), Cuieiras River (N = 10), Preto da Eva River (N = 14), and Urubu River (N = 10). The Copacá (03º21'02.6"S 65º45'54.7"W) is a 25m - wide river that lies within the eastern limit of the 'Reserva Extrativista do Baixo Juruá', which is a protected area of the lower Juruá River (a southern tributary of the Amazon River). The Cuieiras River (02º32'10.2"S 60º19'33.1"W) is a western tributary of the Negro River, and our sampling sites were located near its narrow headwaters (18m - wide) in the southern portion of the Parque Estadual do rio Negro, a protected area near Manaus, where the river is narrow (~20m). The Preto da Eva (02º44'26.8"S 59º40'17.1"W) and the Urubu rivers (02º07'33.6"S 59º56'53.6"W) are northern tributaries of the Amazon River just east of Manaus, where the samplings were done in narrow stretches (7-15m - wide) of the river channel. Both sampling sites in the Branco River basin (a major northern tributary of the Negro River) were located in the Aliança Stream (01º27'51.2"N 61º14'46.8"W; channel width = 20-30m), in more lentic, "ria lake" conditions (Irion, 1984), in Roraima State. Aside from the above noted difference for Aliança Stream, all other collecting sites show very similar general environmental conditions: narrow channels of moderate current speed with pristine riparian forests in the margins, sampled during the flooding season.
Data collection and fish sampling. The size of each kinon (using the terminology suggested by Fitkau, 1977) bank was recorded by measuring its maximum width, length and depth with a measuring tape. The measurements were made by gently placing the tape over the surface of the kinon bank in order to avoid frightening the fishes and disrupting the integrity of the litter accumulation. These data were later used to estimate the volume (m3) of each floating litter bank based on the formula for an ellipsoid structure (Equation 1):
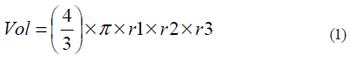
where r1, r2 and r3 represent width, length and depth of the floating litter bank, respectively.
After measuring the dimensions of a floating litter bank, we sampled its associated fish assemblage using fine mesh (1-mm2 aperture) hand nets. Fish specimens were euthanized using a lethal dose of clove oil (Eugenol, cf. American Veterinary Medical Association, 2001) and were immediately preserved in a 10% formalin solution.
Small floating litter banks were sampled completely, but larger ones were subsampled. We standardized our sample data by estimating the proportion of each floating litter bank and extrapolating it to the whole; this allowed us to standardize the sampling effort and make direct quantitative comparisons among the samples. The quantitative data about the fish fauna of the floating litter banks were combined with natural history information gathered during fish faunal inventories and ecological studies that we conducted in several river systems of the Brazilian Amazon (e.g., Carvalho et al., 2006; Sazima et al., 2006; Zuanon et al., 2006; Zuanon & Ferreira, 2008; Carvalho et al., 2009). Based on these surveys, we generated a comprehensive list of fish known to be associated with floating litter banks. Voucher specimens were deposited in the fish collection at the Instituto Nacional de Pesquisas da Amazônia (INPA), Manaus, Amazonas State, Brazil.
Trophic groups. In order to understand the role of kinon banks in the natural history of fishes, we have classified each fish species in broad trophic categories depicting the main food source (e.g. invertebrates, fruits, algae) and its origin (allochthonous or autochthonous). To accomplish this, dietary analyzes using the frequency-of-occurrence method were performed for most of the fish species represented in our samples (1 - 20 specimens/species). This method divides the number of times that a given food item is observed by the total number of examined stomachs that contained food. The species were then classified into a trophic group based on the most important food items present in their diet (food items with frequencies of occurrence of 60% or higher). Species for which few or single individuals were collected were assigned to trophic categories based on information available for closely related species with similar ecological habits, or based on unpublished data obtained by JZ and LNC for those species. This was the case for Steatogenys ocellatus, which dietary information was based on data gathered for Steatogenys duidae.
Data analysis. Differences in the number of floating litter banks sampled at each study site, as well as in the sampling effort (some banks were examined completely for fish, whereas others had only a fraction of their material sorted) could generate bias in the species-richness estimates for the different study sites. To correct for this problem, we employed rarefaction curves based on the number of studied samples (cf. Gotelli & Colwell, 2001) and applied the Mao Tau index (EstimateS 8, Colwell et al., 2004; Colwell, 2005). Rarefaction methods are considered adequate to generate estimates of species richness when comparing samples with different numbers of individuals (Gotelli & Colwell, 2001), which could be expected as a consequence of the differences in the size of the sampled kinon banks.
Despite the procedures employed to minimize the possible bias generated by the differences in sampling effort, we choose not to include the Branco River samples in the statistical models presented below. Some exploratory analyses showed that the two Branco River samples were strong outliers in the quantitative analyses (based on the abundance of individuals). Nevertheless, the presence/absence data from those samples were used to construct a similarity matrix among all the sampled river basins with the use of Jaccard's coefficient index (Krebs, 1999).
The dimensionality of the fish assemblage composition was reduced using Principal Coordinates Analysis (PCoA) without inclusion of the Branco River sampling units. Standardization we applied involved a transformation of each species' total abundance to their relative abundance in each sampled floating litter banks. The PCoA association matrix was constructed using Bray-Curtis distance for quantitative (abundance) data and the Sørensen index for qualitative (presence/absence) data (Legendre & Legendre, 1998). The quantitative data revealed patterns based on the most common species, which tended to have the greatest influence on the quantitative differences among sites. Presence/absence data usually give more weight to uncommon species, as common species occur in most sites and thus contribute little to qualitative differences among sites. The first three components of the PCoA analyzes (for qualitative and quantitative data) explained more than 50% of the variance and were retained for subsequent analyzes. The fish assemblages, represented by these multivariate axes, were used as dependent variables in a Multivariate Regression Analysis, in which the volumes of the floating litter banks sampled were used as predictor variables. To determine whether the species richness of the fish assemblages associated with the floating litter banks could be predicted by the volume of the samples, we applied a Generalized Linear Model with a Poisson error distribution and used the number of species found in each floating litter bank (raw species counts) as the dependent variable. The use of this technique was chosen in function of the non-normal distribution of the data. Further, to analyze the effect of the basins on the relation between fish sample composition and kinon bank area, we applied a Partial Redundancy Analysis and a Non-Metric Multidimensional Scaling followed by a Multivariate Analysis of Covariance (using the first two NMDS axes as dependent variables and basin as a categorical variable). The possible effect of the different basins on species richness in the kinon samples was tested with separate Pearson's correlations. Multivariate Regression Analysis was undertaken using SYSTAT. The ordinations and GLM richness model, RDA, NMDS and MANCOVA were proceeded with R statistical analysis software (R Development Core Team, 2011) using the vegan package (Oksanen et al., 2011).
Results
Of the 46 floating litter banks sampled, 15 did not contain fish and were excluded from the analyses. The remaining 31 samples contained 455 fish individuals distributed in 40 species, 15 families and five orders (Table 1). The observed/estimated species richness in the sampled floating litter banks varied among the river basins (Copacá = 15/26; Urubu = 9/11; Cuieiras = 7/12; Rio Preto da Eva = 7/11; Branco = 18/not estimated - see Methods). The order Siluriformes had the largest number of families and species in the samples (Table 1). The rarefaction curves indicate that our sampling effort was insufficient (Fig. 2), which may be reflected in the differences found in the fish faunas associated to the kinon banks of the five studied river stretches.
Species composition, abundance, and trophic categories of fishes in the floating litter banks samples of 4th and 5th order streams of five river basins in the Brazilian Amazon
Rarefaction curve (Mao Tau Index) based on the number of samples collected in floating litter banks in four river basins in the Brazilian Amazon (Copacá, Cuieiras, Preto da Eva, and Urubu rivers). (a) All floating litter banks recorded, and (b) disentangling rarefaction models for each river basin.
Most of the species found occurred in only one or two basins (e.g., Elachocharax pulcher, Crenuchidae, occurred in four basins and Brachyhypopomus sp.1 occurred in three; Table 1, Fig. 3). This fact highlights the low overall similarity in the fish assemblage composition among the samples of the different drainages. The highest similarity values were observed for the Cuieiras and Urubu Rivers (23%), followed by the Cuieiras and Preto da Eva Rivers (15.4%), and the Cuieiras and Copacá Rivers (14.3%). The lowest similarities involved the Urubu and Copacá Rivers (4.3%) and the Preto da Eva and Copacá Rivers (4.5%). The Branco River samples presented low similarity values in relation to all other studied basins (8.3-8.7%), and the highest similarity involving the Branco River was with the Copacá River (13%).
The fish fauna associated with kinon banks is dominated by species of characins (Characiformes), catfishes (Siluriformes) and electric knife fishes (Gymnotiformes). Some of the more common and abundant species in our samples were the following: (a) Elachocharax pulcher, Crenuchidae; (b) Hypopygus lepturus, Hypopomidae; (c) Batrochoglanis raninus, Pseudopimelodidae; and (d) Brachyglanis microphthalmus, Heptapteridae.
A family-level comparison of the fish assemblages of the floating litter banks revealed a high similarity among the samples of the Cuieiras and Urubu Rivers (83.3%), followed by the Branco and Copacá Rivers (69.2%). The lowest similarity occurred in the Preto da Eva and Cuieiras River (22.2%) samples. Crenuchidae and Hypopomidae were the only families that occurred in all of the river basins (Table 1, Fig. 3).
The fish fauna sampled in the floating litter banks was composed predominantly of species typically found in submerged litter banks of Amazonian streams (29 out of 40 species, or 72.5%; our personal observations). These include species of the families Erythrinidae, Hypopomidae, Rhamphichthyidae, Auchenipteridae, Doradidae, Cetopsidae, Heptapteridae, Trichomycteridae, and Pseudopimelodidae. Most of these species are carnivores (Table 1). More specifically, they are autochthonous insectivores that consume immature insects of the families Chironomidae (Diptera), Baetidae, and Leptophlebidae (Ephemeroptera). Only Moenkhausia collettii (Characidae), Parauchenipterus galeatus (Auchenipteridae) and Helogenes marmoratus (Cetopsidae) were found to consume insects of allochthonous origin (Table 1). A single specimen of the piscivorous characin Hoplias malabaricus (Erythrinidae) was found in the floating litter banks samples.
The quantitative ordination of the fish community captured 53%, and the qualitative ordination captured 52%, of the variation of the original distances in three dimensions. The mean volume of the floating litter banks sampled in the rivers Copacá, Cuieiras, Rio Preto da Eva and Urubu was 0.24 m3 (0.001 - 1.23 m3), and the Branco River banks contained a mean volume of 9.2 m3 (6.4 - 12.0 m3, not used in statistical analysis). The fish assemblage composition by presence/absence (Pillai Trace = 0.29, F3, 25 = 3.44, p = 0.03) and abundance data (Pillai Trace = 0.30, F3, 25 = 3.51, p = 0.03) can be predicted by the volume of the floating litter banks, but species richness cannot be predicted by their volume (Null deviance = 27.19, Residual deviance = 25.16, z = 1.50, p = 0.13). Correlation analyzes conducted separately for each basin revealed a significant relation between species richness and litter bank volume only for the Cuieiras samples (r= 0.97; Bartlett's Chi-square statistic = 7.111, df = 1, p = 0.008). Finally, both Partial RDA analysis and MANCOVA did not reveal significant effects of the basin on the relation between fish species composition and kinon bank area.
Discussion
Most of the fish that occupy the floating litter banks have sedentary habits and are commonly found in submerged litter banks that accumulate along stream banks or in backwaters (Henderson & Walker, 1986, 1990). Such submerged litter banks are frequently washed away during the rapid floods that typically occur during the rainy season, or they may develop unsuitable or less favorable environmental conditions for the fish fauna (e.g., low oxygen levels, or smaller prey availability). Thus, the floating litter banks studied here may represent an important refuge for the fish fauna of submerged litter banks in larger (e.g., 4th- to 5th-order) streams during the high-water season. In fact, our results demonstrate that the fish fauna observed in the floating litter banks includes species that require strong water currents and high oxygen concentrations to survive. Fittkau (1977) argued that despite the relatively low species richness of the fauna (insects and other microorganisms) of floating litter banks (in comparison with other lotic environments), their inhabitants seem to occupy the kinon for long periods (mainly during the flooding season). Such floating-litter-bank dwellers even include an apparently exclusive species, namely the belostomatid water bug Weberiella rhomboides (JL Nessimian, pers. comm.), which corroborates the importance of this kind of microhabitat for several invertebrates and fish species in Amazonian streams and rivers.
Our results have shown that the vast majority of fish species present on floating litter banks is composed of autochthonous insectivores, which probably forage for prey among the organic debris that compose the kinon. Aquatic insect larvae possibly also benefit from the high oxygen content of the upper layers of the stream channel where kinon banks are anchored, as well as from the shelter and foraging grounds provided by the complex structure of that microhabitat. Thus, the species-rich insect fauna that inhabit such banks (Fidélis, Carvalho & Zuanon, unpublished data) possibly provide a favorable habitat for foraging by micro-carnivorous fish, especially during the flooding season when the stream volume may increase dramatically and frequently disturbing the bottom substrate.
The low similarity of the fish fauna of the floating litter banks, at least when analyzed at the species level, may reflect the randomness of the process of accumulation of floating litter banks as well as possible differences in the colonization time of the banks. For example, Elachocharax pulcher occurred just in a small number of the basins included in this study, but the species is widely distributed in leaf litter habitats across the Amazon lowlands (Buckup, 1993, 2003). On the other hand, the overall similarity increased when assessed by family, which suggests that important geographic components influence the composition of the fish fauna of floating litter banks. In this sense, fish composition probably reflects the regional species pool, the distance relationships among the studied basins, and the different levels of connectivity among them, as well as the effects of the limnological characteristics of the streams and rivers. Overall, the fish fauna of floating litter banks is composed by species that typically inhabit submerged litter banks of streams, and do not constitute a simple random subsample of the ichthyofauna present in the river or basin.
Henderson & Walker (1986) state that the litter banks supply the benthic community (including the fish fauna) with food and shelter, especially during the dry season when other habitats are unavailable. Thus, the structural change resulting from the seasonal disintegration of bottom litter banks and the formation of floating litter banks may also lead the fishes to move from the bottom to the surface. This alternative habitat seems to be especially important for the juveniles of several species such as the pseudopimelodid catfishes of the genus Batrochoglanis, the hypopomid electric fishes of the genera Brachyhypopomus and Hypopygus, and erythrinids (our personal observations).
Henderson & Walker (1990) report on the existence of vertical stratification in the occupation of submerged litter banks by fish; such stratification also seems to occur among the fish species of floating litter banks. Our observations during the sampling of floating litter banks have shown that some electric fishes (Hypopomidae) and several catfishes (species of Auchenipteridae, Doradidae, Cetopsidae, Heptapteridae, Trichomycteridae, and Pseudopimelodidae) occupy the inner portions of the floating litter banks. Several of these species show adaptations for living under low oxygen conditions, which possibly allow them to occupy confined spaces inside the banks. On the other hand, several species were observed at the periphery of the floating litter banks and may benefit from the stronger currents and higher oxygen levels that result from the positioning of the floating litter banks at the water's surface. This is the case for the crenuchids Ammocryptocharax elegans (Zuanon et al., 2006) and Melanocharacidium dispilomma (our pers. obs.), the cetopsid Helogenes marmoratus (our pers. obs.), the hypopomids Steatogenys spp. (Sazima et al., 2006), and the loricariid Acestridium discus (Retzer et al., 1999). These species inhabit fast-flowing stretches of streams and maintain their positions on plant leaves, twigs and branches that are continuously washed by the current. Finally, some fish apparently seek refuge under floating litter banks and do not enter the dense entanglements of twigs, leaves, and other organic material that comprises the actual banks. These species include some cichlids, the sternopygid electric fishes, the nectonic characins of the genera Moenkhausia and Hemigrammus (and possibly many others), and the cyprinodont Rivulus ornatus that is commonly found close to the water's surface at stream margins.
Fittkau (1977) suggested that the "age" of the floating litter banks (i.e., the time available for colonization by fishes) is an important factor influencing the density and diversity of the inhabitants of the floating litter banks (i.e., in recent formations there would be high abundances of relatively few species). The present study has shown that the occurrence of fish fauna and the abundance of individuals could be predicted by the volume of the floating litter banks. Such predictability of occurrence in the kinon banks of certain fish species (or higher taxonomic categories, such as families) reflects the relationships between its natural history characteristics and the environmental constraints of the floating litter banks (e.g., a large volume of decaying organic debris with narrow spaces for fish passage). On the other hand, fish abundance predictability may represent a simple relation with habitat availability.
The lack of area-based increments in fish species richness can be explained by a disproportional relationship between area and heterogeneity. Empirical evidence shows that species-area relationships are absent when increases in area per se do not represent significant increases in habitat heterogeneity (Báldi, 2008); in the case of kinon banks, the increase in area (a square dimension) may be accompanied by a higher increase in volume (a cubic dimension), possibly resulting in inadequate conditions for the fish fauna at the innermost portion of the banks. Thus, habitat heterogeneity in floating litter banks may play an important role in determining species richness (e.g., Kalmar & Currie, 2006; Báldi, 2008), but not as a linear relation.
In his original article, Fittkau (1977) suggested that the seemingly low occurrence and ecological importance of floating litter banks in European rivers could be an effect of historical anthropogenic disturbances. The great reduction in riparian vegetation along Old World rivers and streams along centuries of human occupation may have impaired the formation of floating litter banks and consequently an adequate perception of its importance for the regional fish fauna. Although little is known about the subject, floating litter banks seem to constitute an important microhabitat for the fish fauna of 4th- and 5th-order forest streams in the Amazon. Unfortunately, during our surveys we observed several signs of environmental disturbances caused by human activities. Study sites in streams close to human settlements (such as the Rio Preto da Eva basin) frequently showed the presence of PET (Polyethylene terephthalate) plastic bottles among the floating litter banks. PET bottles currently occupy a large volume in the floating litter banks, but they do not have the structural complexity to be used as shelter for fishes, and they cannot be used as a substrate for the invertebrates that are consumed by the ichthyofauna. Thus, the increasing presence of plastic debris (and other types of pollution as well) in lotic water bodies may mask relations between fish assemblage composition and litter bank volume, and may disrupt the dynamics of the seasonal occupation of floating litter banks by fishes and invertebrates in streams and low-order rivers in the Amazon.
In conclusion, we observed a species-rich fish fauna in floating litter banks. The fish species recorded were mostly small and sedentary and are typical of submerged litter banks, which can disappear or become unsuitable for fish fauna during flooding events. In this case, floating litter banks become a refuge for fish providing shelter and food from associated invertebrates. Our results show that the composition of the fish fauna of floating litter banks can be modeled by species-area predictions, but that these cannot be used for species richness. The floating litter banks that we studied have proven to be important locations for shelter and foraging by fishes. However, our results are only initial attempts at establishing a deeper understanding of the temporal and spatial dynamics of the associated species. We suggest niche-based or random-based hypothesis testing, colonization experiments and/or competitive interaction studies of species associated with floating litter banks (e.g., fish/fish and fish/invertebrate interactions), which would improve our knowledge about the faunal assemblage processes of this interesting microhabitat in streams and rivers of different geographic regions and biomes.
We thank Alberto Akama and Ocírio Pereira for their help with fieldwork. Fabíola ártemis Souza do Valle helped with the identification of food items. Oscar Shibatta checked the identification of the Pseudopimelodidae species. The Fundação Nacional da Saúde (FUNASA), the Instituto de Pesquisas Ecológicas (IPE) and the Instituto Brasileiro do Meio Ambiente e dos Recursos Naturais Renováveis (IBAMA/Programa ARPA) provided logistical support. Fernando P. Mendonça by photos of Figs. 1(b) and 2 (a,b), and Leandro M. Sousa by photos of Fig. 2 (c, d). CNPq, FAPEAM, and the Fundação "O Boticário" de Proteção à Natureza provided essential financial support. This study was supported by fellowships from CNPq to JZ (process 307464/2009-1); graduate fellowships from CNPq and IIEB (BECA program, B/2006/01/BDP/05, the Gordon and Betty Moore Foundation) to LNC. This is contribution number 612 in the BDFFP Technical Series and contribution 28 of Projeto Igarapés.
Literature Cited
- American Veterinary Medical Association. 2001. Report of the AVMA panel on euthanasia. Journal of the American Veterinary Medical Association, 218: 669-696.
- Angermeier, P. L. & J. R. Karr. 1984. Fish communities along environmental gradients in a system of tropical streams. Pp. 39-57. In: Zaret, T. M. (Ed.). Evolutionary ecology of neotropical freshwater fishes. Dr. W. Junk Publishers, The Hague, Netherlands, 184p.
- Báldi, A. 2008. Habitat heterogeneity overrides the species-area relationship. Journal of Biogeography, 35: 675-681.
- Buckup, P. A. 1993. Review of the characidiin fishes (Teleostei: Characiformes), with descriptions of four new genera and ten new species. Ichthyological Exploration of Freshwaters, 4: 97-154.
- Buckup, P. A. 2003. Family Crenuchidae (South American darters). Pp. 87-95. In: Reis, R. E., S. O. Kullander & C. J. Ferraris (Eds.). Check List of the Freshwater Fishes of South and Central America, Porto Alegre, Edipucrs, 729p.
- Carvalho, L. N., J. Zuanon & I. Sazima. 2006. The almost invisible league: crypsis and association between minute fishes and shrimps as a possible defence against visually hunting predators. Neotropical Ichthyology, 4: 219-224.
- Carvalho, L. N., J. Zuanon & I. Sazima. 2009. Natural history of Amazon fishes. Pp. 113-144. In: Del-Claro, K., P. S. Oliveira & V. Rico-Gray (Orgs.). Tropical Biology and Conservation Management: Case studies, Eolss Publishers Co. Ltd., Oxford, 215p.
- Colwell, R. K. 2005. EstimateS: Statistical estimation of species richness and shared species from samples. Version 8.0. User's guide and application published at: http://purl.oclc.org/estimates.
» http://purl.oclc.org/estimates - Colwell, R. K., C. X. Mao & J. Chang. 2004. Interpolating, extrapolating, and comparing incidence-based species accumulation curves. Ecology, 85: 2717-2727.
- Drakare, S., J. J. Lennon & H. Hillebrand. 2006. The imprint of the geographical, evolutionary and ecological context on species-area relationships. Ecology Letters, 9: 215-227.
- Fittkau, E. J. 1977. Kinal and kinon, habitat and coenosis of the surface drift as seen in Amazonian running waters. International Journal of Tropical Geology, Geography and Ecology, 1: 9-20.
- Gotelli, N. J. & R. K. Colwell. 2001. Quantifying biodiversity: procedures and pitfalls in the measurement and comparison of species richness. Ecology Letters, 4: 379-391.
- Goulding, M. 1980. The fishes and the forest - Explorations on Amazonian natural history. Berkeley, University of California Presss, 280p.
- Henderson, P. A. & I. Walker. 1986. On the leaf litter community of the Amazonian blackwater stream Tarumazinho. Journal of Tropical Ecology, 2: 1-17.
- Henderson, P. A. & I. Walker. 1990. Spatial organization and population density of the fish community of the litter banks within a central Amazonian black water stream. Journal of Fish Biology, 37: 401-411.
- Irion, G. 1984. Sedimentation and sediments of Amazonian rivers and evolution of the Amazonian landscape since Pliocene. Pp. 201-214. In: Sioli, H. (Ed.). The Amazon - Limnology and landscape ecology of a mighty tropical river and its basin. Monographiae Biologicae, Vol. 56, Dr. W. Junk Publishers, Dordrecht, 763p.
- Kalmar, A. & D. J. Currie. 2006. A global model of island biogeography. Global Ecology and Biogeography, 15: 72-81.
- Krebs, C. J. 1999. Ecological Methodology. 2nd ed. California, Benjamin/Cummings, 620p.
- Legendre, P. & L. Legendre. 1998. Numerical Ecology: Developments in environmental modeling 20. 2nd english ed. Amsterdam, Elsevier Science B.V., 870p.
- Lowe-McConnell, R. H. 1987. Ecological studies in tropical fish communities. Cambridge, Cambridge University Press, 382p.
- Mendonça, F. P., W. E. Magnusson & J. Zuanon. 2005. Relationships between habitat characteristics and fish assemblages in small streams of Central Amazonia. Copeia, 2005: 751-764.
- Nessimian, J. L., L. F. M. Dorvillé, A. M. Sanseverino & D. F. Baptista. 1998. Relation between flood pulse and functional composition of the macroinvertebrate benthic fauna in the lower Rio Negro, Amazonas, Brazil. Amazoniana, 15: 35-50.
- Oksanen, J., F. Guillaume Blanchet, R. Kindt, P. Legendre, R. B. O'Hara, G. L. Simpson, P. Solymos, M. H. H. Stevens & H. Wagner. 2011. vegan: Community Ecology Package. R package version 1.17-6. http://CRAN.R-project.org/package=vegan
» http://CRAN.R-project.org/package=vegan - Petts, G. E. 1994. Rivers: dynamic components of catchment ecosystems. Pp. 3-22. In: Calow, P. & G. E. Petts (Eds.). The River Handbook. Blackwell Scientific, Oxford.
- R Development Core Team (2011) R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. ISBN 3-900051-07-0, URL http://www.R-project.org.
» URL http://www.R-project.org - Retzer, M. E., L. G. Nico & F. R. Provenzano. 1999. Two new species of Acestridium (Siluriformes: Loricariidae) from southern Venezuela, with observations on camouflage and color change. Ichthyological Exploration of Freshwaters, 10: 313-326.
- Sabino, J. & J. Zuanon. 1998. A stream fish assemblage in Central Amazonia: distribution, activity patterns and feeding behavior. Ichthyological Exploration of Freshwaters, 8: 201-210.
- Sazima, I. , L. N. Carvalho, F. P. Mendonça & J. Zuanon. 2006. Fallen leaves on the water-bed: diurnal camouflage of three night active fish species in an Amazonian streamlet. Neotropical Ichthyology, 4: 119-122.
- Zuanon J., L. N. Carvalho & I. Sazima. 2006. A chamaeleon characin: the plant clinging and colour-changing Ammocryptocharax elegans (Characidiinae: Crenuchidae). Ichthyological Exploration of Freshwaters, 17: 225-232.
- Zuanon, J. & E. Ferreira. 2008. Feeding ecology of fishes in the Brazilian Amazon - a naturalistic approach. Pp. 1-34. In: Cyrino, J. E. P., D. Bureau & B. G. Kapoor (Eds.). Feeding and Digestive Functions of Fishes. Science Publishers Inc., USA, 589p.
-
Published March 31, 2013
Publication Dates
-
Publication in this collection
Jan-Mar 2013
History
-
Received
05 May 2011 -
Reviewed
29 Aug 2012 -
Accepted
26 Dec 2012



