
Abstracts
Neoplecostomus jaguari, a new neoplecostomine, is described based on material from the rio Jaguari, rio Tietê basin, southeastern Brazil. The new species differs from all congeners by having a conspicuous sexual dimorphism: females with 30-38 teeth in the dentary and 30-39 in the premaxilla, and males with 13-18 and 14-20 teeth, respectively (vs. number of teeth in the dentary and premaxilla approximately equivalent in all other species, except for N. langeanii in the premaxilla); females with larger mandibullary width, 18.5-21.6% HL, males 11.8-15.7% HL (vs.mandibullary width in males and females approximately equivalent); females with teeth longer and thin, males with teeth shorter and robust (vs. teeth similar in other species, except for N. langeaniiin the premaxilla). Furthermore the new species is distinguished from all other species from the upper rio Paraná basin by having a lateronasal plate either forming the border of the nostril or posteriorly displaced (vs. lateronasal plate absent). Finally, other useful character includes four to six plates between the frontal and prefrontal, herein called posteronasal plates (vs.one to three in all other species). An identification key to all Neoplecostomus species is provided.
Cascudos; Neoplecostomus paranensis; Neotropical region; Ostariophysi; Systematics
Neoplecostomus jaguari, espécie nova de Neoplecostominae, é descrita com base em material do rio Jaguari, drenagem do rio Tietê, sudeste do Brasil. A espécie nova difere de todas as congêneres por apresentar dimorfismo sexual conspícuo: fêmeas com 30-38 dentes no dentário e 30-39 no pré-maxilar e machos com 13-18 e 14-20 dentes, respectivamente (vs. número de dentes no dentário e pré-maxilar aproximadamente equivalente em todas as outras espécies, exceto N. langeanii para o pré-maxilar); fêmeas com maior largura do mandibular, 18,5-21,6% CC, machos 11,8-15,7% CC (vs. largura do mandibular aproximadamente equivalente em machos e fêmeas); fêmeas com dentes longos e finos, machos com dentes curtos e robustos (vs. dentes similares nas outras espécies, exceto N. langeanii no pré-maxilar). Adicionalmente diferencia-se de todas as demais espécies da bacia do alto rio Paraná por apresentar a placa lateronasal formando a borda lateral da narina ou deslocada posteriormente (vs. placa lateronasal ausente). Finalmente, caracteres úteis para o reconhecimento da espécie nova incluem quatro a seis placas entre o frontal e pré-frontal, aqui denominadas placas posteronasais (vs. uma a três em todas as outras espécies). Uma chave de identificação para todas as espécies de Neoplecostomus é fornecida.
Introduction
Representatives of Neoplecostomus are endemic to the high streams in the Brazilian Cristaline Shield and to date includes 14 valid species distributed in the rio Doce, rio Paraíba do Sul, upper rio Paraná basin, rio Ribeira de Iguape, rio São Francisco, and some other small costal drainages (e.g., Jucu and São Lourenço in the Espírito Santo State). It was first proposed as a subgenus of Plecostomus by Eigenmann & Eigenmann (1888), including the type species Plecostomus microps Steindachner, 1876, from Rio de Janeiro, rio Paraíba do Sul basin, and Hypostomus granosus Valenciennes, 1840, from Cayenne (an error) and Rio de Janeiro. Posteriorly, Eigenmann & Eigenmann (1889) elevated Neoplecostomus to genus, which was latter diagnosed by Langeani, (1990) by having two or three conspicuous rows of enlarged and transversally flattened papillae just posterior to the dentary teeth, larger than those on the remaining portions of the lower lip, and the abdomen covered with conspicuous and small platelets, between insertions of pectoral and pelvic fins, surrounded by naked areas.
Neoplecostomus paranensis Langeani, 1990 until very recently was the unique species described to the upper rio Paraná basin. Zawadzki et al. (2008)Zawadzki, C. H., C. S. Pavanelli & F. Langeani. 2008. Neoplecostomus (Teleostei: Loricariidae) from the upper Rio Paraná basin, Brazil, with description of three new species. Zootaxa, 1757: 31-48. proposed three new species to the area: N. corumba, N. selenae, and N. yapo; few years later, Roxo et al. (2012a)Roxo, F. F., C. Oliveira & C. H. Zawadzki. 2012a. Three new species of Neoplecostomus (Teleostei: Siluriformes: Loricariidae) from the Upper Rio Paraná basin of southeastern Brazil. Zootaxa, 3233: 1-21. described three additional species: N. bandeirante, N. botucatu, and N. langeanii, increasing by almost 100% the total number of known species to the genus and making the upper rio Paraná the richest basin concerning Neoplecostomus diversity. In the present paper, we describe an additional new species to the upper rio Paraná basin, from tributaries of the rio Jaguari, drainage of the rio Tietê, and provide an identification key for the species in the genus.
Material and Methods
Measurements and counts of bilaterally symmetrical characters were made point-to-point with digital calipers, to the nearest 0.1 mm, on the left side of the specimens under a stereomicroscope. Body plate nomenclature follows Schaefer (1997)Schaefer, S. A. 1997. The Neotropical cascudinhos: Systematics and biogeography of the Otocinclus catfishes (Siluriformes: Loricariidae). Proceedings of the Academy of Natural Sciences of Philadelphia, 148: 1-120. and Armbruster (2004)Armbruster, J. W. 2004. Phylogenetic relationships of the suckermouth armored catfishes (Loricariidae) with emphasis on the Hypostominae and the Ancistrinae. Zoological Journal of the Linnean Society, 141: 1-80., and measurements follow Langeani (1990)Langeani, F. 1990. Revisão do gênero Neoplecostomus Eigenmann & Eigenmann, 1888, com a descrição de quatro novas espécies do sudeste brasileiro (Ostariophysi, Siluriformes, Loricariidae). Comunicações do Museu de Ciências, PUCRS, série Zoologia, 3: 3-31. and Zawadzki et al. (2008)Zawadzki, C. H., C. S. Pavanelli & F. Langeani. 2008. Neoplecostomus (Teleostei: Loricariidae) from the upper Rio Paraná basin, Brazil, with description of three new species. Zootaxa, 1757: 31-48.. Osteological preparations (c&s) follow Taylor & Van Dyke (1985)Taylor, W. R. & G. C. Van Dyke. 1985. Revised procedures for staining and clearing small fishes and other vertebrates for bone and cartilage study. Cybium, 9: 107-119.. Sex determination was based primarily on secondary sexual characters: mature males with a dermal flap on the dorsal surface of the pelvic-fin rays (vs. absent in females) and a conspicuous urogenital papilla posterior to anus (vs. absent in females). The gonads were also observed in the specimens, which were latter cleared and stained.
Institutional abbreviations follow Reis et al. (2003)Reis, R. E., S. O. Kullander & C. J. Ferraris Jr. (Orgs.). 2003. Check List of the Freshwater Fishes of South and Central America. Porto Alegre, Edipucrs., with addition of Coleção Ictiológica da Universidade Federal do Espírito Santo (CIUFES); Departamento de Zoologia e Botânica, Universidade Estadual Paulista "Júlio de Mesquita Filho", São José do Rio Preto (DZSJRP); Laboratório de Biologia e Genética de Peixes do Departamento de Morfologia da Universidade Estadual Paulista "Júlio de Mesquita Filho", Botucatu (LBP); Laboratório de Ictiologia de Ribeirão Preto, Universidade de São Paulo, Ribeirão Preto (LIRP); Núcleo de Pesquisas em Limnologia, Ictiologia e Aquicultura, Universidade Estadual de Maringá, Maringá (NUP). Abbreviations used in text are CC (comprimento da cabeça), HL (head length), MW (mandibullary width), and SL (standard length).
Results
Neoplecostomus jaguari, new species
Figs. 1-3

Neoplecostomus jaguari, holotype, LIRP 2277, 89.1 mm SL, male, ribeirão do Forja, rio Jaguari drainage, sub-basin of rio Piracicaba-Capivari-Jundiaí, rio Tietê drainage, Extrema, Minas Gerais State, Brazil.
Neoplecostomus sp. 5: Roxo et al., 2012b: 35 (tab. 1), 38 (fig. 2). -Roxo et al., 2012c: 6 (fig. 2), 7 (fig. 3).

Part of skull of Neoplecostomus jaguari, paratype, LIRP 2278, 64.6 mm SL, male, left side, dorsal view, (specimen dissected on the left side), scale 1 mm. F = frontal; IO1-5 = infraorbitals 1-5; LNP = lateronasal plate; N = nasal; OP = opercle; PF = prefontral plate; PN = prenasal plate; PNP = posteronasal plate; POP = preopercle; PR1-6 = postrostrals plates 1-6; PT-SC = compound pterotic; SOC = supraoccipital; SPC = suprapreopercle; SPH = sphenotic.
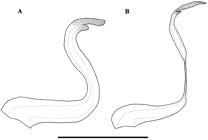
Neoplecostomus jaguari, paratypes, LIRP 2278, replacement dentary teeth, left side, lateral view. A) male, 64.6 mm SL. B) female, 58.8 mm SL. Scale 1 mm.
Morphometric and meristic data for Neoplecostomus jaguari. Holotype (H) and 53 paratypes, 32 females (F), and 21 males (M), range includes holotype, CD = caudal fin, CP = caudal peduncle, ID = interorbital distance, OD = orbital diameter, PDS = predorsal, SD = standard deviation.
Neoplecostomus sp. "Casca d'Antas": Lucena et al., 2012: 326 (fig. 1), 327 (fig. 2), 328 (tab. 1), 329 (tab. 2).
Holotype. LIRP 2277, male, 89.1 mm SL, Brazil, Minas Gerais State, Extrema, ribeirão do Forja, 22°52'20''S 46°17'27''W, 05 Jun 2001, R. M. C. Castro et al.
Paratypes. Brazil, Minas Gerais State, upper rio Paraná basin, rio Tietê basin, rio Jaguari basin. LIRP 2278, 83, 23 males, 35.0-76.3 mm SL, 49 females, 43.7-69.9 mm SL, 7 unsexed, 28.3-32.8 mm SL + 4 c&s, 58.8-65.6 mm SL, (43, 47.6-76.3 mm SL), same data of holotype. MZUSP 73240, 11, 3 males, 30.9-63.7 mm SL, 8 females, 49.8-78.6 mm SL, (7, 61.8-78.6 mm SL), Camanducaia, road Camanducaia to Monte Verde, about 18 km of Camanducaia, sítio São Geraldo, córrego Casca d'Antas, 22°50'2.00"S 46°7'43.99"W, 05 Aug 2001, P. Gerhard & F. C. T. Lima. MZUSP 73243, 11, 5 males, 41.0-93.3 mm SL, 1 female, 64.4 mm SL, 5 unsexed, 23.3-39.2 mm SL, (2, 65.5-93.3 mm SL), Camanducaia, rodovia Camanducaia to Monte Verde, about 12 km of Camanducaia, bairro Quilombo. NUP 6586, 1 female, 72.7 mm SL, Camanducaia, road dos Peregrinos, córrego Casca d'Antas, 1352 m, 45°59'25"W 22°46'08"S, 24 Jul 2008, C. H. Zawadzki, B. F. Morales & V. S. Ferreira.
Diagnosis. Neoplecostomus jaguari can be distinguished from all species of Neoplecostomus by having a conspicuous sexual dimorphism: females with 30-38 teeth in the dentary and 30-39 in the premaxilla, and males with 13-18 and 14-20 teeth respectively (vs. number of teeth in the premaxilla and dentary equivalent in all other species, except for N. langeanii in the premaxilla); females with larger MW, 18.5-21.6% HL, males 11.8-15.7% HL (vs. females and males with MW/HL equivalent in the other species); females with teeth longer and thin, males with teeth shorter and robust (vs. teeth similar in males and females in all other species, except for N. langeanii in the premaxilla). Furthermore the new species is distinguished from all other species from the upper rio Paraná basin by having a lateronasal plate either forming the lateral border of the nostril or posteriorly displaced (vs. lateronasal plate absent). Finally, other useful character includes four to six plates between the frontal and prefrontal, here called posteronasal plates (vs. one to three in all other species).
Description. Counts and measurements in Table 1. Body relatively short and depressed. Greatest width at anterior portion of cleithrum, narrowing posteriorly to caudal peduncle. Dorsal body profile gently convex, elevating from tip of snout to dorsal-fin origin, then descending to first caudal-fin procurrent ray. Greatest body depth at dorsal-fin origin. Trunk and caudal peduncle dorsally rounded in cross-section; body ventrally flattened to anal-fin origin, flattened or slightly rounded to caudal fin.
Dorsal body surface completely covered by plates, except for naked area around dorsal-fin base. Snout tip without plates and odontodes. Head naked ventrally except by canal plate bearing odontodes directed mesially in front of gill openings. Head wide and moderately depressed. Head and snout slightly obtuse in specimens smaller than 50 mm SL in dorsal view and slightly rounded in specimens larger than 50 mm SL. Abdomen with conspicuous small platelets between pectoral and pelvic-fin insertions, forming thoracic shield surrounded by naked areas.
Mature males without hypertrophied odontodes and swollen skin along lateral margins of snout and dorsal anterior portion of head. Interorbital space slightly concave in frontal view. Eye moderately small and dorsolaterally placed. Lips well developed and rounded; lower lip almost reaching pectoral girdle and covered by papillae, wider and transversely flattened anteriorlly forming two or three irregular and conspicuous rows, just posterior to dentary teeth. Maxillary barbel short and almost entirely joined to lower lip, with free tip. Teeth long and thin in females and short and robust in males (Fig. 3), bicuspid, mesial cusp longer than lateral; premaxilla with 30-39 teeth in females and 14-20 in males; dentary with 30-38 teeth in females and 13-18 in males. Dentary rami forming an angle of approximately 110°-115°.
Dorsal-fin origin posterior to vertical passing through pelvic-fin origin, nuchal plate not covered by skin; dorsal-fin spinelet with same width or wider than dorsal-fin unbranched thick ray base; dorsal-fin locking mechanism absent. Dorsal-fin rays ii,7; distal margin straight, surpassing vertical through origin of pelvic fin. Adipose fin moderate to well developed, generally preceded by one azygous plate. Pectoral-fin rays i,6, unbranched thick ray depressed and inward curved, more curved in larger specimens and always shorter than longest branched ray; its distal margin nearly straight, when adpressed reaching first third of pelvic fin. Pelvic-fin rays i,5; distal margin nearly straight, when adpressed reaching or almost reaching anal-fin origin; pelvic-fin unbranched thick ray ventrally flattened; dermal flap on dorsal surface of pelvic-fin rays in males, more commonly along unbranched thick ray and first branched rays. Pectoral and pelvic-fin unbranched thick ray with odontodes on lateral and ventral portions. Anal fin rays i,5, posterior margin straight; unbranched thick ray with odontodes lateroventrally. Caudal fin rays i,14,i, bifurcate; lower lobe longer than upper.
Color in alcohol. Head and body light brown to dark in adults, and yellowish to dark brown in juveniles. Head, dorsum and flank with inconspicuous dark blotches, absent in some specimens. Dorsal, pectoral, pelvic, anal, and caudal fins with irregular series of dark bands on branched rays and usually conspicuous blotches in unbranched rays. Juveniles and some adults with five transverse dark bars: first and second inconspicuous, first through supraoccipital, second at dorsal-fin origin; third at dorsal-fin end; fourth at adipose fin; last at posterior portion of caudal peduncle. Head usually with two light, short and parallel lines anterior to nares, bordering naked area on tip of snout; sometimes another stripe between nares, generally absent in young individuals smaller than 50 mm SL. Orbital margin lighter, mainly on superior and posterior portions. Small, usually conspicuous, light spot on interorbital space.
Sexual dimorphism. Males with conspicuous urogenital papilla immediately posterior to anus (vs. absent in females); dermal flap on dorsal surface of the unbranched thick ray and branched pelvic-fin rays, less developed in branched rays (vs. absent in females); males with 14-20 teeth in the premaxilla and 13-18 in the dentary (vs. 30-39 and 30-38 in females); females with larger mandibullary width, 18.5-21.6% of the head length (vs. 11.8-15.7% in males), and teeth longer and thin in females (vs. shorter and robust in males) (Fig. 3).
Distribution. The species is known from the ribeirão do Forja and from córrego Casca d'Antas, both tributaries from the rio Jaguari, sub-basin from the rio Piracicaba-Capivari-Jundiaí, rio Tietê drainage, upper rio Paraná basin (Fig. 4).
Etymology. The specific epithet jaguari is a reference to the rio Jaguari, basin which ribeirão do Forja and the córrego Casca d'Antas belong. The epithet jaguari, from the Tupi Guarani, meaning river of jaguar ('onça').

Type localities of the Neoplecostomus species from the upper rio Paraná basin. Clover leaf - N. bandeirante; rhombus - N. botucatu; square - N. corumba; star - N. jaguari; circle - N. langeanii; pentagon - N. paranensis; cross - N. selenae; triangle - N. yapo.
Discussion
The wide geographic distribution of taxa such as Neoplecostomus paranensis was reported by Bizerril (1996) as possible cases of species complexes. Neoplecostomus paranensis was until very recently the only species occuring in a wide range in the upper rio Paraná basin; however, a review made by Zawadzki et al. (2008)Zawadzki, C. H., C. S. Pavanelli & F. Langeani. 2008. Neoplecostomus (Teleostei: Loricariidae) from the upper Rio Paraná basin, Brazil, with description of three new species. Zootaxa, 1757: 31-48.redescribed N. paranensis and described three new species; latter Roxo et al. (2012a)Roxo, F. F., C. Oliveira & C. H. Zawadzki. 2012a. Three new species of Neoplecostomus (Teleostei: Siluriformes: Loricariidae) from the Upper Rio Paraná basin of southeastern Brazil. Zootaxa, 3233: 1-21. described three additional species. Those two papers increased by almost 100% the total number of the species in the genus, also suggesting that its diversity may be greater than previously observed.
Morphological character sets which to date proved effective to differentiate species relate to body plates (e.g., N. ribeirensis having a divided canal plate and the spinelet absent or narrower than the dorsal-fin unbranched thick ray, and N. bandeirante having the first plates in the mid-ventral series shorter than the naked area between them), and sexual dimorphism as proposed by Zawadzki et al.(2008)Zawadzki, C. H., C. S. Pavanelli & F. Langeani. 2008. Neoplecostomus (Teleostei: Loricariidae) from the upper Rio Paraná basin, Brazil, with description of three new species. Zootaxa, 1757: 31-48. for N. selenae and N. yapo (adult males having large odontodes and swelling of skin on the lateral margins of the snout). Sexual dimorphism was also informative to diagnose Neoplecostomus jaguari, as reported above for some morphological features related to the number and form of teeth and mandibulary width, that until now were never referred to any other species of Neoplecostomus or even Neoplecostominae. Related to this, our analysis have shown that some of these dimorphims also occur in N. langeanii, the putative most related species to N. jaguari; these dimorphisms, however, were not reported in the original description of that species by Roxo et al. (2012a)Roxo, F. F., C. Oliveira & C. H. Zawadzki. 2012a. Three new species of Neoplecostomus (Teleostei: Siluriformes: Loricariidae) from the Upper Rio Paraná basin of southeastern Brazil. Zootaxa, 3233: 1-21.. These features such as the dimorphic ones, as already stated by Roxo et al. (2012a)Roxo, F. F., C. Oliveira & C. H. Zawadzki. 2012a. Three new species of Neoplecostomus (Teleostei: Siluriformes: Loricariidae) from the Upper Rio Paraná basin of southeastern Brazil. Zootaxa, 3233: 1-21., are more apparent or only present in mature specimens, and consequently samples lacking such specimens, relatively common in ichthyological collections, make identification of species difficult.
The upper rio Paraná basin is the second largest basin in the South America, the largest one in the Brazilian Cristaline Shield and, because of that, harbors a larger number of Neoplecostomus species when compared to other basins in the shield. We can, however, suppose a similar situation in those other basins, which also drain large geographic extensions in the area; consequently, material from such other basins is currently being analyzed in more detail in order to reveal putative undiscovered diversity.
Key to species of Neoplecostomus
1a. Enlarged, fleshy folds between dentaries in all specimens, more evident in mature males N. doceensis
1b. Absence of the enlarged fleshy folds............................... 2
2a. Lateronasal plate absent .................................................. 3
2b. Lateronasal plate present, between nare and infraorbital 2 or posteriorly 10
3a. Adipose fin absent or poorly developed ........................ 4
3b. Adipose fin present and well developed ........................ 5
4a. Conspicuous dark spots all over the body; head depth 55.6-63.9% of head length N. botucatu
4b. Conspicuous dark spots all over the body absent; head depth 44.4-55.4% of head length N. paranensis
5a. Moderate keels along each plate of lateral series, more evident in mature males; first plates in the mid-ventral series shorter than the area between them................................................................. N. bandeirante
5b. Characters above absent .................................................. 6
6a. Mature males with enlarged odontodes and distinct swollen skin along lateral margins of snout 7
6b. Mature males without enlarged odontodes and distinct swollen skin along lateral margins of snout 8
7a. Ridge of swollen skin and enlarged odontodes in front of eyes in mature males; 17-24 teeth in premaxilla and 15-24 teeth in dentary ................................................................................ N. selenae
7b. Ridge of swollen skin and enlarged odontodes in front of eyes in mature males absent; 10-15 teeth in premaxilla and 8-12 teeth in dentary ..................................................................... N. yapo
8a. Azygous preadipose plate present; cleithral width 19-23% of standard length; 5-13 teeth in dentary N. microps
8b. Azygous preadipose plate absent; cleithral width 25-30% of standard length; 10-39 teeth in dentary 9
9a. Orbital diameter 8.3-11.4% of head length; 7-9 plates between the adipose fin and caudal fin N. langeanii
9b. Orbital diameter 12.2-13.0% of head length ; 4-6 plates between the adipose fin and caudal fin N. corumba
10a. Spinelet absent or weakly developed, usually narrower than dorsal-fin unbranched thick ray; canal plate divided in three smaller plates .............................................................. N. ribeirensis
10b. Spinelet present and always wider than dorsal-fin unbranched thick ray; canal plate simple, not divided 11
11a. Lateral-line plates 34-43; 10-13 plates between the adipose fin and caudal fin .. N. granosus
11b. Lateral-line plates 27-33; 4-9 plates between the adipose fin and caudal fin 12
12a. Head length 26.2-26.8% of SL; head width 96.7-97.0% of head length N. variipictus
12b. Head length 28.0-34.5 of SL; head width 77.0-92.0% of head length 13
13a. Thoracic length 10.0-13.0% of SL ... N. franciscoensis
13b. Thoracic length 13.2-17.6% of SL ............................. 14
14a. Premaxillary and dentary teeth more numerous in females than males, respectively 30-39 (vs. 14-20) and 30-38 (vs. 13-18); cleithral width 25.2-28.7% of SL; orbital diameter 9.5-11.4% of length of the head N. jaguari
14b. Premaxillary and dentary teeth number equivalent for males and females, respectively 15-35 and 19-38; cleithral width 19-21% of SL; orbital diameter 6-9% of head length N. espiritosantensis
Comparative material examined. All from Brazil. Neoplecostomus bandeirante: LBP 2861, 8 paratypes, 75.0-80.5 mm SL, São Paulo State, Salesópolis, rio Tietê basin, rio Paraitinguinha. DZSJRP 14881, 2 paratypes, 93.8-97.2 mm SL, same data as LBP 2861; Neoplecostomus botucatu: LBP 7525, 20 paratypes, 76.2-76.4 mm SL, 1 c&s, 76.5 mm SL, São Paulo State, Botucatu, rio Paranapanema basin, rio Pardo drainage, córrego Águas de Madalena; DZSJRP 14879, 6 paratypes, 55.8-80.4 mm SL, same data as LBP 7525. Neoplecostomus corumba: All from Goiás State, rio Paranaíba drainage, rio Corumbá basin . DZSJRP 6193, 5, 44.0-73.8 mm SL, Corumbá, córrego Gameleira. DZSJRP 6713, holotype, 78.3 mm SL, same data as DZSJRP 6193; DZSJRP 7003, 19, 38.2-80.6 mm SL, 1 c&s, 58.1 mm SL, Bela Vista de Goiás, córrego of the Capoeira Grande. Neoplecostomus espiritosantensis: CIUFES 130722, paratype, 94.7 mm SL, Espírito Santo State, Domingos Martins, rio Jucu drainage; MCP 27340, 3, 53.1-66.5 mm SL, 1 c&s, 56.73 mm SL, same data as CIUFES 130722. Neoplecostomus franciscoensis: All from Minas Gerais State, rio São Francisco basin. DZSJRP 11458, 5, 41.7-65.1 mm SL, São Roque de Minas, córrego Grande; DZSJRP 11464, 3, 46.1-79.2 mm SL, 1 c&s, 73.89 mm SL, same data as DZSJRP 11458; MZUSP 37149, 40 paratypes, 26.5-74.9 mm SL, 1 c&s (unmeasured, dissected), Nova Lima, rio das Velhas basin, stream tributary to córrego Mutuca; DZSJRP 11451, 1, 40.0 mm SL, São Roque de Minas, around the Parque Nacional da Serra da Canastra, córrego Grande. Neoplecostomus langeanii: DZSJRP 14049, 15, 44.9-75.2 mm SL, 1 c&s, 69.15 mm SL, Minas Gerais State, Muzambinho, rio Grande basin, stream tributary to rio Muzambinho; DZSJRP 14048, 5, 41.2-69.9 mm SL, same data as DZSJRP 14049. DZSJRP 14882, 6 paratypes, 44.8-63.9 mm SL, same data as DZSJRP 14049; DZSJRP 14880, 2 paratypes, 69.0-70.6 mm SL, same data as DZSJRP 14049. LBP 5947, 9 of 35 paratypes, 46.6-55.8 mm SL, same data as DZSJRP 14049. Neoplecostomus microps: All from São Paulo State, rio Paraíba do Sul basin. DZSJRP 2767, 1 c&s, 56.0 mm SL, Campos da Cunha, rio Jucuí-Mirim drainage, riacho Jardim do Paraíso; DZSJRP 2768, 1 c&s (unmeasured, dissected); DZSJRP 4267, 9, 31.8-53.0 mm SL, São Luís do Paraitinga, ribeirão do Chapéu drainage, riacho Cachoeirinha; DZSJRP 2144, 17, 37.7-83.5 mm SL, 1 c&s, 62.78 mm SL, Cunha, stream tributary to rio Paraitinga; DZSJRP 13908, 23, 41.0-81.6 mm SL, Silveiras-Cunha, stream in the neighborhood Colinas. Neoplecostomus paranensis: DZSJRP 12320, 9, 33.1-69.6 mm SL, São Paulo State, São João da Boa Vista, rio Grande basin, córrego da Cachoeira; DZSJRP 8029, 35, 21.2-68.8 mm SL, São Paulo State, Analândia, upper rio Corumbataí basin, córrego Santa Terezinha; MZUSP 35822-35824, 3 paratypes, 68.4-92.7 mm SL, São Paulo State, Cajuru, rio Pardo basin, rio Cubatão; DZSJRP 8581, 2 of 3, 56.4-76.1 mm SL, Minas Gerais State, Fortaleza de Minas, rio Grande basin, stream 4 km from MG-050/BR-265, between São Sebastião do Paraíso and Passos; DZSJRP 8590, 10, 40.0-61.7 mm SL, 1 c&s, 51.0 mm SL, same data as DZSJRP 8581; DZSJRP 8608, 1, 47.2 mm SL, Minas Gerais State, Madre de Deus de Minas, rio Grande basin, first stream after rio Grande, MG-338. Neoplecostomus ribeirensis: All from São Paulo State, Ribeira de Iguape basin, DZSJRP 13622, 8, 38.4-70.0 mm SL, Apiaí, Barra do Chapéu, unnamed stream; DZSJRP 13696, 2, 62.5-74.6 mm SL, 1 c&s, 62.5 mm SL, Ibiúna, rio Itaguapeva; MZUSP 35432, 14 paratypes, 40.0-89.8 mm SL, 1 c&s (unmeasured, dissected), Miracatu, km 7 of the road Bigua-Iguape, stream tributary to rio Bananal; MZUSP 62007, 3, 54.0-71.9 mm SL, Iporanga, rio Iporanga, on the trail of the Mina do Espírito Santo; MZUSP 78440, 9, 50.0-82.2 mm SL, Ibiúna, Fazenda Vargedo, outside of the Parque Estadual Jurupará, rio Juquiá drainage, córrego Vergedo, cachoeira da Fumaça; MZUSP 51965, 3, 55.3-80.9 mm SL, Iporanga, neighborhood of the Serra, rio Betari basin. Neoplecostomus selenae: DZSJRP 7449, 4 paratypes, 56.5-95.8 mm SL, 1 c&s, 56.3, São Paulo State, Ribeirão Grande, rio Paranapanema basin, ribeirão das Bateias, upstream from the bridge at Ribeirão Grande to Intervales road; NUP 3572, 5 paratypes, 48.0-84.8 mm SL, same data as DZSJRP 7449; MZUSP 51889, holotype, 101.7 mm SL, same data as DZSJRP 7449. Neoplecostomus variipictus: MNRJ 12806, holotype, 90.2 mm SL, Rio de Janeiro State, Friburgo, rio Paraíba do Sul basin, rio Bengala drainage, rio Santo Antônio. Neoplecostomus yapo: All from rio Paranapanema basin, DZSJRP 6714, holotype, 97.4 mm SL, Paraná State, Tibagi, rio Tibagi basin, rio Yapó drainage, riacho Fortaleza; DZSJRP 6194, 6 paratypes, 69.4-105.3 mm SL, 1 c&s, 69.9 mm SL, same data as DZSJRP 6714. NUP 2609, 15 paratypes, 48.5-109.7 mm SL, same data as DZSJRP 6714; NUP 3569, 6 paratypes, 68.4-106.4 mm SL, same data as DZSJRP 6714; NUP 5685, 2 of 5, 37.0-60.7 mm SL, Paraná State, Imbaú, rio Tibagi basin, rio Charqueado; NUP 5694, 2 of 7, 100.8-102.8 mm SL, Paraná State, Ponta Grossa, rio Tibagi basin, rio Verde; DZSJRP 13651, 10, 49.8-80.2 mm SL, 1 c&s, 65.1 mm SL, São Paulo State, Piedade, rio Paranapanema basin, rio Turvo.
Acknowledgments
We are grateful to Cláudio Zawadzki (NUP), Edson Pereira (MCP-PUCRS), Fernanda Martins and Fernando Carvalho (DZSJRP), and Márcio Chiachio for suggestions; Carla S. Pavanelli and Cláudio H. Zawadzki (NUP), Carlos A. Lucena and Roberto E. dos Reis (MCP-PUCRS), Cláudio Oliveira (LBP), Flávio A. Bockmann and Ricardo M. C. e Castro (LIRP), and José L. de Figueiredo and Osvaldo T. Oyakawa (MZUSP), for the loan of specimens. This research was supported by the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES to BNA), Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP, 2012/23224-9 to FL) and Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq, 305.940/2011-0 to FL).
Literature Cited
- Armbruster, J. W. 2004. Phylogenetic relationships of the suckermouth armored catfishes (Loricariidae) with emphasis on the Hypostominae and the Ancistrinae. Zoological Journal of the Linnean Society, 141: 1-80.
- Bizerril, C. R. S. F. 1995. Description of a new species of Neoplecostomus (Loricariidae: Neoplecostominae), with a synopsis of the taxonomical composition of the Loricariidae in eastern Brazil. Arquivos de Biologia e Tecnologia, 38: 693-704.
- Langeani, F. 1990. Revisão do gênero Neoplecostomus Eigenmann & Eigenmann, 1888, com a descrição de quatro novas espécies do sudeste brasileiro (Ostariophysi, Siluriformes, Loricariidae). Comunicações do Museu de Ciências, PUCRS, série Zoologia, 3: 3-31.
- Lucena, A. L., M. E. Renesto, C. Oliveira, R. P. Mateus & C. H. Zawadzki. 2012. Genetic differentiation among ten populations of the genus Neoplecostomus (Teleostei: Loricariidae) from the upper Paraná River basin. Biochemical Systematics and Ecology, 44: 325-332.
- Reis, R. E., S. O. Kullander & C. J. Ferraris Jr. (Orgs.). 2003. Check List of the Freshwater Fishes of South and Central America. Porto Alegre, Edipucrs.
- Roxo, F. F., C. Oliveira & C. H. Zawadzki. 2012a. Three new species of Neoplecostomus (Teleostei: Siluriformes: Loricariidae) from the Upper Rio Paraná basin of southeastern Brazil. Zootaxa, 3233: 1-21.
- Roxo, F. F., C. H. Zawadzki, G. J. da Costa Silva., F. Foresti & C. Oliveira. 2012b. Molecular systematic of the armored neotropical catfish subfamiliy Neoplecostominae (Siluriformes: Loricariidae). Zootaxa, 3390: 33-42.
- Roxo, F. F., C. H. Zawadzki, M. A. Alexandrou, G. J. da Costa Silva, M. C. Chiachio, F. Foresti & C. Oliveira. 2012c. Evolutionary and biogeographic history of the subfamily Neoplecostominae (Siluriformes: Loricariidae). Ecology and Evolution, 10: 1-12.
- Roxo, F. F., G. S. C. Silva, C. H. Zawadzki & C. Oliveira. 2014. Neoplecostomus doceensis: a new loricariid species (Teleostei, Siluriformes) from the rio Doce basin and comments about its putative origin. Zookeys, 440: 115-127.
- Schaefer, S. A. 1997. The Neotropical cascudinhos: Systematics and biogeography of the Otocinclus catfishes (Siluriformes: Loricariidae). Proceedings of the Academy of Natural Sciences of Philadelphia, 148: 1-120.
- Taylor, W. R. & G. C. Van Dyke. 1985. Revised procedures for staining and clearing small fishes and other vertebrates for bone and cartilage study. Cybium, 9: 107-119.
- Zawadzki, C. H., A. L. Alves, E. Renesto & C. Oliveira. 2004. Biochemical evidence of a possible new species of Neoplecostomus (Teleostei: Loricariidae) from the upper Rio Paraná basin, Brazil. Biochemical Systematics and Ecology, 32: 573-582.
- Zawadzki, C. H., C. S. Pavanelli & F. Langeani. 2008. Neoplecostomus (Teleostei: Loricariidae) from the upper Rio Paraná basin, Brazil, with description of three new species. Zootaxa, 1757: 31-48.
Publication Dates
-
Publication in this collection
21 Nov 2014 -
Date of issue
Oct-Dec 2014
History
-
Received
30 Oct 2013 -
Accepted
08 June 2014




