ABSTRACT
Growth is a fundamental biological process, driven by multiple endogenous (intra-individual) and exogenous (environmental) factors that maintain individual fitness and population stability. The current study aims to assess whether individual, spatial (headwaters and floodplains) and inter-sex variation occurs in the growth of Piaractus mesopotamicus in the Cuiabá River basin. Samples were collected monthly from July 2006 to July 2007, at two areas in the Cuiabá River basin (headwaters and floodplain). Three growth models (individuals; individuals and sex factors; individuals and areas factors) were developed and compared the fish growth parameters using Akaike information criterion (AIC). The best fit to the length-at-age data was obtained by a model that considered individual variation and sex. The theoretical maximum average length (L∞ ) was 64.99 cm for females, and 63.23 cm for males. Females showed a growth rate (k) of 0.230 yr-1and males of 0.196 yr-1. Thus, could be concluded that individual variability and sex were the main sources of variation in P. mesopotamicus somatic growth parameters.
Keywords:
Age; Growth models; Inter-sex variation; Otoliths; Von Bertalanffy
RESUMO
O crescimento é um processo biológico fundamental, impulsionado por múltiplos fatores endógenos (intra-individual) e exógenos (ambientais) que mantém o fitness individual e a estabilidade populacional. Este trabalho tem como objetivo avaliar se ocorre variação individual, espacial (região de cabeceira e planície de inundação) e entre sexo no crescimento de Piaractus mesopotamicus na bacia do rio Cuiabá. Amostragens foram realizadas mensalmente entre julho de 2006 a julho de 2007, em duas áreas da bacia do rio Cuiabá (região de cabeceira e planície de inundação). Três modelos de crescimento (indivíduos, indivíduos e sexo como fator, indivíduos e área como fator) foram elaborados e comparados os parâmetros de crescimentos dos peixes com base no critério de Informação de Akaike (AIC). O melhor ajuste aos dados de comprimento na idade data foi obtido pelo modelo que considerou a variação individual e o sexo. O comprimento médio máximo teórico (L∞ ) das fêmeas foi 64,99 cm e para os machos 63,23 cm. O coeficiente de crescimento (k) médio foi de 0,230 yr-1 e 0,196 yr-1, respectivamente para fêmeas e machos. Portanto, pode-se concluir que variabilidade individual e sexo foram as principais origem de variação dos parâmetros de crescimento somático de P. mesopotamicus.
Palavras-chave:
Dimorfismo sexual; Idade; Modelos de crescimento; Otólitos; Von Bertalanffy
Introduction
From an ecological point of view, somatic growth is an important feature in the life history of individual fish because it is related to such vital characteristics as fertility, sexual maturity, survival and body size - factors that determine individual fitness and, ultimately, the persistence of a population (Stearns, 1976Stearns SC. Life-history tactics: a review of the ideas. Quart Rev Biol. 1976; 51(1):3-47.; Dmitriew, 2011Dmitriew CM. The evolution of growth trajectories: what limits growth rate? Biol Rev. 2011; 86(1):97-116.; Vincenzi et al., 2014Vincenzi S, Mangel M, Crivelli AJ, Munch S, Skaug HJ. Determining individual variation in growth and its implication for life-history and population processes using the empirical Bayes Method. PLoS Computational Biology. 2014; 10(9):1-16.). Fish retain growth records in various calcified structures, such as scales, otoliths, and vertebrae. These structures have an identifiable pattern of opaque and translucent zones. These comprise a sequential record of different frequencies of deposition which reflects the pattern of growth. Among these above-mentioned structures, the otolith is considered the most appropriate structure for analysis due to lack of absorption of growth increments (Green et al., 2009Vigliola L, Meekan MG. The back-calculation of fish growth from otoliths. In: Green BS, Mapstone BD, Carlos G, Begg GA, editors. Tropical Fish Otoliths: Information for Assessment, Management and Ecology. Reviews: Methods and Technologies in Fish Biology and Fisheries. New York: Springer ; 2009. p.174-211.). Factors affecting growth rate and hence annular deposition within otoliths include genetic variability, sexual dimorphism, population density, inter-annual variability, and seasonal and spatial variation in resource availability (Henryon et al., 2002Henryon M, Jokumsen A, Berg P, Lund I, Pedersen PB, Olesen NJ, Slierendrecht WJ. Genetic variation for growth rate, feed conversion efficiency, and disease resistance exists within a farmed population of rainbow trout. Aquaculture. 2002; 209(1-4):59-76.; Rypel, 2007Rypel AL. Sexual dimorphism in growth of freshwater drum. Southeastern Naturalist. 2007; 6(2):333-42.; Arantes et al., 2010Arantes CC, Castello L, Stewart DJ, Cetra M, Queiroz HL. Population density, growth and reproduction of arapaima in an Amazonian river-floodplain. Ecol Freshw Fish. 2010; 19(3):455-65.; Williams et al., 2012Williams AJ, Farley JH, Hoyle SD, Davies CR, Nicol SJ. Spatial and sex-specific variation in growth of albacore tuna (Thunnus alalunga) across the South Pacific Ocean. Plos One. 2012; 7(6):1-10.; Vincenzi et al., 2014Vincenzi S, Mangel M, Crivelli AJ, Munch S, Skaug HJ. Determining individual variation in growth and its implication for life-history and population processes using the empirical Bayes Method. PLoS Computational Biology. 2014; 10(9):1-16.). However, these factors do not act alone, as there is a complex interaction between them (Dieterman et al., 2012Dieterman DJ, Hoxmeier RJH, Staples DF. Factors influencing growth of individual brown trout in three streams of the upper Midwestern United States. Ecol Freshw Fish . 2012; 21(3):483-93.). Growth is simultaneously due to individual response to variability (the individual genotype), and behavioural differences in reproduction, habitat selection and foraging-based migration between sexes (DeAngelis, Grimm, 2014DeAngelis DL, Grimm V. Individual-based models in ecology after four decades. F1000Prime Reports. 2014; 6(39):1-6.). Additionally, the environment in which an individual occurs can generate variations in growth parameters due to the nature and extent of available resources (Siangas et al., 2012Siangas EL, Pouilly M, Vallejos A, Pérez T, Rejas D. Effect of water quality on growth of four fish species in the Iténez basin (Upper Madera, Amazon). Environ Biol Fish . 2012; 95(3):371-81.). Since growth is an attribute that responds to multiple factors, tools and data analysis approaches have been developed recently to estimate growth model parameters with the least possible bias. Among the approaches used are comparisons of different growth models adjusted for the same length-at-age data set (Katsanevakis, 2006Katsanevakis S. Modelling fish growth: model selection, multi-model inference and model selection uncertainty. Fish Res . 2006; 81(2):229-35.). To fit each growth model, the parameters are estimated by adding different factors, as individual, local and sex and then comparisons between the models are performed in order to identify which model best explains the variation in the fish growth parameters (Burnham, Anderson, 2002Burnham KP, Anderson DR. Model selection and inference: a practical information - theoretic approach. 2nd ed. New York: Springer; 2002.; Vigliola, Meekan, 2009Vigliola L, Meekan MG. The back-calculation of fish growth from otoliths. In: Green BS, Mapstone BD, Carlos G, Begg GA, editors. Tropical Fish Otoliths: Information for Assessment, Management and Ecology. Reviews: Methods and Technologies in Fish Biology and Fisheries. New York: Springer ; 2009. p.174-211.). Model selection is a relatively new paradigm in biological sciences and has several advantages over the classical approaches (Katsanevakis, 2006Katsanevakis S. Modelling fish growth: model selection, multi-model inference and model selection uncertainty. Fish Res . 2006; 81(2):229-35.). Model selection methods try to rank models in the candidate set relative to each other; whether any of the models is actually “good” depends primarily on the quality of the data and the science and a priori thinking that went into the modeling (Burnham, Anderson 2002Burnham KP, Anderson DR. Model selection and inference: a practical information - theoretic approach. 2nd ed. New York: Springer; 2002.).
A robust growth model must take into account individual variation at a variety of levels, since any observed difference in somatic growth in populations can result from the interaction of the genotype with exogenous factors (Weisberg et al., 2010Weisberg S, Spangler G, Richmond LS. Mixed effects models for fish growth. Can J Fish Aquat Sci . 2010; 67(2):269-77.). Each individual is born with a particular genetic make-up that to some extent controls its growth profile, but physical and biological processes modify the on-going growth rate of an individual (Shelton et al., 2013Shelton AO, Satterthwaite WH, Beakes MP, Munch SB, Sogard SM, Mangel M. Separating intrinsic and environmental contributions to growth and their population consequences. Am Nat. 2013; 181(6):799-814.; Vincenzi et al., 2014Vincenzi S, Mangel M, Crivelli AJ, Munch S, Skaug HJ. Determining individual variation in growth and its implication for life-history and population processes using the empirical Bayes Method. PLoS Computational Biology. 2014; 10(9):1-16.). Traditional growth models (i.e., a single observation of length-at-age data per individual) clearly ignore such individual variability because they do not consider the growth trajectory of each individual in the population, and estimates of somatic growth parameters are usually described by “average” population parameters. Recent studies emphasize the importance of including intra-population variation in growth models so as to reduce bias in parameter estimation (Pilling et al., 2002Pilling GM, Kirkwood GP, Walker SG. An improved method for estimating individual growth variability in fish, and the correlation between von Bertalanffy growth parameters. Can J Fish Aquat Sci . 2002; 59(3):424-32.; Dieterman et al., 2012Dieterman DJ, Hoxmeier RJH, Staples DF. Factors influencing growth of individual brown trout in three streams of the upper Midwestern United States. Ecol Freshw Fish . 2012; 21(3):483-93.; Shelton et al., 2013Shelton AO, Satterthwaite WH, Beakes MP, Munch SB, Sogard SM, Mangel M. Separating intrinsic and environmental contributions to growth and their population consequences. Am Nat. 2013; 181(6):799-814.; Vincenzi et al., 2014Vincenzi S, Mangel M, Crivelli AJ, Munch S, Skaug HJ. Determining individual variation in growth and its implication for life-history and population processes using the empirical Bayes Method. PLoS Computational Biology. 2014; 10(9):1-16.). This is important because individuals in a population do not have similar growth parameters. The main problem in the use of models that assume the absence of individual variability is the bias in the life expectancy of the species. In the case of selective fishing size, longevity is a attribute dependent on individual body size, and those individuals that grow most rapidly tend to be captured at a younger age than those growing more slowly. This could occurs both in commercial and recreational fishing when minimal capture size was established as a fisheries management strategy. In addition it is important to consider the sex of the individual, since the occurrence of sexual dimorphism is common in both marine and freshwater fish. Consideration of the influence of differences in behaviour and differential investment in reproduction and growth between the sexes has permitted a better understanding of autoecology and dynamics of fish stocks (Araújo, Haimovici, 2000Araújo JN, Haimovici M. Determinação de idades e crescimento do linguado branco Paralichthys patagonicus (Jordan, 1889) no sul do Brasil. Rev Bras Oceanogr. 2000; 48(1):61-70.). Studies investigating the role of sex as a factor in growth models and methods for back-calculating size have shown that much of the variation between individuals is related to sex (Kielbassa et al., 2011Kielbassa J, Charles S, Delignette-Muller ML. The importance of incorporating age and sex when backcalculating length in bullhead Cottus gobio. J Fish Biol . 2011; 78(5):1492-507.; Ambrósio et al., 2014Ambrósio AM, Balbi TJ, Francisco TM, Gomes LC, Zuliani MS, Okada EK. Aging and growth parameter from the Piaractus mesopotamicus (pacu) from the Cuiabá river, Mato Grosso, Brazil. Iheringia Sér Zool. 2014; 104(4):413-17.).
In the current context of increasing fragmentation of aquatic habitats and overfishing of continental fish stocks in the Neotropics, a better understanding of the responses of exploited fish populations is necessary. Piaractus mesopotamicus (Holmberg, 1887) is such a species. Popularly known as pacu, it is a member of the family Serrasalmidae, subfamily Myleinae (Britski et al., 1999Britski HA, Silimon KZS, Lopes BS. Peixes do pantanal. Manual de identificação. Brasília: Embrapa -SPI; 1999.). A long distance migrant with wide geographic distribution in the La Plata Basin (Reis et al., 2003Reis RE, Kullander SO, Ferraris Júnior CJ. Check list of the freshwater fishes of South and Central America. Porto Alegre: EDIPUCRS; 2003.), the pacu is an important seed disperser in the Neotropics (Correa et al., 2015Correa SB, Araujo JK, Penha JMF, Cunha CN, Stevenson PR, Anderson JT. Overfishing disrupts an ancient mutualism between frugivorous fishes and plants in Neotropical wetlands. Biol Conserv. 2015; 191:159-67.). Growth rates for P. mesopotamicus developed in the Cuiabá River, Mato Grosso, and using scales, found variation between sex k=0.14 year-1 and L∞ 50 cm for females and k=0.18 year-1 and L∞ 59.23 cm for males. The maximum age at capture was eight years (Ambrósio et al., 2014Ambrósio AM, Balbi TJ, Francisco TM, Gomes LC, Zuliani MS, Okada EK. Aging and growth parameter from the Piaractus mesopotamicus (pacu) from the Cuiabá river, Mato Grosso, Brazil. Iheringia Sér Zool. 2014; 104(4):413-17.). However, stocks in the Cuiabá River system are overexploited (Vaz, 2001Vaz MM. Problemas no ajuste da curva de crescimento do pacu, Piaractus mesopotamicus (Holmberg, 1887) (Pisces: Characidae), e seu manejo no Pantanal Mato-Grossense. [PhD Thesis]. Jaboticabal, SP: UNESP; 2001.; Peixer et al., 2007Peixer J, Petrere Júnior M. Hook selectivity of the pacu Piaractus mesopotamicus (Holmberg, 1887) in the Pantanal, the state of Mato Grosso do Sul, Brazil. Braz J Biol . 2007; 67(2):339-45.; Peixer, Petrere, 2007Peixer J, Petrere Júnior M. Hook selectivity of the pacu Piaractus mesopotamicus (Holmberg, 1887) in the Pantanal, the state of Mato Grosso do Sul, Brazil. Braz J Biol . 2007; 67(2):339-45.). Piaractus mesopotamicus is the most frequently caught species in Mato Grosso do Sul, representing almost 1/3 of the total production of professional fishing between 1994 and 1999 and corresponding to 2,393.8 tons (30.2%) of the total catch (Catella, 2001Catella AC. A pesca no pantanal de Mato Grosso do Sul, Brasil: descrição, nível de exploração e manejo (1994-1999). [PhD Thesis]. Manaus, AM: Instituto Nacional de Pesquisa da Amazônia; 2001.). In the Cuiabá River in the State of Mato Grosso, the species represented 13.52 tons (8%) of the commercial catch in 2001 and 2002 in Cuiabá, occupying the fourth position in landings (Mateus et al., 2004Mateus LAF, Penha JMF, Petrere M. Fishing resources in the rio Cuiabá Basin, Pantanal do Mato Grosso, Brazil. Neotrop Ichthyol . 2004; 2(4):217-27.). Given the current status of continuing fishing pressure P. mesopotamicus stocks (Vaz, 2001Vaz MM. Problemas no ajuste da curva de crescimento do pacu, Piaractus mesopotamicus (Holmberg, 1887) (Pisces: Characidae), e seu manejo no Pantanal Mato-Grossense. [PhD Thesis]. Jaboticabal, SP: UNESP; 2001.), it is important to assess the sources of variation in somatic growth patterns. Therefore, the aim of this study was to evaluate whether there exists spatial (headwaters and floodplain) and between-sex variation in the growth of P. mesopotamicus individuals from the Cuiabá River basin.
Material and Methods
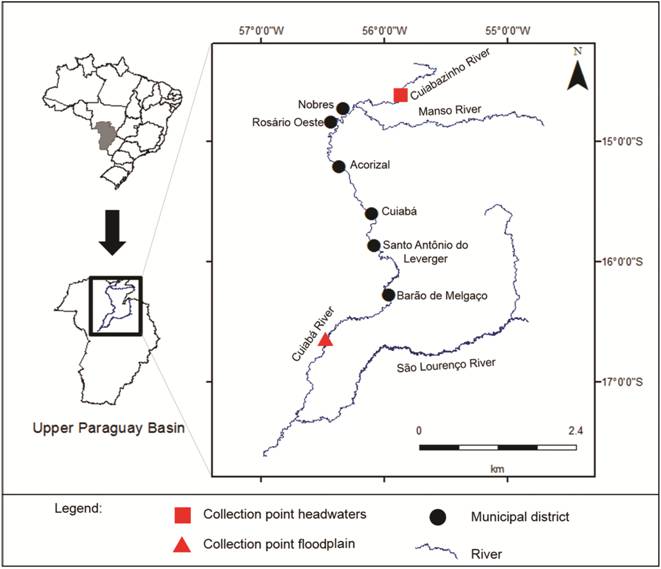
Study area. The Cuiabá River is a major tributary of the Paraguay River, and its basin covers an area of some 28,732 km2 is located entirely in the state of Mato Grosso, Brazil. The Cuiabá River can be divided into Upper Cuiabá- the headwaters region, with considerable variation in altitude, with many rapids and watercourses running through typical Cerrado vegetation; Mid-Cuiaba - the transition region between the headwaters region and floodplain; and the Lower Cuiabá- which comprises the gently sloping floodplain, where open savanna formations are dominant (Figueiredo, Salomão, 2009Figueiredo DM, Salomão FXT. Bacia do Rio Cuiabá: Uma abordagem socioambiental.Cuiabá: EdUFMT; 2009.). The Cuiabá River together with the São Lourenço, Piquiri, Taquari, Miranda and Negro Rivers, seasonally overflows and floods an area of approximately 150,000 km2, and is therefore one of the main formers of the Pantanal floodplain (Cavinatto, 1995Cavinatto V. Caracterização Hidrográfica do Estado de Mato Grosso. Cuiabá: Prodeagro/Seplan/Fema-MT. 1995.). Its headwaters are located in the Rosario Oeste city, state of Mato Grosso, on the slopes of the Serra Azul mountain range, and its main sources are the Cuiabá da Larga and Cuiabá do Bonito Rivers. The river formed by the confluence of these two rivers is called Cuiabazinho River, which in turn joins the Manso River, giving rise to Cuiabá River. The tropical sub-humid climate of the region is characterized by an average high temperature of 25 ◦C and a mean annual precipitation of about 1900 mm in the headwater regions of the mountain ranges, decreasing centripetally in the direction of the floodplain region about 1250 mm (Zeilhofer, Moura, 2009Zeilhofer P, Moura RM. Hydrological changes in the northern Pantanal caused by the Manso dam: Impact analysis and suggestions for mitigation. Ecol Engin. 2009; 35(1):105-17.). In the Cuiabá river basin, the alternation of rainy and dry seasons underpins a multi-annual variability, so that there are alternating multi-year cycles of rainy and dry years (Hamilton et al., 1996Hamilton SK, Sippel SJ, Melack JM. Inundation patterns in the Pantanal wetland of South America determined from passive microwave remote sensing. Arch Hydrobiol. 1996; 137(1):1-23.). Annually, the pattern of rainfall provides four hydrological periods: flood period (October to December); wet (January-March); ebb (April-June) and dry (July-September) (Da Silva, Esteves, 1995Da Silva CJ, Esteves FA. Dinâmica das características limnológicas das baías Porto de Fora e Acurizal (Pantanal de Mato Grosso) em função da variação do nível da água. Oecol Brasil. 1995; 1:47-60.).
Collections occurred at two sites within the basin: one in the headwaters (14°29’55.57”S and 55°45’25.59”W) in Cuiabazinho River and another in the floodplain (16°32’5.75”S and 56°23’58.43”W) in the Porto Cercado (Barão de Melgaço region) (Fig. 1). The distance between the collection points was approximately 390 km. Cuiabazinho River is narrower than the Cuiabá River, with a width varying from 40 to 70 m (Ziober et al., 2012Ziober SR, Bialetzki A, Mateus LAF. Effect of abiotic variables on fish eggs and larvae distribution in headwaters of Cuiabá River, Mato Grosso State , Brazil. Neotrop Ichthyol . 2012; 10(1):123-32.). On the other hand, in Porto Cercado the Cuiaba river channel is 170 m wide and the mean annual discharge at this station is 280 m3 s-1 (Fantin-Cruz et al., 2011Fantin-Cruz I, Pedrollo O, Castro NMR, Girard P, Zeilhofer P, Hamilton SK. Historical reconstruction of floodplain inundation in the Pantanal (Brazil) using neural networks. J Hydrol. 2011; 399(3-4):376-84.).

Sampling points in the headwater (square) and floodplain (triangle) areas of the Cuiabá River basin.
Fish collection and measurements. Monthly, from July 2006 to July 2007, individual Piaractus mesopotamicus were collected, using gill nets with mesh ranging from 17 to 20 cm between opposite knots, gill nets with mesh 18-20 cm between opposite knots, hooks, fishing nets and longlines. For each specimen we recorded total-length (TL, cm) and were weight to the nearest 50 g. After which, a longitudinal incision was made along the abdominal surface of each specimen from the head to the urogenital opening, and the abdominal cavity inspected. Gonads were removed, and histological analysis was performed to identify the sex. For a detailed description of gonad processing the sex identification in P. mesopotamicus, see Costa, Mateus (2009Costa RMR, Mateus LAF. Reproductive biology of pacu Piaractus mesopotamicus (Holmberg, 1887) (Teleostei: Characidae) in the Cuiabá River Basin, Mato Grosso, Brazil. Neotrop Ichthyol. 2009; 7(3):447-58.).
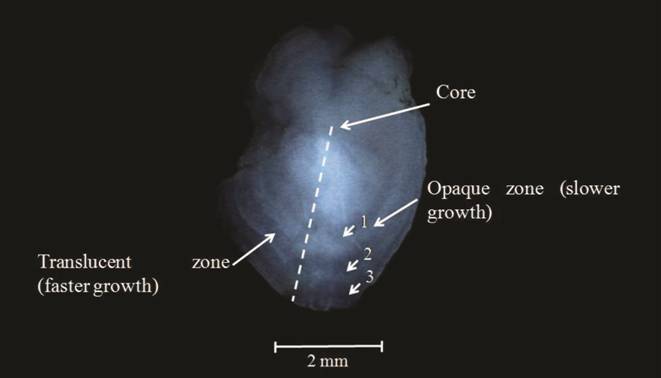
To access otoliths, the head was sectioned with a midline incision in the longitudinal direction, and the otoliths removed with fine tip forceps, one from each side. They were then cleaned with water, dried on paper towels, identified and labelled for storage in dry microtubes. The selection of the pair of otoliths most appropriate for age-estimation and the ageing protocol, is specific to the species and depends on the quality and resolution of the macro- and micro-structures (Secor et al., 1992Secor DH, Dean JM, Laban EH. Otolith removal and preparation for microstructural examination. In: Stevenson DK, Campana SE, editors. Otolith Microstructure Examination and Analysis. Ottawa: Department of Fisheries and Oceans; 1992. p.19-57.). In the case of P. mesopotamicus, the lapillus otolith was chosen for growth analysis because this otolith type was the largest, had the most sharply defined calcified deposits and could be managed easily, all of which facilitated growth ring identification. In the laboratory, whole otoliths were polished using waterproof sandpaper (grit grade 600) until the nucleus and macrostructures of the otholith were visible. Then, to allow the growth increments to be seen greater clarity, polished otoliths were placed under 70% alcohol on a black backed petri-dish and examined under incident light and magnification 10x with a stereomicroscope. Photographs were taken using a camera attachment. Photographs were taken under incident light to make the opaque zones appear light and the translucent zones dark (Secor et al., 1992Secor DH, Dean JM, Laban EH. Otolith removal and preparation for microstructural examination. In: Stevenson DK, Campana SE, editors. Otolith Microstructure Examination and Analysis. Ottawa: Department of Fisheries and Oceans; 1992. p.19-57.). Growth increments were counted and the following measures taken with use of the Leica program: otolith radius (distance from core to otolith edge) radius of each ring (distance from core to the external edge of the translucent area of each annulus) and edge (distance from the last annulus to the end of the otolith) (Fig. 2) (Lai et al., 1996Lai HL, Gallucci VF, Gunderson DR, Donnelly RF. Age determination in fisheries: methods and applications to stock assessment. In: Galucci VF, Saila SB, Gunderson DJ, Rothschild BJ, editors. Stock Assessment: Quantitative Methods and Applications for Small-Scale Fisheries. New York: Lewis Publishers - CRC Press; 1996. p.82-178.). Measurements were made in centimeters (cm), the orientation of measurements was the same for all analysed otoliths. Three independent assays of age ring number were made to attain accurate estimate of the fish age. The interpretation process is complex, involving decisions on interpretation where many factors can influence the judgement call of the reader. To avoid errors interpretations two readers was made by the same reader in intervals of the three weeks. The third reader was conducted by another reader after finalized the others readers. Each reading was completed in one month. Simultaneous to readers the width of the each rings were made, and all readings were made without auxiliary information or reference to any previous set of readings (Secor et al., 1992Secor DH, Dean JM, Laban EH. Otolith removal and preparation for microstructural examination. In: Stevenson DK, Campana SE, editors. Otolith Microstructure Examination and Analysis. Ottawa: Department of Fisheries and Oceans; 1992. p.19-57.; Campana, 2001Campana SE. Accuracy, precision and quality control in age determination, including a review of the use and abuse of age validation methods. J Fish Biol. 2001; 59(2):197-242.). The analysed otoliths were deposited in the fish collection of the Universidade Federal de Mato Grosso under the numbers CPUFMT4459 to CPUFMT4482.

The left otolith of a male Piaractus mesopotamicus that was collected in the headwaters of the Cuiabá River. Three annuli are marked (arrows), and the reading axis indicated (dashed line).
Ageing precision. To assess the reproducibility of age determinations, the three readings of each otolith from each fish was used to determine the coefficient of variation (CV) expressed as the ratio of the standard deviation over the mean, defined as:
Where CVj is the precision of the age estimate for the jth fish, Xij is the i th age determination of the j th fish, X j is the mean age estimate of the j th fish, and R is the number of times the age of each fish was measured (Campana, 2001Campana SE. Accuracy, precision and quality control in age determination, including a review of the use and abuse of age validation methods. J Fish Biol. 2001; 59(2):197-242.).
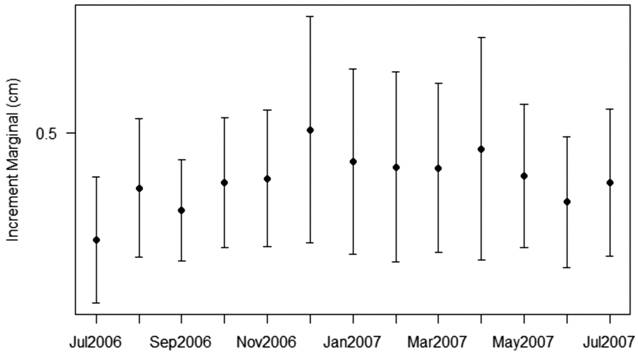
Periodicity of the deposition increments. The periodicity of ring deposition was analyzed by absolute marginal increment analysis (MI) (Lai et al., 1996Lai HL, Gallucci VF, Gunderson DR, Donnelly RF. Age determination in fisheries: methods and applications to stock assessment. In: Galucci VF, Saila SB, Gunderson DJ, Rothschild BJ, editors. Stock Assessment: Quantitative Methods and Applications for Small-Scale Fisheries. New York: Lewis Publishers - CRC Press; 1996. p.82-178.): MI=(rn )/rn-1), where rn =distance between otolith nucleus and the last band; and rn -1=distance between otolith nucleus and the penultimate band.
Relationship between body length and otolith radius. Back-calculation was used to reconstruct individual growth trajectories from otoliths. The approach involves using measurements made on otoliths to infer, or back-calculate, body length at ages prior to capture (Vigliola, Meekan, 2009Vigliola L, Meekan MG. The back-calculation of fish growth from otoliths. In: Green BS, Mapstone BD, Carlos G, Begg GA, editors. Tropical Fish Otoliths: Information for Assessment, Management and Ecology. Reviews: Methods and Technologies in Fish Biology and Fisheries. New York: Springer ; 2009. p.174-211.). To calculate body length from individuals at previous capture ages Linear Biological Intercept models were used (Campana, 1990Campana SE. How reliable are growth back-calculations based on Otoliths? Can J Fish Aquat Sci. 1990; 47:2219-37.). This method is applied when the relation between body length and otolith radius is linear. The equation used was: Li=Lcpt +(Ri-Rcpt)(Lcpt-Lop)/(Rcpt-Rop), where Ri and Liare the radius and size at agei; Rcpt are Lcpt are radius and body length in the catch; Rop and Lop are otolith radius and initial body length (usually at age zero). The value of Lop used was 0.9 cm (Beerli et al., 2004Beerli EL, Logato PVR, Freitas RTF. Alimentação e comportamento de larvas de pacu Piaractus mesopotamicus (Holmberg, 1887). Ciênc Agrotecnol. 2004; 28(1):149-55.). As no larvae were sampled in the study, and no information was found in the available literature on the average value of the otolith radius for P. mesopotamicus larvae, we assumed Rop was equal to 0.001 cm.
Somatic growth models. Once length-at-age data are obtained, growth curve parameters can be estimated for a species. For fish the von Bertalanffy model is one of those commonly used to adjust the growth curve: Lt =L∞ (1 - e -k(t -t0)), Where L t is length at age t, L∞ is the maximum or asymptotic length; k, is a growth rate constant that determines the curvature of the growth function; t 0, the nominal age at length zero, is a location term that shifts the curve to the right or left of the age axis. With length-at-age data a common problem when fitting the growth model is the lack of young and old fish in the samples. Young fish are often not present in the sample due to sampling difficulties (gear selectivity). The lack of old fish in the sample may be due to sampling a population where the mortality rate is so high that fish do not live long enough to approach the asymptote (Francis, 1988Francis RICC. Are growth parameters estimated from tagging and age-length data comparable? Can J Fish Aquat Sci . 1988; 45(6):936-42.). The absence of fish of these classes may restrict the ages to a narrow range that is functionally linear, which make it difficult to fit a nonlinear function to relatively linear data. Due to the resulting linearly problem in length-at-age data, the growth parameters were estimated using an expected-value reparameterization of von Bertalanffy growth function, as proposed by Ratkowsky (1986Ratkowsky DA. Statistical properties of alternative parameterizations of the von Bertalanffy growth curve. Can J Fish Aquat Sci . 1986; 43(4):742-47., 1990Ratkowsky DA. Handbook of Nonlinear Regression Models. New York: Marcel Dekker; 1990.; see also Schnute, Fournier, 1980Schnute J, Fournier D. A new approach to length-frequency analysis: growth structure. Can J Fish Aquat Sci . 1980; 37(9):1337-51.; Cerrato, 1991Cerrato RM. Analysis of nonlinearity effects in expected-value parametrizations of the von Bertalanffy equation. Can J Fish Aquat Sci . 1991; 48(11):2109-17.):
Where Y is length at age X, q=2(X - X1) / (X2 - X1) and the expected-value parameters l 1, l 2 and l 3 are the lengths corresponding to ages X 1 (1yr), X 2 (5 yr) and X3 (3 yr, the arithmetic mean of X 1 and X 2) (Ratkowsky, 1990Ratkowsky DA. Handbook of Nonlinear Regression Models. New York: Marcel Dekker; 1990.). The reparametrization of the von Bertalanffy growth curves are based on the use of the median size in each age class, which allows the resolution of problems with non-convergence caused by such linear length as that used for P. mesopotamicus age, since the use of length for estimation of the parameters produces data with an asymptote. Using the expected-value reparametrization provides parameters that have both direct biological meaning and more statistically favorable properties than the standard parameters of the von Bertalanffy growth function (Francis, 1988Francis RICC. Are growth parameters estimated from tagging and age-length data comparable? Can J Fish Aquat Sci . 1988; 45(6):936-42.; Welsford, Lyle, 2005Welsford DC, Lyle JM. Estimates of growth and comparisons of growth rates determined from length- and age-based models for populations of purple wrasse (Notolabrus fucicola). Fish Bull. 2005; 103(4):697-711.).
To assess growth Piaractus mesopotamicus parameters, we built three models: 1) Individual level, reflecting variation between individuals, 2) Individual level and sex effects on growth parameters, a model taking into account the variation in the growth parameters of individual (random effects) and sex (fixed effects), and 3) Individual level and site as fixed effects, reflecting variation among individuals in the growth trajectory due to site quality.
All three models have the general form: yi (X)=Y(X)+ εi , where yi is the back-calculated length of an individual at age X, and εi is a measurement-level error term, or residual, that reflects the deviations of back-calculated length yi from the length-at-age determined from the estimated growth curve, Y(X), for that individual. The models differ in the specification of the expected-value parameters lk (k=1, 2, 3).
In Model 1 (Individual level): lkp =lko + ςkp, where lkp is the value of parameter lk for individual p, lk0 is a basal value common to all individuals, and ςkp is an individual-level random effect. In Model 2 (Individual level and sex): lkp =lki + ςkp, where lkp is the value of parameter lk for individual p, lki is the sex-based fixed effect, and ςkp is an individual-level random effect. For Model 3 (Individual level and site): lkp =lkj + ςkp, where lkp is the value of parameter lk for individual p, lkj is the site-based fixed effects, and ςkp is an individual-level random effect.
Parameter estimation and model comparison. Estimation for the reparametrized growth model was based on numerical maximization of log-likelihood. Model comparisons and selection were based on Akaike’s Information Criterion (AIC) (Burnham, Anderson, 2002Burnham KP, Anderson DR. Model selection and inference: a practical information - theoretic approach. 2nd ed. New York: Springer; 2002.). In addition to the AIC value, where the lower values indicate the best-fit models, we used two other metrics (∆i values) to analyse the differences between models. These allow the acceptability of each model to be numerically evaluated (∆i < 2=strong support in the data; ∆i >=2 and < 7=little support in the data; ∆i > 10=without support in the data) and Akaike weight (Wi), which is the probability of a given model in the cases of resampling the available data (Burnham & Anderson 2002Burnham KP, Anderson DR. Model selection and inference: a practical information - theoretic approach. 2nd ed. New York: Springer; 2002.). Parameter estimates and associated confidence intervals for the von Bertalanffy growth function were obtained by back-transformation from the corresponding estimates in the reparametrized data (Francis, 1988Francis RICC. Are growth parameters estimated from tagging and age-length data comparable? Can J Fish Aquat Sci . 1988; 45(6):936-42.). Back-transformation of the parameters was made using the mvtnorm package (v. 1.0.5) (R Development Core Team, 2015R Development Core Team. R: A language and environment for statistical computing. [Computer software manual - Internet]. Vienna: R Foundation for Statistical Computing; 2014. Available from: http://www.R-project.org/
http://www.R-project.org/...
), where the mean value of the estimated parameters of each model and their respective covariance matrices were used in a solution procedure in R program. All growth analyses were carried out using the nlme package in the R environment (v. 2.14.1) (R Development Core Team, 2012R Development Core Team. R: A language and environment for statistical computing. [Computer software manual - Internet]. Vienna: R Foundation for Statistical Computing; 2014. Available from: http://www.R-project.org/
http://www.R-project.org/...
). Additionally, a two-way ANOVA was performed (Sokal, Rohlf, 1995Sokal RR, Rohlf FJ. Biometry: the principles of statistics in biological research. 3rd ed. New York: Freeman; 1995.) using body size as the response variable and sex and location as predictors in order to assess if the body size varied by sex and location. Finally, the relationship between growth parameters (L∞ , t
0, k) was used to test the effect of sex on somatic growth, as suggested by Rypel (2007Rypel AL. Sexual dimorphism in growth of freshwater drum. Southeastern Naturalist. 2007; 6(2):333-42.).
Results
A total of 154 specimens were collected from the floodplain region and 179 from the headwaters (a total sample of 333 individuals). Within the total catch, 166 individuals were female and 167 male. For females total length ranged from 29 cm to 73 cm, with an average of 46.47 cm (SD=8.74). Total length of males ranged from 34 cm to 58 cm, with an average of 41.95 cm (SD=4.22). Females had a greater average length than males. Weight varied between 0.70 and 10.2 kg.
Ageing precision. Whole Piaractus mesopotamicus otoliths were used in the growth study because increments could be easily seen. The coefficient of variation between the three readings was 10.29%, which indicated consistency between readings and the protocol adopted is therefore replicable for the species.
Periodicity of the deposition increments. The formation period of a growth ring was considered to be that for which MI had the lowest value. The marginal increment had the lowest average in July and September 2006, and in 2007 there was a decrease in May, June and July (Fig. 3). We conclude that the period of ring formation occurred during months of lowest regional rainfall; these occur at the end of the ebb and early dry. Therefore, P. mesopotamicus otolith structures are suitable for growth ring counts, and deposition of translucent and opaque zones correspond to 1 year intervals. Opaque zones were much wider than translucent ones, indicating a higher growth rate. The formation of the translucent zone coincided with a period of reduced growth and the opaque zone corresponded to faster growth (Fig. 2).

Monthly mean values and standard deviation of the marginal increment from otolith of the Piaractus mesopotamicus sampled from July 2006 to July 2007 in the Cuiabá River basin.
Males and females between 2 and 5 years of age were captured. No individuals with one age ring were captured. As only one 5 year old male (total length of 43 cm) was captured in the headwater region, this age class was not considered in the analysis. For both sexes 3 and 4 year old individuals were the most abundant in the samples (Tab. 1).
Sample size (N), minimum total length (TL min), maximum total length (TL max), mean total length for each age group and standard deviation (SD) and coefficient of variation (CV) of the mean total length observed for each age, for males and females of the Piaractus mesopotamicus sampled from July 2006 to July 2007 in the Cuiabá River basin.
Relationship between body length and otolith radius at capture. The relationship between otolith radius and fish body length was linear (r2=0.50 and p <0.001) (Fig. 4). Because the existence of proportionality between body size and otolith radius, the Biological Linear Intercept method was used to calculate the lengths of previously captured known-age individuals. To provide the most robust base for the study, the mean lengths of each sex were compared with the mean lengths by age and age group (Tab. 2), the back-calculated lengths in year-groups 2 and 3 were below the mean for both sexes compared to the length of the total catch.

Relationship between body length and otolith radius at capture of the Piaractus mesopotamicus from the Cuiabá River basin.
Comparison of the length at capture and back-calculated length by age groups for P. mesopotamicus sampled in the the Cuiabá River basin.
Parameter estimation and model comparison. The models providing best fit for P. mesopotamicus length-at-age data were those which considered the variation in somatic growth parameters via sex and locality. By rank, the best model was that which considered the individual growth trajectory (random factor) and the effect of sex (fixed factor) with somatic growth parameters (L∞, k and t 0), with a value of ∆i=0.0 and Wi=1 (Tab. 3). The second best model considered the spatial variation (Individual and site), i.e., the local effect as a fixed factor in growth parameters ∆i=17.0 and Wi<0.001 (Tab. 4), but this model did not provide as good fit to the data.
Results of model selection based on Akaike Information Criterion (AIC). Δi is the acceptability of each model, Wi is the plausibility of each model and k is the number of parameters from the nonlinear regression model.
Estimates of growth parameters (L ∞ , k e t 0 ) and confidence intervals for females and males Piaractus mesopotamicus of the Cuiabá River basin.
Based on the best model, the theoretical maximum average length (L∞ ) of females was slightly greater than that of males (64.99 cm and 63.23 cm, respectively), and females had a growth rate of 0.230 yr-1, while this was 0.196 yr-1 for males. Although average values of the maximum length (L∞ ) and growth rate (k) for females are slightly greater than that of males, there was no significant variation between the growth parameters, because there was an overlap in estimated parameter confidence intervals (Tab. 4, Fig. 5). Values for the growth parameters estimated in Model 1 (Individual) and Model 3 (Individual and site) are described in supplementary materials (Tab. S1 and S2).

Growth curves for females (black line) and males (gray line) for Piaractus mesopotamicus sampled in the Cuiabá River basin. The dotted lines are the lower and upper confidence interval limits of the estimated parameters for females (black) and males (gray).
The length-at-age data for P. mesopotamicus were highly variable (Fig. 5), with the largest dispersion of length measurements per age group observed in females. When the data length-at-age were analyzed by location and sex, females showed greater variation in the length data in the age, independent of sample site. However, females from the headwaters region had the greatest mean length by age, as well as the highest coefficient of variation (Tab. S3 and S4). Accordingly, from the descriptive analysis of variation in age-at-length data by sex and capture location, it appears that the greatest variation in length per age group occurred in females, as average length and longevity (maximum observed age) differed between sampling sites, and in the floodplain region only specimens 4 years old were sampled. In addition, average length of females (mean=42.85 cm and SD=5.14) and males (mean=41.62 cm and SD=3.64) was smaller in the floodplain than in the headwaters region (mean=48.73 cm and SD=9.75 for females and mean=42.65 cm and SD=5.29 for males). It should be noted that only in the headwaters sample were females significantly larger than males (ANOVA two-way for sex and location interaction: F1, 328 = 8.10, p <0.001). A model offering a more robust explanation for the variation in length-at-age data would examine the combined effect of location and sex but, due to non-convergence issues, such a model was not considered in this study.
Individual growth trajectories (Fig. 6) indicate that, regardless of sex, not all individuals have a similar growth profile in the average population. Some individuals had a growth performance above average and others below, and the range of variation from individual growth profile exceeded the confidence interval (Fig. 5).

Individual growth trajectories for Piaractus mesopotamicus collected in the Cuiabá River basin.
When the relationship between growth parameters, suggested by Rypel (2007Rypel AL. Sexual dimorphism in growth of freshwater drum. Southeastern Naturalist. 2007; 6(2):333-42.), was used to test the effect of sex on somatic growth, no differences in growth parameters of P. mesopotamicus was detected in relation to k and L∞ (Fig. 7a), t0 and L∞ (Fig. 7b) or between t0 and k (Fig. 7c).

Relationship between Piaractus mesopotamicus growth parameters: a. growth rate constant and asymptotic length b. age at length zero and asymptotic length and c. age at length zero and constant growth rate for males (black) and females (red).
Discussion
From marginal increment analysis it was determined that the formation of otolith growth rings occurred annually. Frequency of growth marks formation in otoliths, based on the marginal increment method (MI) indicated two band-forming periods; linked to the reduction in P. mesopotamicus growth rate, it is likely that the first band is formed during the late the ebb and early dry season, about a year after hatching of the cohort (Costa, Mateus, 2009Costa RMR, Mateus LAF. Reproductive biology of pacu Piaractus mesopotamicus (Holmberg, 1887) (Teleostei: Characidae) in the Cuiabá River Basin, Mato Grosso, Brazil. Neotrop Ichthyol. 2009; 7(3):447-58.). The results of this study confirm that cyclical process (dry season and wet) occur in tropical wetland environments with distinct seasonal variation, and that these have direct impacts on fish life cycles (Pérez, Fabré, 2009Pérez A, Fabré NN. Seasonal growth and life history of the catfish Calophysus macropterus (Lichtenstein, 1819) (Siluriformes: Pimelodidae) from the Amazon floodplain. J Appl Ichthyol. 2009; 25(3):343-49.). Consequently, seasonal growth variation was inferred from the relative widths of the two seasonal increments on the otoliths (Duponchelle et al., 2007Duponchelle F, Lino F, Hubert N, Panfili J, Renno JF, Baras E, Torrico JP, Dugue R, Nuñez J. Environment-related life-history trait variations of the red-bellied piranha Pygocentrus nattereri in two river basins of the Bolivian Amazon. J Fish Biol . 2007; 71(4):1113-34.). Ambrósio et al. (2014Ambrósio AM, Balbi TJ, Francisco TM, Gomes LC, Zuliani MS, Okada EK. Aging and growth parameter from the Piaractus mesopotamicus (pacu) from the Cuiabá river, Mato Grosso, Brazil. Iheringia Sér Zool. 2014; 104(4):413-17.) evaluated the deposition frequency of P. mesopotamicus growth rings in the Cuiabá River headwaters, and found that increment deposition was annual. Studies in seasonally-flooded environments suggest that the timing of ring formation in calcified and mineralized structures is annual and occurs during the dry season (Mateus, Petrere, 2004Mateus LAF, Petrere Júnior M. Age, growth and yield per recruit analysis of the Pintado Pseudoplatystoma corruscans (Agassiz, 1829) in the Cuiaba River Basin, Pantanal Matogrossense. Braz J Biol. 2004; 64(2):257-64.; Duponchelle et al., 2007Duponchelle F, Lino F, Hubert N, Panfili J, Renno JF, Baras E, Torrico JP, Dugue R, Nuñez J. Environment-related life-history trait variations of the red-bellied piranha Pygocentrus nattereri in two river basins of the Bolivian Amazon. J Fish Biol . 2007; 71(4):1113-34.). This pattern of growth ring deposition was also found for P. mesopotamicus by the current study. The maximum age of individual P. mesopotamicus was 5 years. Vaz (2001Vaz MM. Problemas no ajuste da curva de crescimento do pacu, Piaractus mesopotamicus (Holmberg, 1887) (Pisces: Characidae), e seu manejo no Pantanal Mato-Grossense. [PhD Thesis]. Jaboticabal, SP: UNESP; 2001.) found a similar age structure (5 years) based in scales readers, and Ambrósio et al. (2014Ambrósio AM, Balbi TJ, Francisco TM, Gomes LC, Zuliani MS, Okada EK. Aging and growth parameter from the Piaractus mesopotamicus (pacu) from the Cuiabá river, Mato Grosso, Brazil. Iheringia Sér Zool. 2014; 104(4):413-17.) observed the maximum of seven growth rings in the scales. Based on this study and previous (Vaz, 2001Vaz MM. Problemas no ajuste da curva de crescimento do pacu, Piaractus mesopotamicus (Holmberg, 1887) (Pisces: Characidae), e seu manejo no Pantanal Mato-Grossense. [PhD Thesis]. Jaboticabal, SP: UNESP; 2001.; Ambrósio et al., 2014Ambrósio AM, Balbi TJ, Francisco TM, Gomes LC, Zuliani MS, Okada EK. Aging and growth parameter from the Piaractus mesopotamicus (pacu) from the Cuiabá river, Mato Grosso, Brazil. Iheringia Sér Zool. 2014; 104(4):413-17.) the estimative from maximum age indicates a short life cycle for P. Mesopotamicus.
Based on the back-calculated lengths of both sexes, P. mesopotamicus has a greater increase in somatic growth in the first year of life, when it reaches an average length of 30 cm. Individuals then keep increasing until the second year and then reduce the rate of body growth. This slow-down in body growth increase rate is probably associated with the onset of gonadal maturation that occurs when total length reaches around 35 cm (Costa, Mateus, 2009Costa RMR, Mateus LAF. Reproductive biology of pacu Piaractus mesopotamicus (Holmberg, 1887) (Teleostei: Characidae) in the Cuiabá River Basin, Mato Grosso, Brazil. Neotrop Ichthyol. 2009; 7(3):447-58.), which was the average length of two-year old individuals.
Individual variability and sex were the main sources of variation in P. mesopotamicus somatic growth parameters. Implementing models that take into account the growth trajectory of individuals in a population has provided a better understanding of the source variation in the growth parameters, as the pattern that emerges at the population level is derived from processes generated at the individual level (Alós et al., 2010Alós J, Palmer M, Alonso-Fernández A, Morales-Nin B. Individual variability and sex-related differences in the growth of Diplodus annularis (Linnaeus, 1758). Fish Res. 2010; 101(1-2):60-69.; Vincenzi et al., 2014Vincenzi S, Mangel M, Crivelli AJ, Munch S, Skaug HJ. Determining individual variation in growth and its implication for life-history and population processes using the empirical Bayes Method. PLoS Computational Biology. 2014; 10(9):1-16.; DeAngelis, Grimm, 2014DeAngelis DL, Grimm V. Individual-based models in ecology after four decades. F1000Prime Reports. 2014; 6(39):1-6.). Furthermore, the use of an approach at the individual level is more appropriate since individual variation is controlled by considering the growth trajectory of each specimen. This means that the source of changes in growth trajectory generated by other biotic factors or abiotic could be evaluated more robustly (Pilling et al., 2002Pilling GM, Kirkwood GP, Walker SG. An improved method for estimating individual growth variability in fish, and the correlation between von Bertalanffy growth parameters. Can J Fish Aquat Sci . 2002; 59(3):424-32.).
Food resources are among the main factors that can alter fish physiology, metabolic rate, growth and deposition pattern of spatial growth increments (Williams et al., 2012Williams AJ, Farley JH, Hoyle SD, Davies CR, Nicol SJ. Spatial and sex-specific variation in growth of albacore tuna (Thunnus alalunga) across the South Pacific Ocean. Plos One. 2012; 7(6):1-10.; Shelton et al., 2013Shelton AO, Satterthwaite WH, Beakes MP, Munch SB, Sogard SM, Mangel M. Separating intrinsic and environmental contributions to growth and their population consequences. Am Nat. 2013; 181(6):799-814.; Vincenzi et al., 2014Vincenzi S, Mangel M, Crivelli AJ, Munch S, Skaug HJ. Determining individual variation in growth and its implication for life-history and population processes using the empirical Bayes Method. PLoS Computational Biology. 2014; 10(9):1-16.). However, for the species studied, exogenous factors such as food resources are unlikely to be a limiting factor for growth because individuals migrate between habitats in search of food (trophic migration). Except, of course, in situations where there are barriers that impede fish mobility between environments, as in the case of natural blockages (ox-bows lakes isolated in the dry season in the floodplain) or human-made dams. The best model showed individual and sex effect on growth parameters. The sex effect in fish species, with females being larger than males or vice versa (Rljnsdorp, Ibelings, 1989Rljnsdorp AD, Ibelings B. Sexual dimorphism in the energetics of reproduction and growth of North Sea plaice, Pleuronectes platessa L. J Fish Biol . 1989; 35(3):401-15.; Imsland et al., 1997Imsland AK, Folkvord A, Grung GL, Stefansson SO, Taranger GL. Sexual dimorphism in growth and maturation of turbot, Scophthalmus maximus (Rafinesque, 1810). Aquac Res. 1997; 28(2):101-14.; Ambrósio et al., 2014Ambrósio AM, Balbi TJ, Francisco TM, Gomes LC, Zuliani MS, Okada EK. Aging and growth parameter from the Piaractus mesopotamicus (pacu) from the Cuiabá river, Mato Grosso, Brazil. Iheringia Sér Zool. 2014; 104(4):413-17.) is not rare to detect. The difference in body length between males and females is related to variation in growth parameters (Morita et al., 2015Morita SH, Morita K, Hamatsu T, Chimura M, Yamashita Y, Sasaki K, Sato T. Differential effects of the environment on the growth of arabesque greenling (Pleurogrammus azonus): does rising temperature benefit young but not old fish? Environ Biol Fish. 2015; 98(3):801-09.). From the adaptive point of view, this variation allows greater fecundity per unit biomass, since larger females can produce more eggs (Costa, Mateus, 2009Costa RMR, Mateus LAF. Reproductive biology of pacu Piaractus mesopotamicus (Holmberg, 1887) (Teleostei: Characidae) in the Cuiabá River Basin, Mato Grosso, Brazil. Neotrop Ichthyol. 2009; 7(3):447-58.), or the production of larger eggs with greater food reserves and thus the promotion of enhanced larval survival rates (Stearns, 1976Stearns SC. Life-history tactics: a review of the ideas. Quart Rev Biol. 1976; 51(1):3-47.; Araújo, Haimovici, 2000Araújo JN, Haimovici M. Determinação de idades e crescimento do linguado branco Paralichthys patagonicus (Jordan, 1889) no sul do Brasil. Rev Bras Oceanogr. 2000; 48(1):61-70.).
Length-at-age data for P. mesopotamicus from the current study were highly variable. However, it is notable that studies of same age of the P. mesopotamicus in cultivation cages have shown inter-individual variation in somatic growth when stocking density is altered (Bittencourt et al., 2010Bittencourt F, Feiden A, Signor AA, Boscolo WR, Lorenz EK, Maluf MLF. Densidade de estocagem e parâmetros eritrocitários de pacus criados em tanques-rede. Rev Bras Zootec. 2010; 39(11):2323-29.). In the current study, variation in growth was explained by individual variation and sex, with the analyzed individuals showing different growth potential. Hence, some individuals (but not others) may be genetically predisposed to rapid growth, and due to individual (genetic) differences. Thus, variation in growth arises because although individuals are operating in the same biotic and abiotic conditions, growth performance of the individuals within the population is not homogeneous.
Knowledge of the life history traits of commercialized fish species is a basic requirement to enable proposals for rational use and effective management planning of stocks (Bayley, Petrere, 1989Bayley PB, Petrere Júnior M. Amazon fisheries: assessment methods, current status and management options. In: Dodge DP, editor. Proceedings of the International Large River Symposium. Can Spec Public Fish Aquat Sci; 1989; 106:385-398.). The inclusion of the effect of individual variability (genotype), sex and locality on growth parameters contributes to a better and more nuanced understanding of the variation of somatic growth of fish. It can also provide more accurate estimates of growth parameters and so generate less biased information that can be useful when evaluating stock inventories. This approach is important because the use of traditional models (which disregard the effects of individual variability and factors like sex) can lead to erroneous assumptions about the best ways to manage fish stocks (Pilling et al., 2002Pilling GM, Kirkwood GP, Walker SG. An improved method for estimating individual growth variability in fish, and the correlation between von Bertalanffy growth parameters. Can J Fish Aquat Sci . 2002; 59(3):424-32.; Vigliola, Meekan, 2009Vigliola L, Meekan MG. The back-calculation of fish growth from otoliths. In: Green BS, Mapstone BD, Carlos G, Begg GA, editors. Tropical Fish Otoliths: Information for Assessment, Management and Ecology. Reviews: Methods and Technologies in Fish Biology and Fisheries. New York: Springer ; 2009. p.174-211.).
Because of its high commercial value, Piaractus mesopotamicus one the most important species in the Paraguay River basin, and is one of the species most sought after by amateur and professional fishermen (Peixer et al., 2007Peixer J, Petrere Júnior M. Hook selectivity of the pacu Piaractus mesopotamicus (Holmberg, 1887) in the Pantanal, the state of Mato Grosso do Sul, Brazil. Braz J Biol . 2007; 67(2):339-45.). Therefore, the results from this study can be practically applied because the models used in the management of fishery require not only stock biomass but also a knowledge of the age structure and quantified growth parameters.
In summary, it can be concluded that the variation in the growth parameters from P. mesopotamicus was due to sex and individual characteristics. Locality is not an important factor in the proposed model, because the species is a long-distance migrant, and does not depend on local conditions to meet the nutritional needs. However, the site is nonetheless an essential factor to complete the life cycle of the species, separate localities can provide shelter and food at different stages of the P. mesopotamicus life cycle.
Acknowledgments
The authors thank Nilso Estevão da Silva and Davino Dias de Campos for capturing the samples, Estância Ecológica Sesc Pantanal and Fazenda Santo Antônio for logistic support. Centro de Pesquisa do Pantanal (CPP)/ Ministério da Ciência e Tecnologia (MCT) and Programa PELD (Sítio 12) for financial assistance. L.S. Lourenço thanks FAPEMAT∕CNPq for Desenvolvimento Científico e Regional - DCR grant. Adrian Barnett helped with the English.
References
- Alós J, Palmer M, Alonso-Fernández A, Morales-Nin B. Individual variability and sex-related differences in the growth of Diplodus annularis (Linnaeus, 1758). Fish Res. 2010; 101(1-2):60-69.
- Ambrósio AM, Balbi TJ, Francisco TM, Gomes LC, Zuliani MS, Okada EK. Aging and growth parameter from the Piaractus mesopotamicus (pacu) from the Cuiabá river, Mato Grosso, Brazil. Iheringia Sér Zool. 2014; 104(4):413-17.
- Arantes CC, Castello L, Stewart DJ, Cetra M, Queiroz HL. Population density, growth and reproduction of arapaima in an Amazonian river-floodplain. Ecol Freshw Fish. 2010; 19(3):455-65.
- Araújo JN, Haimovici M. Determinação de idades e crescimento do linguado branco Paralichthys patagonicus (Jordan, 1889) no sul do Brasil. Rev Bras Oceanogr. 2000; 48(1):61-70.
- Bayley PB, Petrere Júnior M. Amazon fisheries: assessment methods, current status and management options. In: Dodge DP, editor. Proceedings of the International Large River Symposium. Can Spec Public Fish Aquat Sci; 1989; 106:385-398.
- Beerli EL, Logato PVR, Freitas RTF. Alimentação e comportamento de larvas de pacu Piaractus mesopotamicus (Holmberg, 1887). Ciênc Agrotecnol. 2004; 28(1):149-55.
- Bittencourt F, Feiden A, Signor AA, Boscolo WR, Lorenz EK, Maluf MLF. Densidade de estocagem e parâmetros eritrocitários de pacus criados em tanques-rede. Rev Bras Zootec. 2010; 39(11):2323-29.
- Britski HA, Silimon KZS, Lopes BS. Peixes do pantanal. Manual de identificação. Brasília: Embrapa -SPI; 1999.
- Burnham KP, Anderson DR. Model selection and inference: a practical information - theoretic approach. 2nd ed. New York: Springer; 2002.
- Campana SE. How reliable are growth back-calculations based on Otoliths? Can J Fish Aquat Sci. 1990; 47:2219-37.
- Campana SE. Accuracy, precision and quality control in age determination, including a review of the use and abuse of age validation methods. J Fish Biol. 2001; 59(2):197-242.
- Catella AC. A pesca no pantanal de Mato Grosso do Sul, Brasil: descrição, nível de exploração e manejo (1994-1999). [PhD Thesis]. Manaus, AM: Instituto Nacional de Pesquisa da Amazônia; 2001.
- Cavinatto V. Caracterização Hidrográfica do Estado de Mato Grosso. Cuiabá: Prodeagro/Seplan/Fema-MT. 1995.
- Cerrato RM. Analysis of nonlinearity effects in expected-value parametrizations of the von Bertalanffy equation. Can J Fish Aquat Sci . 1991; 48(11):2109-17.
- Correa SB, Araujo JK, Penha JMF, Cunha CN, Stevenson PR, Anderson JT. Overfishing disrupts an ancient mutualism between frugivorous fishes and plants in Neotropical wetlands. Biol Conserv. 2015; 191:159-67.
- Costa RMR, Mateus LAF. Reproductive biology of pacu Piaractus mesopotamicus (Holmberg, 1887) (Teleostei: Characidae) in the Cuiabá River Basin, Mato Grosso, Brazil. Neotrop Ichthyol. 2009; 7(3):447-58.
- Da Silva CJ, Esteves FA. Dinâmica das características limnológicas das baías Porto de Fora e Acurizal (Pantanal de Mato Grosso) em função da variação do nível da água. Oecol Brasil. 1995; 1:47-60.
- DeAngelis DL, Grimm V. Individual-based models in ecology after four decades. F1000Prime Reports. 2014; 6(39):1-6.
- Dieterman DJ, Hoxmeier RJH, Staples DF. Factors influencing growth of individual brown trout in three streams of the upper Midwestern United States. Ecol Freshw Fish . 2012; 21(3):483-93.
- Dmitriew CM. The evolution of growth trajectories: what limits growth rate? Biol Rev. 2011; 86(1):97-116.
- Duponchelle F, Lino F, Hubert N, Panfili J, Renno JF, Baras E, Torrico JP, Dugue R, Nuñez J. Environment-related life-history trait variations of the red-bellied piranha Pygocentrus nattereri in two river basins of the Bolivian Amazon. J Fish Biol . 2007; 71(4):1113-34.
- Fantin-Cruz I, Pedrollo O, Castro NMR, Girard P, Zeilhofer P, Hamilton SK. Historical reconstruction of floodplain inundation in the Pantanal (Brazil) using neural networks. J Hydrol. 2011; 399(3-4):376-84.
- Figueiredo DM, Salomão FXT. Bacia do Rio Cuiabá: Uma abordagem socioambiental.Cuiabá: EdUFMT; 2009.
- Francis RICC. Are growth parameters estimated from tagging and age-length data comparable? Can J Fish Aquat Sci . 1988; 45(6):936-42.
- Green BS, Mapstone BD, Carlos G, Begg GA. Introduction to otoliths and fisheries in the tropics. In: Green BS, Mapstone BD, Carlos G, Begg GA, editors. Tropical Fish Otoliths: Information for Assessment, Management and Ecology. Reviews: Method and Technologies in Fish Biology and Fisheries. New York: Springer ; 2009. p.1-22.
- Hamilton SK, Sippel SJ, Melack JM. Inundation patterns in the Pantanal wetland of South America determined from passive microwave remote sensing. Arch Hydrobiol. 1996; 137(1):1-23.
- Henryon M, Jokumsen A, Berg P, Lund I, Pedersen PB, Olesen NJ, Slierendrecht WJ. Genetic variation for growth rate, feed conversion efficiency, and disease resistance exists within a farmed population of rainbow trout. Aquaculture. 2002; 209(1-4):59-76.
- Imsland AK, Folkvord A, Grung GL, Stefansson SO, Taranger GL. Sexual dimorphism in growth and maturation of turbot, Scophthalmus maximus (Rafinesque, 1810). Aquac Res. 1997; 28(2):101-14.
- Katsanevakis S. Modelling fish growth: model selection, multi-model inference and model selection uncertainty. Fish Res . 2006; 81(2):229-35.
- Kielbassa J, Charles S, Delignette-Muller ML. The importance of incorporating age and sex when backcalculating length in bullhead Cottus gobio J Fish Biol . 2011; 78(5):1492-507.
- Lai HL, Gallucci VF, Gunderson DR, Donnelly RF. Age determination in fisheries: methods and applications to stock assessment. In: Galucci VF, Saila SB, Gunderson DJ, Rothschild BJ, editors. Stock Assessment: Quantitative Methods and Applications for Small-Scale Fisheries. New York: Lewis Publishers - CRC Press; 1996. p.82-178.
- Mateus LAF, Petrere Júnior M. Age, growth and yield per recruit analysis of the Pintado Pseudoplatystoma corruscans (Agassiz, 1829) in the Cuiaba River Basin, Pantanal Matogrossense. Braz J Biol. 2004; 64(2):257-64.
- Mateus LAF, Penha JMF, Petrere M. Fishing resources in the rio Cuiabá Basin, Pantanal do Mato Grosso, Brazil. Neotrop Ichthyol . 2004; 2(4):217-27.
- Morita SH, Morita K, Hamatsu T, Chimura M, Yamashita Y, Sasaki K, Sato T. Differential effects of the environment on the growth of arabesque greenling (Pleurogrammus azonus): does rising temperature benefit young but not old fish? Environ Biol Fish. 2015; 98(3):801-09.
- Peixer J, Catella AC, Petrere Júnior M. Yield per recruit of pacu Piaractus mesopotamicus (Holmberg, 1887) in the Pantanal of Mato Grosso do Sul, Brazil. Braz J Biol . 2007; 67(3):561-67.
- Peixer J, Petrere Júnior M. Hook selectivity of the pacu Piaractus mesopotamicus (Holmberg, 1887) in the Pantanal, the state of Mato Grosso do Sul, Brazil. Braz J Biol . 2007; 67(2):339-45.
- Pérez A, Fabré NN. Seasonal growth and life history of the catfish Calophysus macropterus (Lichtenstein, 1819) (Siluriformes: Pimelodidae) from the Amazon floodplain. J Appl Ichthyol. 2009; 25(3):343-49.
- Pilling GM, Kirkwood GP, Walker SG. An improved method for estimating individual growth variability in fish, and the correlation between von Bertalanffy growth parameters. Can J Fish Aquat Sci . 2002; 59(3):424-32.
- R Development Core Team. R: A language and environment for statistical computing. [Computer software manual - Internet]. Vienna: R Foundation for Statistical Computing; 2014. Available from: http://www.R-project.org/
» http://www.R-project.org/ - Ratkowsky DA. Statistical properties of alternative parameterizations of the von Bertalanffy growth curve. Can J Fish Aquat Sci . 1986; 43(4):742-47.
- Ratkowsky DA. Handbook of Nonlinear Regression Models. New York: Marcel Dekker; 1990.
- Reis RE, Kullander SO, Ferraris Júnior CJ. Check list of the freshwater fishes of South and Central America. Porto Alegre: EDIPUCRS; 2003.
- Rljnsdorp AD, Ibelings B. Sexual dimorphism in the energetics of reproduction and growth of North Sea plaice, Pleuronectes platessa L. J Fish Biol . 1989; 35(3):401-15.
- Rypel AL. Sexual dimorphism in growth of freshwater drum. Southeastern Naturalist. 2007; 6(2):333-42.
- Schnute J, Fournier D. A new approach to length-frequency analysis: growth structure. Can J Fish Aquat Sci . 1980; 37(9):1337-51.
- Secor DH, Dean JM, Laban EH. Otolith removal and preparation for microstructural examination. In: Stevenson DK, Campana SE, editors. Otolith Microstructure Examination and Analysis. Ottawa: Department of Fisheries and Oceans; 1992. p.19-57.
- Shelton AO, Satterthwaite WH, Beakes MP, Munch SB, Sogard SM, Mangel M. Separating intrinsic and environmental contributions to growth and their population consequences. Am Nat. 2013; 181(6):799-814.
- Siangas EL, Pouilly M, Vallejos A, Pérez T, Rejas D. Effect of water quality on growth of four fish species in the Iténez basin (Upper Madera, Amazon). Environ Biol Fish . 2012; 95(3):371-81.
- Sokal RR, Rohlf FJ. Biometry: the principles of statistics in biological research. 3rd ed. New York: Freeman; 1995.
- Stearns SC. Life-history tactics: a review of the ideas. Quart Rev Biol. 1976; 51(1):3-47.
- Vaz MM. Problemas no ajuste da curva de crescimento do pacu, Piaractus mesopotamicus (Holmberg, 1887) (Pisces: Characidae), e seu manejo no Pantanal Mato-Grossense. [PhD Thesis]. Jaboticabal, SP: UNESP; 2001.
- Vigliola L, Meekan MG. The back-calculation of fish growth from otoliths. In: Green BS, Mapstone BD, Carlos G, Begg GA, editors. Tropical Fish Otoliths: Information for Assessment, Management and Ecology. Reviews: Methods and Technologies in Fish Biology and Fisheries. New York: Springer ; 2009. p.174-211.
- Vincenzi S, Mangel M, Crivelli AJ, Munch S, Skaug HJ. Determining individual variation in growth and its implication for life-history and population processes using the empirical Bayes Method. PLoS Computational Biology. 2014; 10(9):1-16.
- Weisberg S, Spangler G, Richmond LS. Mixed effects models for fish growth. Can J Fish Aquat Sci . 2010; 67(2):269-77.
- Welsford DC, Lyle JM. Estimates of growth and comparisons of growth rates determined from length- and age-based models for populations of purple wrasse (Notolabrus fucicola). Fish Bull. 2005; 103(4):697-711.
- Williams AJ, Farley JH, Hoyle SD, Davies CR, Nicol SJ. Spatial and sex-specific variation in growth of albacore tuna (Thunnus alalunga) across the South Pacific Ocean. Plos One. 2012; 7(6):1-10.
- Zeilhofer P, Moura RM. Hydrological changes in the northern Pantanal caused by the Manso dam: Impact analysis and suggestions for mitigation. Ecol Engin. 2009; 35(1):105-17.
- Ziober SR, Bialetzki A, Mateus LAF. Effect of abiotic variables on fish eggs and larvae distribution in headwaters of Cuiabá River, Mato Grosso State , Brazil. Neotrop Ichthyol . 2012; 10(1):123-32.
Publication Dates
-
Publication in this collection
2017
History
-
Received
04 Aug 2016 -
Accepted
03 May 2017