Abstract
Among Neotropical freshwater ecoregions, the Mid-Northeastern Caatinga (MNCE) is a fish knowledge gap. Its temporary drainages are receptors of the São Francisco interbasin water transfer project (SFR-IWT) in the Brazilian semiarid. We provide a comprehensive baseline of fish richness of the five SFR-IWT basins. Species richness, shared, endemic, threatened and non-native species were obtained using sampling, ichthyologic collections, literature and online repositories (306 localities). In total 121, species were recorded, 111 of them native, and 16 (14.41%) listed for all basins. Higher richness of native species (78, 70.27%) was recorded in the São Francisco lower-middle stretch (SFRE), including 23 endemic, 61 (54.95%) in MNCE basins (13 endemic), and 28 (25.23%) shared between both ecoregions. In the MNCE, 50 species were recorded in Jaguaribe (JAG), 39 in Piranhas-Açu, 36 in Paraíba do Norte, and 32 in Apodi-Mossoró (APO). The number of species shared between the SFRE and each receptor basin varied from 24 (23.08%, JAG) to 20 (22.22%, APO). JAG contains 81.97% of the receptor basins’ species. Its higher richness and endemism deserve special attention regarding the ongoing hydrological changes. This study will help detect possible modifications in the ichthyofauna of the main MNCE drainages.
Keywords:
Anthropogenic impacts; Biological invasions; Caatinga’s fishes; Mid-Northeastern Caatinga Ecoregion; Temporary rivers
Resumo
Dentre as ecorregiões dulcícolas neotropicais, a Nordeste Médio-Oriental (MNCE) é uma lacuna de conhecimento sobre peixes. Suas drenagens temporárias são receptoras do projeto de transposição do São Francisco (SFR-IWT) no semiárido brasileiro. Fornecemos uma linha de base abrangente da riqueza de peixes das cinco bacias do SFR-IWT. Riqueza de espécies, espécies compartilhadas, endêmicas, ameaçadas e não-nativas foram obtidas através de amostragem, coleções ictiológicas, literatura e repositórios online (306 localidades). No total, foram registradas 121 espécies, 111 nativas e 16 (14,41%) listadas para todas as bacias. A maior riqueza de espécies nativas (78, 70,27%) foi registrada no trecho sub-médio do São Francisco (SFRE), incluindo 23 endêmicas e 61 (54,95%) nas bacias da MNCE (13 endêmicas), e 28 (25,23 %) compartilhadas entre ambas ecorregiões. Na MNCE, foram registradas 50 espécies na Jaguaribe (JAG), 39 na Piranhas-Açu, 36 na Paraíba do Norte e 32 na Apodi-Mossoró (APO). O número de espécies compartilhadas entre SFRE e cada bacia receptora variou de 24 (23,08%, JAG) a 20 (22,22%, APO). JAG contém 81,97% das espécies das bacias receptoras. Sua maior riqueza e endemismo merecem atenção especial em relação às mudanças hidrológicas em curso. Este estudo ajudará a detectar possíveis modificações na ictiofauna das principais drenagens da MNCE.
Palabras-chave:
Ecorregião Caatinga NordesteMédio Oriental; Impactos antropogênicos; Invasões biológicas; Peixes da Caatinga; Rios temporários
INTRODUCTION
Global aquatic ecosystems have been losing part of their biodiversity in response to increasingly deleterious anthropogenic pressure, caused mainly by rampant population growth, preposterous consumption of natural resources, energy production, and the absence of efficient management and conservation mechanisms (Abell et al., 2008Abell R, Thieme ML, Revenga C, Bryer M, Kottelat M, Bogutskaya N et al. Freshwater ecoregions of the world: a new map of biogeographic units for freshwater biodiversity conservation. Bioscience. 2008; 58(5):403-14. https://doi.org/10.1641/B580507
https://doi.org/10.1641/B580507...
; Barletta et al., 2010Barletta M, Jaureguizar AJ, Baigun C, Fontoura NF, Agostinho AA, Almeida-Val VMF et al. Fish and aquatic habitat conservation in South America: a continental overview with emphasis on Neotropical systems. J Fish Biol. 2010; 76(9):2118-76. https://doi.org/10.1111/j.1095-8649.2010.02684.x
https://doi.org/10.1111/j.1095-8649.2010...
). This scenario is even more worrying in freshwater systems, which, besides having a smaller physical area, have been historically undergoing severe environmental alterations, such as damming, illegal water catchment, introduction of species and contamination by pesticides and effluents (L’vovich, White, 1990L’vovich MI, White GF. Use and transformation of terrestrial water systems. In: Turner-II BL, Clark WC, Kates RW, Richards JF, Mathews JT, Meyer WB, editors. The Earth as transformed by human action. University Cambridge Press; 1990. p.235-52.; Meybeck, 2003Meybeck M. Global analysis of river systems: from Earth system controls to Anthropocene syndromes. Philos Trans R Soc Lond B Biol Sci. 2003; 358(1440):1935-55. https://doi.org/10.1098/rstb.2003.1379
https://doi.org/10.1098/rstb.2003.1379...
; Dudgeon, 2019Dudgeon D. Multiple threats imperil freshwater biodiversity in the Anthropocene. Curr Biol. 2019; 29(19):960-7. https://doi.org/10.1016/j.cub.2019.08.002
https://doi.org/10.1016/j.cub.2019.08.00...
). All these impacts have a cumulative effect by reducing habitat quality and quantity and increasing extinction risk for freshwater fishes, making those systems into one of the most threatened worldwide (Meybeck, 2003Meybeck M. Global analysis of river systems: from Earth system controls to Anthropocene syndromes. Philos Trans R Soc Lond B Biol Sci. 2003; 358(1440):1935-55. https://doi.org/10.1098/rstb.2003.1379
https://doi.org/10.1098/rstb.2003.1379...
; Revenga et al., 2005Revenga C, Campbell I, Abell R, Villiers P, Bryer M. Prospects for monitoring freshwater ecosystems towards the 2010 targets. Philos Trans R Soc Lond B Biol Sci. 2005; 360(1454):397-413. https://doi.org/10.1098/rstb.2004.1595
https://doi.org/10.1098/rstb.2004.1595...
; Abell et al., 2008Abell R, Thieme ML, Revenga C, Bryer M, Kottelat M, Bogutskaya N et al. Freshwater ecoregions of the world: a new map of biogeographic units for freshwater biodiversity conservation. Bioscience. 2008; 58(5):403-14. https://doi.org/10.1641/B580507
https://doi.org/10.1641/B580507...
; Reis et al., 2016Reis RE, Albert JS, Di Dario F, Mincarone MM, Petry P, Rocha LA. Fish biodiversity and conservation in South America. J Fish Biol. 2016; 89(1):12-47. https://doi.org/10.1111/jfb.13016
https://doi.org/10.1111/jfb.13016...
; Castro, Polaz, 2020Castro RMC, Polaz CNM. Small-sized fish: the largest and most threatened portion of the megadiverse neotropical freshwater fish fauna. Biota Neotrop. 2020; 20(1):e20180683. https://doi.org/10.1590/1676-0611-bn-2018-0683
https://doi.org/10.1590/1676-0611-bn-201...
).
One of the main issues related to aquatic ecosystems is water scarcity, especially in arid and semiarid areas. When the demand for water increases and almost reaches the available limit, debates over management and attainment of new resources emerge as an attempt to converge on an integrated management model (L’vovich, White, 1990L’vovich MI, White GF. Use and transformation of terrestrial water systems. In: Turner-II BL, Clark WC, Kates RW, Richards JF, Mathews JT, Meyer WB, editors. The Earth as transformed by human action. University Cambridge Press; 1990. p.235-52.; Meybeck, 2003Meybeck M. Global analysis of river systems: from Earth system controls to Anthropocene syndromes. Philos Trans R Soc Lond B Biol Sci. 2003; 358(1440):1935-55. https://doi.org/10.1098/rstb.2003.1379
https://doi.org/10.1098/rstb.2003.1379...
; Gupta, Van der Zaag, 2008Gupta J, Van der Zaag P. Interbasin water transfers and integrated water resources management: Where engineering, science and politics interlock. Phys Chem Earth. 2008; 33(1-2):28-40. https://doi.org/10.1016/j.pce.2007.04.003
https://doi.org/10.1016/j.pce.2007.04.00...
). Some countries have solved this problem by expanding their hydrographic network through aqueducts and canals (Van der Zaag, 2007Van der Zaag P. Asymmetry and equity in water resources management; critical governance issues for Southern Africa. Water Resour Manag. 2007; 21(12):1993-2004. https://doi.org/10.1007/s11269-006-9124-1
https://doi.org/10.1007/s11269-006-9124-...
; Gupta, Van der Zaag, 2008Gupta J, Van der Zaag P. Interbasin water transfers and integrated water resources management: Where engineering, science and politics interlock. Phys Chem Earth. 2008; 33(1-2):28-40. https://doi.org/10.1016/j.pce.2007.04.003
https://doi.org/10.1016/j.pce.2007.04.00...
). Currently, water transfers projects have started or are under discussion in South America, in the São Francisco Interbasin Water Transfer Project (SFR-IWT, in Brazil), Africa, in the Lesotho Highland Water Project (between Lesotho and South Africa) and Asia, in the South to North Water Diversion Project (SNWDP, in China), and in the Par Tapi Narmada Link Project (PTNLP, in India).These projects differ, mainly in the number of connected ecoregions, basins, canal length, and captured water volume (Brasil, 2004Brasil. Relatório de Impacto Ambiental-RIMA do Projeto de Integração do rio São Francisco com as Bacias Hidrográficas do Nordeste Setentrional. Brasília: Ministério da Integração Nacional/Ecology and Environment do Brasil; 2004.; India, 2017India (National Water Development Agency). Par Tapi Narmada Link Project. India: Ministry of Jal Shakti; 2017. p.844–926. Available from: http:// nwda.gov.in/content/innerpage/par-tapinarmada-link-project.php
http:// nwda.gov.in/content/innerpage/pa...
; Lesotho, 2020Lesotho (Lesotho Highlands Development Authority). Project description [Internet]. Lesotho; 2020. Available from: http://www.lhda.org.ls/lhdaweb
http://www.lhda.org.ls/lhdaweb...
; Long et al., 2020Long D, Yang W, Scanlon BR, Zhao J, Liu D, Burek P et al. South-to-North Water Diversion stabilizing Beijing’s groundwater levels. Nat Commun. 2020; 11(3665). https://doi.org/10.1038/s41467-020-17428-6
https://doi.org/10.1038/s41467-020-17428...
).
However, such projects are considered as imminent threats to freshwater biodiversity (Albert et al., 2020Albert JS, Destouni G, Duke-Sylvester SM, Magurran AE, Oberdorff T, Reis RE et al. Scientist's warning to humanity on the freshwater biodiversity crisis. Ambio. 2020. https://doi.org/10.1007/s13280-020-01318-8
https://doi.org/10.1007/s13280-020-01318...
) and face many criticisms and heated debates over the socio-economic benefits versus environmental issues they can cause (Meador, 1992Meador MR. Inter-basin water transfer: Ecological concerns. Fisheries. 1992; 17(2):17-22. https://doi.org/10.1577/1548-8446(1992)017<0017:IWTEC>2.0.CO;2
https://doi.org/10.1577/1548-8446(1992)0...
; Muller, 1999Muller M. Interbasin water sharing: A South African perspective. In: Proceedings of the International Workshop. Paris: UNESCO, Technical Documents in Hydrology 28, International Hydrological Program - V; 1999. p.61-70.; Dyrnes, Vatn, 2005Dyrnes GV, Vatn A. Who owns the water? A study of a water conflict in the Valley of Ixtlahuaca, México. Water Policy. 2005; 7(3):295-312. https://doi.org/10.2166/wp.2005.0019
https://doi.org/10.2166/wp.2005.0019...
; Pittock et al., 2009Pittock J, Meng JH, Geiger M, Chapagain AK. Interbasin water transfers and water scarcity in a changing world - a solution or a pipedream?. Germany: WWF; 2009.). The latter include species introductions, loss of local adaptations and biota homogenization, genetic introgression, competitive exclusion, hybridization, habitat modification (including terrestrial vegetation fragmentation by canals), and water availability and quality (Meador, 1992Meador MR. Inter-basin water transfer: Ecological concerns. Fisheries. 1992; 17(2):17-22. https://doi.org/10.1577/1548-8446(1992)017<0017:IWTEC>2.0.CO;2
https://doi.org/10.1577/1548-8446(1992)0...
; Pittock et al., 2009Pittock J, Meng JH, Geiger M, Chapagain AK. Interbasin water transfers and water scarcity in a changing world - a solution or a pipedream?. Germany: WWF; 2009.; Albert et al., 2020Albert JS, Destouni G, Duke-Sylvester SM, Magurran AE, Oberdorff T, Reis RE et al. Scientist's warning to humanity on the freshwater biodiversity crisis. Ambio. 2020. https://doi.org/10.1007/s13280-020-01318-8
https://doi.org/10.1007/s13280-020-01318...
).
Water shortage is one of the main problems in the Brazilian Northeast region, which is mostly inserted in the semiarid Caatinga (Lima et al., 2017Lima SMQ, Ramos TPA, da Silva MJ, Rosa RS. Diversity, distribution, and conservation of the Caatinga fishes: advances and challenges. In: Silva, JMC, Leal IR, Tabarelli M, editors. Caatinga the largest tropical dry forest region in South America. Springer; 2017. p.97-131. https://doi.org/10.1007/978-3-319-68339-3_4
https://doi.org/10.1007/978-3-319-68339-...
). It is a highly impacted area, some of which is in the process of desertification (Albuquerque et al., 2012Albuquerque UP, Araújo EL, El-Deir ACA, Lima ALA, Souto A, Bezerra BM et al. Caatinga revisited: ecology and conservation of an important seasonal dry forest. Sci World J. 2012; 205182:1-18. https://doi.org/10.1100/2012/205182
https://doi.org/10.1100/2012/205182...
), and whose aquatic habitats are considered the most modified and threatened in the country (Silva et al., 2004Silva JMC, Tabarelli M, Fonseca MT, Lins LV, editors. Biodiversidade da Caatinga: áreas e ações prioritárias para a conservação. Brasília: Ministério do Meio Ambiente; 2004.). Recurrent droughts limit water supply and demand huge engineering projects (Rosa et al., 2004Rosa RS, Gomes-Filho G, Menezes NA, Shibatta OA, Costa WJEM. Biota aquática: áreas e ações prioritárias para conservação da caatinga. In: Silva JMC, Tabarelli M, Fonseca MT, Lins LV, editors. Biodiversidade da caatinga: áreas e ações prioritárias para a conservação. Brasília: Ministério do Meio Ambiente; 2004. p.163-71.).
The idea to transpose waters from the rio São Francisco was introduced in 1847 by the engineer Marcos de Macedo to Emperor Dom Pedro II. In 1913, after the ‘Great Drought’ (1875-1879), the construction of a canal that would connect the São Francisco and Jaguaribe basins was outlined. Nevertheless, it became impracticable due to technical reasons, lack of financial resources and ignorance of its benefits (Andrade et al., 2011Andrade JGP, Barbosa PSF, Souza LCA, Makino DL. Interbasin water transfers: The Brazilian experience and international case comparisons. Water Resour Manag. 2011; 25(8):1915-34. https://doi.org/10.1007/s11269-011-9781-6
https://doi.org/10.1007/s11269-011-9781-...
). After other frustrated attempts, in 2005 the project was redesigned and started to be known as the São Francisco Interbasin Water Transfer to the Northeastern Hydrographic Basins Project (SFR-IWT, Projeto de Integração do Rio São Francisco com as Bacias Hidrográficas do Nordeste Setentrional, in Portuguese) (Brasil, 2004Brasil. Relatório de Impacto Ambiental-RIMA do Projeto de Integração do rio São Francisco com as Bacias Hidrográficas do Nordeste Setentrional. Brasília: Ministério da Integração Nacional/Ecology and Environment do Brasil; 2004.).
The SFR-IWT aims to transfer 3.5% of the São Francisco basin water to the Jaguaribe, Apodi-Mossoró, Piranhas-Açu, and Paraíba do Norte basins, through a two-canal system (North and East Axis), aqueducts, tunnels, dams, and pumping stations, resulting in 720 km of extension (577 km, excepting the secondary canals, 357 km in the North Axis and 220 km in the East Axis), whose catchment areas are located in the lower-middle stretch of the São Francisco basin (Brasil, 2004Brasil. Relatório de Impacto Ambiental-RIMA do Projeto de Integração do rio São Francisco com as Bacias Hidrográficas do Nordeste Setentrional. Brasília: Ministério da Integração Nacional/Ecology and Environment do Brasil; 2004.). Since its approval in 2007 and the beginning of construction, in 2008 (Andrade et al., 2011Andrade JGP, Barbosa PSF, Souza LCA, Makino DL. Interbasin water transfers: The Brazilian experience and international case comparisons. Water Resour Manag. 2011; 25(8):1915-34. https://doi.org/10.1007/s11269-011-9781-6
https://doi.org/10.1007/s11269-011-9781-...
), the project has faced many delays but is now almost concluded, with the East Axis in operation since March 2017 and the North Axis partially operating since February 2018 (Brasil, 2019Brasil. Projeto de Integração do Rio São Francisco: o andamento das obras [Internet]. Brasília: Ministério da Integração Nacional; 2019. Available from: http://www.mi.gov.br/web/projeto-sao-francisco/o-andamento-das-obras
http://www.mi.gov.br/web/projeto-sao-fra...
). Besides the SFR-IWT, the São Francisco basin was artificially connected to the upper Paraná basin in the 1960s through the rio Piumhi, during the construction of the Furnas Hydroelectric Power Plant, which resulted in an input of fish fauna from the captured basin (Moreira-Filho, Buckup, 2005Moreira-Filho O, Buckup PA. A poorly known case of watershed transposition between the São Francisco and upper Paraná river basins. Neotrop Ichthyol. 2005; 3(3):449-52. https://doi.org/10.1590/S1679-62252005000300015
https://doi.org/10.1590/S1679-6225200500...
).
The SFR-IWT installation license was granted in 2007 based on an environmental impact study (Brasil, 2004Brasil. Relatório de Impacto Ambiental-RIMA do Projeto de Integração do rio São Francisco com as Bacias Hidrográficas do Nordeste Setentrional. Brasília: Ministério da Integração Nacional/Ecology and Environment do Brasil; 2004.). This report, written at the same time as publication of the seminal studies on Caatinga fish (Rosa et al., 2003Rosa RS, Menezes NA, Britski HA, Costa WJEM, Groth F. Diversidade, padrões de distribuição e conservação dos peixes da Caatinga. In: Leal IR, Silva JMC, Tabarelli M, editors. Ecologia e Conservação da Caatinga. Recife: EDUFPE; 2003. p.135-81., 2004; Rosa, 2004Rosa RS, Gomes-Filho G, Menezes NA, Shibatta OA, Costa WJEM. Biota aquática: áreas e ações prioritárias para conservação da caatinga. In: Silva JMC, Tabarelli M, Fonseca MT, Lins LV, editors. Biodiversidade da caatinga: áreas e ações prioritárias para a conservação. Brasília: Ministério do Meio Ambiente; 2004. p.163-71.), came across taxonomic issues and species distribution inaccuracies, reflecting the incipient knowledge on this ichthyofauna. Recently, a global database of freshwater fish species listed by drainage channels included data from more than 3,000 basins; however, the Northeast of Brazil was indicated as spatial gap, with data available from few basins (Tedesco et al., 2017Tedesco PA, Beauchard O, Bigorne R, Blanchet S, Buisson L, Conti L, et al. A global database on freshwater fish species occurrence in drainage basins. Scientific Data. 2017; 4:170141. https://doi.org/10.1038/sdata.2017.141
https://doi.org/10.1038/sdata.2017.141...
). This database is useful to evaluate non-native species’ influence on the native ichthyofauna, which are under homogenization processes (Tedesco et al., 2017Tedesco PA, Beauchard O, Bigorne R, Blanchet S, Buisson L, Conti L, et al. A global database on freshwater fish species occurrence in drainage basins. Scientific Data. 2017; 4:170141. https://doi.org/10.1038/sdata.2017.141
https://doi.org/10.1038/sdata.2017.141...
). The limited information on the distributional ranges of the species also hampers evaluation of the conservation status of the species (Dias et al., 2016Dias MS, Zuanon J, Couto TBA, Carvalho M, Carvalho LN, Espirito-Santo HMV et al. Trends in studies of Brazilian stream fish assemblages. Nat Conserv. 2016; 14(2):106-11. https://doi.org/10.1016/j.ncon.2016.06.003
https://doi.org/10.1016/j.ncon.2016.06.0...
).
Due to climatic and edaphic conditions, which resulted in temporary rivers, the fish diversity of the Caatinga was underestimated until recently (Rosa et al., 2003Rosa RS, Menezes NA, Britski HA, Costa WJEM, Groth F. Diversidade, padrões de distribuição e conservação dos peixes da Caatinga. In: Leal IR, Silva JMC, Tabarelli M, editors. Ecologia e Conservação da Caatinga. Recife: EDUFPE; 2003. p.135-81.). Lima et al. (2017)Lima SMQ, Ramos TPA, da Silva MJ, Rosa RS. Diversity, distribution, and conservation of the Caatinga fishes: advances and challenges. In: Silva, JMC, Leal IR, Tabarelli M, editors. Caatinga the largest tropical dry forest region in South America. Springer; 2017. p.97-131. https://doi.org/10.1007/978-3-319-68339-3_4
https://doi.org/10.1007/978-3-319-68339-...
updated this list, increasing it to 386 species, 203 of them endemic to a single freshwater ecoregion (sensuAbell et al., 2008Abell R, Thieme ML, Revenga C, Bryer M, Kottelat M, Bogutskaya N et al. Freshwater ecoregions of the world: a new map of biogeographic units for freshwater biodiversity conservation. Bioscience. 2008; 58(5):403-14. https://doi.org/10.1641/B580507
https://doi.org/10.1641/B580507...
), and 33 threatened species. However, the biological diversity and the conservation status of its biota are still poorly known, and inventories should be performed on a regional scale to support public conservation policies (Rosa et al., 2004Rosa RS, Gomes-Filho G, Menezes NA, Shibatta OA, Costa WJEM. Biota aquática: áreas e ações prioritárias para conservação da caatinga. In: Silva JMC, Tabarelli M, Fonseca MT, Lins LV, editors. Biodiversidade da caatinga: áreas e ações prioritárias para a conservação. Brasília: Ministério do Meio Ambiente; 2004. p.163-71.; Lévêque et al., 2005Lévêque C, Balian EV, Martens K. An assessment of animal species diversity in continental waters. Hydrobiologia. 2005; 542:39-67. https://doi.org/10.1007/s10750-004-5522-7
https://doi.org/10.1007/s10750-004-5522-...
, 2008; Tedesco et al., 2017Tedesco PA, Beauchard O, Bigorne R, Blanchet S, Buisson L, Conti L, et al. A global database on freshwater fish species occurrence in drainage basins. Scientific Data. 2017; 4:170141. https://doi.org/10.1038/sdata.2017.141
https://doi.org/10.1038/sdata.2017.141...
). The Caatinga’s hydrography is composed of four freshwater ecoregions: Maranhão-Piauí (MAPE), Mid-Northeastern Caatinga (MNCE), São Francisco (SFRE), and Northeastern Atlantic Forest (NAFE) (Rosa et al., 2003Rosa RS, Menezes NA, Britski HA, Costa WJEM, Groth F. Diversidade, padrões de distribuição e conservação dos peixes da Caatinga. In: Leal IR, Silva JMC, Tabarelli M, editors. Ecologia e Conservação da Caatinga. Recife: EDUFPE; 2003. p.135-81.; Albert et al., 2011Albert JS, Petry P, Reis RE. Major biogeographic and phylogenetic patterns. In: Albert JS, Reis RE, editors. Historical biogeography of Neotropical freshwater fishes. Berkeley and Los Angeles, California: University of California Press; 2011. p.21-57.; Lima et al., 2017Lima SMQ, Ramos TPA, da Silva MJ, Rosa RS. Diversity, distribution, and conservation of the Caatinga fishes: advances and challenges. In: Silva, JMC, Leal IR, Tabarelli M, editors. Caatinga the largest tropical dry forest region in South America. Springer; 2017. p.97-131. https://doi.org/10.1007/978-3-319-68339-3_4
https://doi.org/10.1007/978-3-319-68339-...
). Most MNCE basins exhibit an intermittent regime and small to medium extensions (Rosa et al., 2004Rosa RS, Gomes-Filho G, Menezes NA, Shibatta OA, Costa WJEM. Biota aquática: áreas e ações prioritárias para conservação da caatinga. In: Silva JMC, Tabarelli M, Fonseca MT, Lins LV, editors. Biodiversidade da caatinga: áreas e ações prioritárias para a conservação. Brasília: Ministério do Meio Ambiente; 2004. p.163-71.). They also have lower fish species richness and endemism (88 and 38, respectively) when compared to the adjacent SFRE (211 and 135) and to the MAPE (151 and 54) (Albert et al., 2011Albert JS, Petry P, Reis RE. Major biogeographic and phylogenetic patterns. In: Albert JS, Reis RE, editors. Historical biogeography of Neotropical freshwater fishes. Berkeley and Los Angeles, California: University of California Press; 2011. p.21-57.; Ramos et al., 2014Ramos TPA, Lima JAS, Costa SYL, Silva MJ, Avellar RC, Oliveira-Silva L. Continental ichthyofauna from the Paraíba do Norte River basin pre-transposition of the São Francisco River, Northeastern Brazil. Biota Neotrop. 2018; 18(4):e20170471. https://doi.org/10.1590/1676-0611-bn-2017-0471
https://doi.org/10.1590/1676-0611-bn-201...
; Silva et al., 2015Silva MJ, Costa BG, Ramos TPA, Auricchio P, Lima SMQ. Ichthyofauna of the Gurgueia river, Parnaíba river basin, Northeastern Brazil. Check List. 2015; 11(5):1-8. https://doi.org/10.15560/11.5.1765
https://doi.org/10.15560/11.5.1765...
; Reis et al., 2016Ramos TPA, Ramos RTC, Ramos SAQA. Ichthyofauna of the Parnaíba river basin, Northeastern Brazil. Biota Neotrop. 2014; 14(1):1-8. https://doi.org/10.1590/S1676-06020140039
https://doi.org/10.1590/S1676-0602014003...
). The high endemism level of these ecoregions suggests ancient separation and low connectivity among the basins (Albert, Carvalho, 2011Albert JS, Carvalho TP. Neogene assembly of modern faunas. In: Albert JS, Reis RE, editors. Historical biogeography of Neotropical freshwater fishes. Berkeley and Los Angeles, California: University of California Press; 2011. p.119-36.), but these numbers are still partial, especially for the MNCE, since the regional collections are scarcely digitalized, and only a few species of this ecoregion, are available (Buckup et al., 2007Buckup PA, Menezes NA, Ghazzi MS. Catálogo das Espécies de Peixes de Água Doce do Brasil. Rio de Janeiro: Museu Nacional; 2007. ; Lévêque et al., 2008Lévêque C, Oberdorff T, Paugy D, Stiassny MLJ, Tedesco PA. Global diversity of fish (Pisces) in freshwater. Hydrobiologia. 2008; 595:545-67. https://doi.org/10.1007/978-1-4020-8259-7_53
https://doi.org/10.1007/978-1-4020-8259-...
; Langeani et al., 2009Langeani F, Buckup PA, Malabarba LR, Py-Daniel LHR, Lucena CAS, Rosa RS et al. Peixes de água doce. In: Rocha RM, Boeger WA, editors. Estado da arte e perspectivas para a Zoologia no Brasil. Curitiba: EDUFPR; 2009. p.211-30.).
Regarding the SFR-IWT basins, there is a huge data deficiency, and comparative studies are paramount to inform which species are endemic, naturally shared and potentially invasive before the artificial connection (Langeani et al., 2009Langeani F, Buckup PA, Malabarba LR, Py-Daniel LHR, Lucena CAS, Rosa RS et al. Peixes de água doce. In: Rocha RM, Boeger WA, editors. Estado da arte e perspectivas para a Zoologia no Brasil. Curitiba: EDUFPR; 2009. p.211-30.). This urgency encouraged the present comprehensive study of the SFR-IWT’s freshwater fish fauna. Therefore, the purpose of this study was to elaborate an original ichthyofaunal species list of the basins encompassed in the SFR-IWT, proposing a taxonomic nomenclatural standardization, and indicating which species are endemic, endangered, non-native, and naturally shared between basins and ecoregions. Notwithstanding, this baseline is not definitive and should be refined and updated regularly. Besides that, it may help in the detection of possible changes in fish composition resulting from the SFR-IWT and the proposition of public policies for the conservation of Caatinga ichthyofauna.
MATERIAL AND METHODS
Information source. The freshwater fish species occurrence data comprised the rio São Francisco lower-middle stretch, the SFR-IWT catchment area in SFRE; and receptor’s drainages of the Jaguaribe (JAG), Apodi-Mossoró (APO), Piranhas-Açu (PIA), and Paraíba do Norte (PAR) river basins in the MNCE. The map was produced with QuantumGIS free program (www.qgis.org). Calculations of the drainage area and number of municipalities were done using georeferenced shapefiles of the National Water Agency (Brasil, 2014aBrasil. Metadados [Internet]. Agência Nacional de Águas; 2014a. Available from: http://metadados.ana.gov.br/geonetwork/srv/pt/main.home
http://metadados.ana.gov.br/geonetwork/s...
) and Ministry of the Environment (Brasil, 2014bBrasil. Dados georreferenciados [Internet]. Ministério do Meio Ambiente; 2014b. Available from: http://www.mma.gov.br/areas-protegidas/cadastro-nacional-de-ucs/dados-georreferenciados
http://www.mma.gov.br/areas-protegidas/c...
), while the São Francisco Hydrographic Basin Committee provided the SFR-IWT canal trails (CBHSF, 2014CBHSF (Comitê da Bacia Hidrográfica do Rio São Francisco). Shapefiles [Internet]. CBHSF; 2014. Available from: http://www.saofrancisco.cbh.gov.br/Shapes/SHAPE.sigeo.Transposicao.SF.zip
http://www.saofrancisco.cbh.gov.br/Shape...
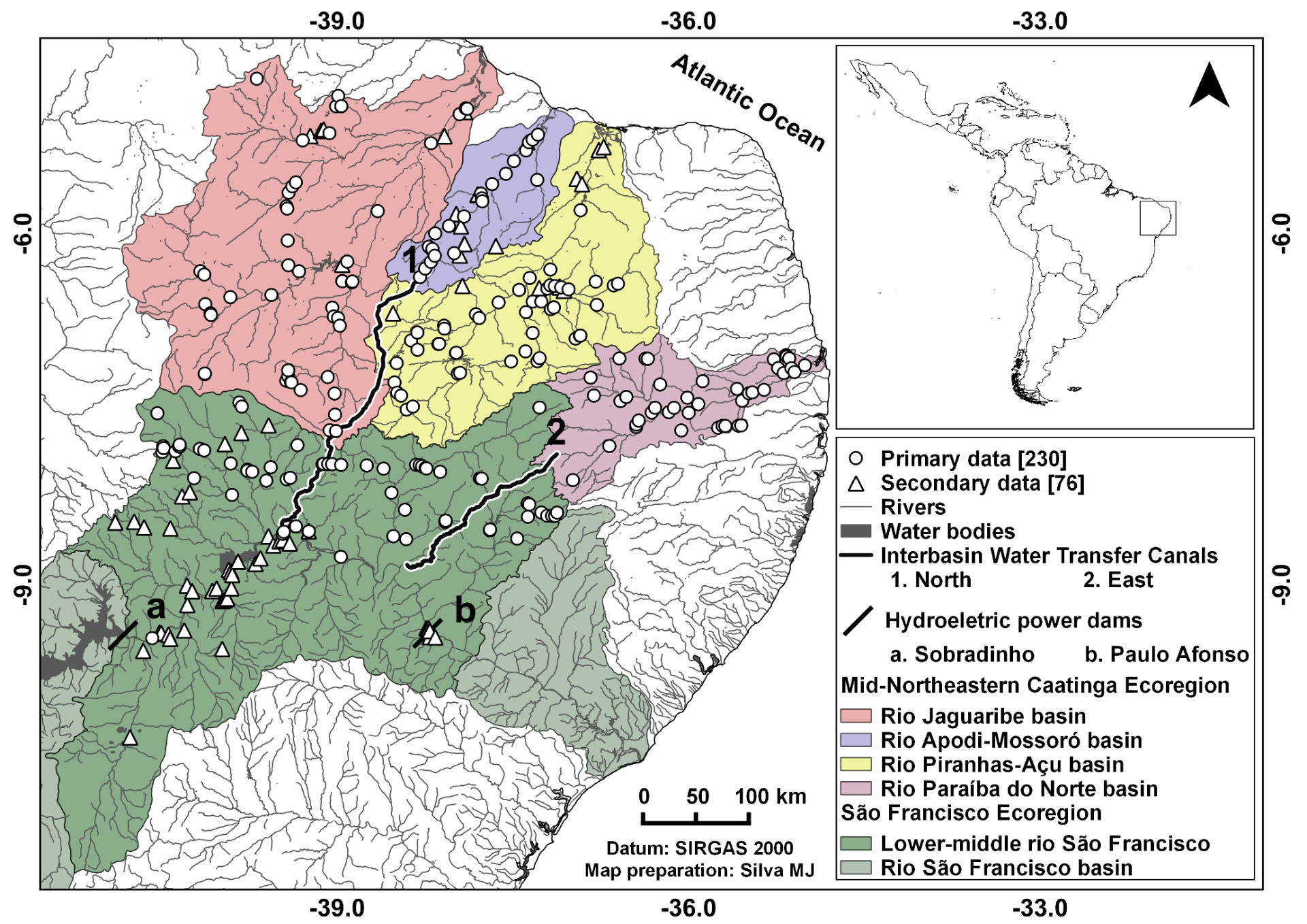
) (Fig. 1).

Sampling sites of the freshwater fish species in the São Francisco Interbasin Water Transfer Project basins in the Brazilian semiarid.
The São Francisco basin and ecoregion has an approximate area of 640,000 km². Its main river, the São Francisco, is the largest in the Northeast region, and the third in Brazil (Rosa et al., 2003Rosa RS, Menezes NA, Britski HA, Costa WJEM, Groth F. Diversidade, padrões de distribuição e conservação dos peixes da Caatinga. In: Leal IR, Silva JMC, Tabarelli M, editors. Ecologia e Conservação da Caatinga. Recife: EDUFPE; 2003. p.135-81.). It is subdivided in to four stretches: upper, middle, lower-middle, and lower (Brasil, 2014aBrasil. Metadados [Internet]. Agência Nacional de Águas; 2014a. Available from: http://metadados.ana.gov.br/geonetwork/srv/pt/main.home
http://metadados.ana.gov.br/geonetwork/s...
). Herein, only records in the lower-middle stretch (112,093.68 km², 17.51% of São Francisco basin and 42.52% of SFR-IWT) were considered, and this stretch is under direct influence of the SFR-IWT, including 98 municipalities, from downstream Sobradinho Dam reservoir (Sobradinho Municipality, Bahia State) to the upstream Lajeadinho Stream (Canindé de São Francisco Municipality, Sergipe State), or approximately 50 km downstream of Paulo Afonso Dam (Brasil, 2014aBrasil. Metadados [Internet]. Agência Nacional de Águas; 2014a. Available from: http://metadados.ana.gov.br/geonetwork/srv/pt/main.home
http://metadados.ana.gov.br/geonetwork/s...
) (Fig. 1). In this semiarid area, most tributaries of the São Francisco are intermittent (Rosa et al., 2003Rosa RS, Menezes NA, Britski HA, Costa WJEM, Groth F. Diversidade, padrões de distribuição e conservação dos peixes da Caatinga. In: Leal IR, Silva JMC, Tabarelli M, editors. Ecologia e Conservação da Caatinga. Recife: EDUFPE; 2003. p.135-81.), and some of them (Moxotó, Pajeú, Terra Nova, and Brígida rivers) will also receive water inflow from the main São Francisco channel (Lima, 2005Lima LC. Além das águas, a discussão no nordeste do rio São Francisco. Revista do Departamento de Geografia. 2005; 17:94-100. https://doi.org/10.7154/RDG.2005.0017.0007
https://doi.org/10.7154/RDG.2005.0017.00...
).
The receptor basins are the four main drainage areas of the Mid-Northeatern Caatinga ecoregion (MNCE), presented in the west-east direction. The Jaguaribe basin supplies water to 88 municipalities of Ceará State and is the largest MNCE basin, with an area of 74,077.01 km² (28.10% of SFR-IWT). Besides the Jaguaribe River (633 km of extension), the basin includes tributaries, such as the Banabuiú and Salgado rivers (Brasil, 2014aBrasil. Metadados [Internet]. Agência Nacional de Águas; 2014a. Available from: http://metadados.ana.gov.br/geonetwork/srv/pt/main.home
http://metadados.ana.gov.br/geonetwork/s...
). The Apodi-Mossoró basin encompasses 68 municipalities in Rio Grande do Norte State with an area of 14,303.71 km² (5.43% of SFR-IWT), and approximately 200 km of extension (Brasil, 2014aBrasil. Metadados [Internet]. Agência Nacional de Águas; 2014a. Available from: http://metadados.ana.gov.br/geonetwork/srv/pt/main.home
http://metadados.ana.gov.br/geonetwork/s...
). Extending about 350 km, the Piranhas-Açu basin represents the second largest MNCE basin (43,141.54 km², 16.37% of SFR-IWT), draining 158 municipalities in Paraíba and Rio Grande do Norte states (Brasil, 2014aBrasil. Metadados [Internet]. Agência Nacional de Águas; 2014a. Available from: http://metadados.ana.gov.br/geonetwork/srv/pt/main.home
http://metadados.ana.gov.br/geonetwork/s...
). These three basins will receive water from the SFR-IWT North Axis (Brasil, 2004Brasil. Relatório de Impacto Ambiental-RIMA do Projeto de Integração do rio São Francisco com as Bacias Hidrográficas do Nordeste Setentrional. Brasília: Ministério da Integração Nacional/Ecology and Environment do Brasil; 2004.) (Fig. 1). Lastly, the Paraíba do Norte basin, with 19,977.48 km² (7.58% of SFR-IWT) and 280 km length in 90 municipalities (Brasil, 2014aBrasil. Metadados [Internet]. Agência Nacional de Águas; 2014a. Available from: http://metadados.ana.gov.br/geonetwork/srv/pt/main.home
http://metadados.ana.gov.br/geonetwork/s...
) in Paraíba State, is the largest one in the MNCE that flows to the east coast (the other ones drain to the north) (Brasil, 2019Brasil. Projeto de Integração do Rio São Francisco: o andamento das obras [Internet]. Brasília: Ministério da Integração Nacional; 2019. Available from: http://www.mi.gov.br/web/projeto-sao-francisco/o-andamento-das-obras
http://www.mi.gov.br/web/projeto-sao-fra...
) (Fig. 1).
Species records. Species occurrences were obtained through latitude and longitude data, taking into account strictly freshwater fish families (Buckup et al., 2007Buckup PA, Menezes NA, Ghazzi MS. Catálogo das Espécies de Peixes de Água Doce do Brasil. Rio de Janeiro: Museu Nacional; 2007. ). In the few cases in which the geographical coordinates were not available in the original source, they were approximated using the GeoNames website (www.geonames.org). All records were made before any water input from the SFR-IWT canals in March 2017.
Primary data were obtained from collected and deposited material in ichthyological collections at the Federal Universities of Paraíba (UFPB, 129 localities - 56.09%) and Rio Grande do Norte (UFRN, 101 - 43.91%). All locality data were listed in the supplementary material (S1). Distinct active (trawls, dip nets, sieves, fishhooks, and cast nets) and passive (gill nets and traps) fishing methods were used to reduce selectivity (Uieda, Castro, 199922 Uieda VS, Castro RMC. Coleta e fixação de peixes de riachos. In: Caramaschi EP, Mazzoni R, Peres-Neto PR, editors. Ecologia de Peixes de Riachos. Rio de Janeiro: Oecologia Brasiliensis; 1999. p.22.).
Secondary data (76 localities) were acquired in two steps: 1) data from digitalized ichthyological collections available in speciesLink, PRONEX/NEODAT, and Portal da Biodiversidade (www.splink.org.br, www.mnrj.ufrj.br/pronex/, and https://portaldabiodiversidade.icmbio.gov.br/portal/), and 2) literature review (e.g.Reis et al., 2003Reis RE, Kullander SO, Ferraris-Jr CJ, editors. Check list of the freshwater fishes of South and Central America. Porto Alegre: EDIPUCRS; 2003.; Buckup et al., 2007Buckup PA, Menezes NA, Ghazzi MS. Catálogo das Espécies de Peixes de Água Doce do Brasil. Rio de Janeiro: Museu Nacional; 2007. ). At this stage, the bibliographic databases Web of Science and Google Scholar were consulted during searches of species records in the SFR-IWT basins (filtering dubious occurrences) after January 2006, the time frame used by Buckup et al. (2007)Buckup PA, Menezes NA, Ghazzi MS. Catálogo das Espécies de Peixes de Água Doce do Brasil. Rio de Janeiro: Museu Nacional; 2007. , until March 2017, current frame (e.g.Luz et al., 2012Luz SCS, Lima HC, Severi W. Composição da ictiofauna em ambientes marginais e tributários do médio-submédio rio São Francisco. Braz J Agricult Sci. 2012; 7(2):358-66. https://doi.org/10.5039/agraria.v7i2a1436
https://doi.org/10.5039/agraria.v7i2a143...
; Nascimento et al., 2014Nascimento WS, Barros NHC, Araújo AS, Gurgel LL, Canan B, Molina WF, et al. Composição da ictiofauna das bacias hidrográficas do Rio Grande do Norte, Brasil. Biota Amazônia. 2014; 4(1):126-31. https://doi.org/10.18561/2179-5746/biotaamazonia.v4n1p126-131
https://doi.org/10.18561/2179-5746/biota...
), including new species descriptions (e.g.Ribeiro, Lucena, 2006Ribeiro FRV, Lucena CAS. A new species of Pimelodus LaCépède, 1803 (Siluriformes: Pimelodidae) from the rio São Francisco drainage, Brazil. Neotrop Ichthyol. 2006; 4(4):411-8. https://doi.org/10.1590/S1679-62252006000400004
https://doi.org/10.1590/S1679-6225200600...
; Lucena, 2007Lucena CAS. Revisão taxonômica das espécies do gênero Roeboides grupo-affinis (Ostariophysi, Characiformes, Characidae). Iheringia Ser Zool. 2007; 97(2):117-36. https://doi.org/10.1590/S0073-47212007000200001
https://doi.org/10.1590/S0073-4721200700...
; Lima, Britski, 2007Lima FCT, Britski HA. Salminus franciscanus, a new species from the Rio São Francisco basin, Brazil (Ostariophysi: Characiformes: Characidae). Neotrop Ichthyol. 2007; 5(3):237-44. https://doi.org/10.1590/S1679-62252007000300001
https://doi.org/10.1590/S1679-6225200700...
; Ramos et al., 2013Ramos TPA, Barros-Neto LF, Britski HA, Lima SMQ. Parotocinclus seridoensis, a new hypoptopomatine catfish (Siluriformes: Loricariidae) from the upper rio Piranhas-Açu basin, northeastern Brazil. Neotrop Ichthyol. 2013; 11(4):787-96. https://doi.org/10.1590/S1679-62252013000400006
https://doi.org/10.1590/S1679-6225201300...
; Zawadzki et al., 2017Zawadzki CH, Ramos TPA, Sabaj M. Hypostomus sertanejo (Siluriformes: Loricariidae), new armoured catfish species from north-eastern Brazil. J Fish Biol. 2017; 91(1):317-30. https://doi.org/10.1111/jfb.13349
https://doi.org/10.1111/jfb.13349...
). The abbreviation “aff.” was used for species that have affinity with, but should be distinct from, the nominal taxon to which they are currently assigned.
Taxonomic validation. Nominal species validity, synonyms, and systematic classification were carried out according to Fricke et al. (2020)Fricke R, Eschmeyer WN, Van-Der-Laan R, editors. Eschmeyer’s Catalog of Fishes: genera, species, references [Internet]. San Francisco: California Academy of Science; 2020. Available from: https://www.calacademy.org/scientists/projects/catalog-of-fishes
https://www.calacademy.org/scientists/pr...
. Some species with uncertain taxonomic status were considered as species inquirendae, following the recommendation of Lima et al. (2017)Lima SMQ, Ramos TPA, da Silva MJ, Rosa RS. Diversity, distribution, and conservation of the Caatinga fishes: advances and challenges. In: Silva, JMC, Leal IR, Tabarelli M, editors. Caatinga the largest tropical dry forest region in South America. Springer; 2017. p.97-131. https://doi.org/10.1007/978-3-319-68339-3_4
https://doi.org/10.1007/978-3-319-68339-...
. Specific studies on species introduction in the Northeast region were examined in order to determine non-native species (Gurgel, Fernando, 1994Gurgel JJS, Fernando CH. Fisheries in Semi-Arid Northeast Brazil with special reference to the role of Tilapias. Int Rev Hydrobiol. 1994; 79(1):77-94. https://doi.org/10.1002/iroh.19940790109
https://doi.org/10.1002/iroh.19940790109...
; DNOCS, 2002DNOCS (Departamento Nacional de Obras Contra a Seca). Relatório das atividades desenvolvidas pela coordenação de pesca e aquicultura, durante o ano de 2002. Coordenação de Pesca e Aquicultura; 2002.; Rosa et al., 2003Rosa RS, Menezes NA, Britski HA, Costa WJEM, Groth F. Diversidade, padrões de distribuição e conservação dos peixes da Caatinga. In: Leal IR, Silva JMC, Tabarelli M, editors. Ecologia e Conservação da Caatinga. Recife: EDUFPE; 2003. p.135-81.; Mattheus, 2005Mattheus S. América do Sul invadida: a crescente ameaça das espécies exóticas invasoras. Programa Global de Espécies Invasoras; 2005.; Leão et al., 2011Leão TCC, Almeida WR, Dechoum M, Ziller SR. Espécies exóticas invasoras no nordeste do Brasil: contextualização, manejo e políticas públicas. Recife: CEPAN; 2011.; Paiva, Mesquita, 2013Paiva MP, Mesquita PEC. Uma semente fecunda: commissão téchnica de piscicultura do Nordeste (1932-1945). Revista do Instituto do Ceará. 2013; 127:9-40.). The Brazilian list of endangered fish and aquatic invertebrates was consulted to indicate the threatened ones (Brasil, 2014cBrasil. Portaria Ministério do Meio Ambiente n° 445, de 17 de Dezembro de 2014. Lista Nacional Oficial de Espécies da Fauna Ameaçada de Extinção: Peixes e Invertebrados Aquáticos. Brasília: Diário Oficial da União; 2014c. p.126-30), given that it follows IUCN parameters. Reis et al. (2003)Reis RE, Kullander SO, Ferraris-Jr CJ, editors. Check list of the freshwater fishes of South and Central America. Porto Alegre: EDIPUCRS; 2003., Buckup et al. (2007)Buckup PA, Menezes NA, Ghazzi MS. Catálogo das Espécies de Peixes de Água Doce do Brasil. Rio de Janeiro: Museu Nacional; 2007. , Fricke et al. (2020)Fricke R, Eschmeyer WN, Van-Der-Laan R, editors. Eschmeyer’s Catalog of Fishes: genera, species, references [Internet]. San Francisco: California Academy of Science; 2020. Available from: https://www.calacademy.org/scientists/projects/catalog-of-fishes
https://www.calacademy.org/scientists/pr...
and Lima et al. (2017)Lima SMQ, Ramos TPA, da Silva MJ, Rosa RS. Diversity, distribution, and conservation of the Caatinga fishes: advances and challenges. In: Silva, JMC, Leal IR, Tabarelli M, editors. Caatinga the largest tropical dry forest region in South America. Springer; 2017. p.97-131. https://doi.org/10.1007/978-3-319-68339-3_4
https://doi.org/10.1007/978-3-319-68339-...
were examined in order to determine the endemic species, defined herein as those restricted to a single basin or hydrographic ecoregion.
All specimens were deposited in one of these museums: ANSP (The Academy of Natural Sciences, Drexel University); FMNH (Field Museum of Natural History); LIRP (Laboratório de Ictiologia de Ribeirão Preto); MCP (Museu de Ciências e Tecnologia da Pontifícia Universidade Católica do Rio Grande do Sul); MNHN (Muséum National d’Histoire Naturelle); MNRJ (Museu Nacional, Universidade Federal do Rio de Janeiro); MZUEL (Museu de Zoologia, Universidade Estadual de Londrina); MZUSP (Museu de Zoologia, Universidade de São Paulo); NMW (Naturhistorisches Museum, Wien); NUPELIA (Núcleo de Pesquisa em Limnologia, Ictiologia e Aquicultura); UFPB (Universidade Federal da Paraíba); UFRN (Universidade Federal do Rio Grande do Norte); UMMZ (University of Michigan Museum of Zoology); ZMB (Museum für Naturkunde). All lots are listed in the supplementary material (S2).
Spatial patterns of species richness. All species inquirendae, and those of the secondary and vicarious divisions families (e.g. Sciaenidae, Gobiidae, Clupeidae, Engraulidae) (sensuMyers, 1937Myers GS. Fresh-water fishes and west Indian zoogeography. Annu Rep Board Regents Smithson Inst. 1937; 3(465):339-64.) were removed from comparisons to avoid inflating the number of shared species due to taxonomic uncertainty or possible marine dispersion. However, the distribution of these species in literature was presented in S3 (available only in the online version) for future research. For the evaluation of spatial patterns of richness, each hydrographic basin had its species-area relationship calculated for native and endemic species (Albert et al., 2011Albert JS, Petry P, Reis RE. Major biogeographic and phylogenetic patterns. In: Albert JS, Reis RE, editors. Historical biogeography of Neotropical freshwater fishes. Berkeley and Los Angeles, California: University of California Press; 2011. p.21-57.). This relationship is extensively used as a way of comparing distinct and size-diverse ecosystems worldwide (Arrhenius, 1921Arrhenius O. Species and area. J Ecol. 1921; 9:95-99. https://doi.org/10.2307/2255763
https://doi.org/10.2307/2255763...
; Connor, McCoy, 1979Connor EF, McCoy ED. The statistics and biology of the species area relationship. Am Nat. 1979; 113:791-833. Available from: http://www.jstor.org/stable/2460305
http://www.jstor.org/stable/2460305...
). Thus, in each region, the number of species is correlated with their spatial extent, through a function of type S=S0Ab. Where S is the number of species in area A, S0 is proportional species density (i.e., species richness per unit area), and b is the species-area scaling exponent, often with values in the range 0.25-0.50 (Dengler, 2009Dengler J. Which function describes the species-area relationship best? A review and empirical evaluation. J Biogeogr. 2009; 36(4):728-44. https://doi.org/10.1111/j.1365-2699.2008.02038.x
https://doi.org/10.1111/j.1365-2699.2008...
). Since the scaling exponent b defines the slope of the species-area regression, it may be interpreted as a measure of gamma diversity among areas, with higher values indicating greater differences in the taxonomic composition of areas. The similarities of native species composition between river basins were calculated using Jaccard’s similarity coefficient index, using the following formula J=(c/(a+b+c)).100, where c is the number of fish species existing at both sampling sites and a and b are the number of fish species at different sampling sites. This index takes into account only the values of presence and absence of species in each basin (Lévêque et al., 2013Li J, Dong S, Peng M, Yang Z, Liu S, Li X et al. Effects of damming on the biological integrity of fish assemblages in the middle Lancang-Mekong River basin. Ecol Indic. 2013; 34:94-102. https://doi.org/10.1016/j.ecolind.2013.04.016
https://doi.org/10.1016/j.ecolind.2013.0...
).
RESULTS
Species richness and species-area relationships. In total, 17,002 lots of fish were compiled, of which 3,331 (2,729 primary data) were selected and recorded in 306 sampling sites (230 primaries), 117 (66 primaries) in SFRE (lower-middle São Francisco basin) and 189 (164 primaries) in the MNCE. In the receptor drainages (MNCE’s localities), there were 56 (50 primaries) in the Jaguaribe (JAG), 30 (22) in the Apodi-Mossoró (APO), 59 (all primaries) in the Piranhas-Açu (PIA), and 44 (all primaries) in the Paraíba do Norte basins (PAR) (Figs. 1-2).

Sampling sites in the São Francisco Interbasin Water Transfer Project basins in the Brazilian semiarid. A = Canals under construction near the rio São Francisco main channel, and B = near Sertânia, Pernambuco State (PE), C = Rio Pajeú, tributary of the São Francisco basin, PE, D = Rio São Francisco near Petrolina, PE, E = Temporary pool in rio Jaguaribe basin in Russas, Ceará State (CE), F = Rio Jaguaribe in Crato, CE, G = Rio Apodi-Mossoró in Pau dos Ferros, Rio Grande do Norte State (RN), H = Rio Apodi-Mossoró in Pau dos Ferros, RN, I = Rio Seridó, tributary of the Piranhas-Açu basin in Caicó, RN, J = Rio Piranhas-Açu, Jardim de Piranhas, RN, K = Rio Paraíba do Norte in Barra de Santana, Paraíba State (PB), L = Rio Paraíba do Norte in São João do Cariri, PB.
In this study, 121 valid freshwater fish species were recorded, from 25 families and seven orders, or 111, 23 and six, respectively, considering only the native species (Tab. 1, Fig. 3A-I). Additionally, five species belonging to families of the vicarious divisions were found, but disregarded in the list and analyses: three native, Anchoviella vaillanti (Steindachner, 1908) endemic to SFRE, A. lepidentostole (Fowler, 1911) present in JAG, and Awaous tajasica (Lichtenstein, 1822) recorded in PAR, and two introduced ones, Plagioscion auratus (Castelnau, 1855) in SFRE and P. squamosissimus (Heckel, 1840), recorded in all basins of the SFR-IWT, except in PAR (Alves et al., 2011Alves CBM, Vieira F, Pompeu PS. Ictiofauna da bacia hidrográfica do rio São Francisco. Diagnóstico do macrozoneamento ecológico-econômico da Bacia Hidrográfica do Rio São Francisco. Brasília: Ministério do Meio Ambiente; 2011. p.226-41.; Leão et al., 2011Leão TCC, Almeida WR, Dechoum M, Ziller SR. Espécies exóticas invasoras no nordeste do Brasil: contextualização, manejo e políticas públicas. Recife: CEPAN; 2011.). Furthermore, six species inquirendae were listed, Psalidodon rivularis (Lütken, 1875), Pseudancistrus papariae Fowler, 1941, Pimelodella papariae (Fowler, 1941), P. witmeri Fowler, 1941, Hypostomus carvalhoi (Miranda Ribeiro, 1937), and H. jaguribensis (Fowler, 1915) (S3).
Five endangered species were recorded, three of them in SFRE, Lophiosilurus alexandri Steindachner, 1876 as vulnerable, Conorhynchos conirostris (Valenciennes, 1840) as endangered (EN), and Hypsolebias flavicaudatus (Costa & Brasil, 1990) as critically endangered), and two in MNCE, Apareiodon davisi Fowler, 1941 (EN) and Parotocinclus spilurus (Fowler, 1941) (EN) (Tab. 1 , Fig. 3F). Eleven non-native species were recorded, eight of which occur in SFRE and MNCE receptor basins (Colossoma macropomum, Parachromis managuensis, Poecilia reticulata, Astronotus ocellatus, Cichla monoculus, C. kelberi, Oreochromis niloticus, and Coptodon rendalli); the last six exist in all SFR-IWT basins, and three only in the MNCE (Arapaima gigas, Megaleporinus obtusidens, native in SFRE, and Xiphophorus helleri) (Tab. 1, Fig. 3J-L).
List of freshwater fish species recorded in the São Francisco Interbasin Water Transfer Project basins in the Brazilian semiarid. SFRE = São Francisco Ecoregion (lower-middle São Francisco basin); MNCE = Mid-Northeastern Caatinga Ecoregion; JAG = Jaguaribe basin; APO = Apodi-Mossoró basin; PIA = Piranhas-Açu basin; PAR = Paraíba do Norte basin; P = Primary data; S = Secondary data; NNA = Non-Native Species; EN = Endangered; VU = Vulnerable; CR = Critically Endangered. Fish classification follows Fricke et al. (2020)Fricke R, Eschmeyer WN, Van-Der-Laan R, editors. Eschmeyer’s Catalog of Fishes: genera, species, references [Internet]. San Francisco: California Academy of Science; 2020. Available from: https://www.calacademy.org/scientists/projects/catalog-of-fishes
https://www.calacademy.org/scientists/pr... .

Freshwater fish species from the São Francisco Interbasin Water Transfer Project basins in the Brazilian semiarid. A = Hemigrammus brevis, endemic species of São Francisco Ecoregion (SFRE); B = Moenkhausia costae and C = Psellogrammus kennedyi, shared species between SFRE and Mid-Northeastern Caatinga Ecoregion (MNCE); D = Aspidoras menezesi, endemic species of Jaguaribe basin (JAG) ; E = Hypostomus sertanejo, endemic species of MNCE; F = Parotocinclus spilurus, endemic and endangered species of JAG; G = Tatia bockmanni, endemic species of SFRE; H = Cichlasoma orientale, shared species from all basins of SFR-IWT; I = Geophagus brasiliensis, shared species between SFRE and MNCE; J = Colossoma macropomum, non-native species shared between SFRE and Piranhas-Açu basin; K = Parachromis managuensis, non-native species shared between SFRE and Paraíba do Norte basin; L = Xiphophorus helleri, non-native species of JAG.
Regarding the hydrographic basins, considering only native species, the highest richness (78, corresponding to 70.27%) was recorded in the SFRE donor basin, while the MNCE’s receptor basins accounted for 61 species (54.95%). Of those, 50 species (44.05% of total and 81.97% of MNCE) were recorded in JAG, 39 (35.14% and 63.93%) in PIA, 36 (32.43% and 59.02%) in PAR, and 32 (28.83% and 52.46%) in APO (Tab. 2). The species-area relationship calculation indicated that the SFRE (lower-middle stretch) exhibits the highest native species density (1.75), followed by PAR (1.42), APO (1.41), JAG (1.29), MNCE (1.24, all receptor basins) and PIA (1.20) (Tab. 3). Considering only the endemic species, the following values were obtained: SFRE (0.52), MNCE (0.26), JAG (0.05), PAR (0.04), and PIA (0.03). The APO basin did not contain any endemic species, and its higher species-area value might be related to its smaller size in comparison to the other basins (Tab. 3).
Number and percentage of shared native fish species in the São Francisco Interbasin Water Transfer Project basins in the Brazilian semiarid. Number of species in each basin and ecoregion on the diagonal in bold. SFRE = São Francisco Ecoregion (lower-middle São Francisco basin); MNCE = Mid-Northeastern Caatinga Ecoregion (JAG+APO+PIA+PAR); JAG = Jaguaribe basin; APO = Apodi-Mossoró basin; PIA = Piranhas-Açu basin; PAR = Paraíba do Norte basin.
Species-area index of the native freshwater fish species in the São Francisco Interbasin Water Transfer Project basins in the Brazilian semiarid. A = Area (km2); ST = Total Species; %A = Proportion of Area; %ST = Proportion of Species; C=ST/Ab = Species Density; SE = Endemic Species; %SE = Proportion of Endemic species; E=SE/Ab = Endemic Species Density; SFRE = São Francisco Ecoregion (lower-middle São Francisco basin); MNCE = Mid-Northeastern Caatinga Ecoregion (JAG+APO+PIA+PAR); JAG = Jaguaribe basin; APO = Apodi-Mossoró basin; PIA = Piranhas-Açu basin; PAR = Paraíba do Norte basin; b* = the species-area scaling work (0.3264).
Spatial distribution and species sharing. Among the 111 native freshwater fish species recorded, 16 are present in all SFR-IWT basins (Prochilodus brevis, Leporinus piau, Hoplias aff. malabaricus, Compsura heterura, Hemigrammus marginatus, Psalidodon fasciatus, Serrapinnus heterodon, S. piaba, Triportheus signatus, Trachelyopterus galeatus, Rhamdia aff. quelen, Hypostomus pusarum, Poecilia vivipara, Synbranchus aff. marmoratus, Cichlasoma orientale and Crenicichla brasiliensis) (Tab. 1, Fig. 3H). The ichthyofaunal composition similarity between SFRE and MNCE was composed of 28 (25.23%) shared species. Furthermore, a pairwise comparison of the sharing values of SFRE with the four receptor basins of MNCE indicated that the similarity species composition was even lower: between 20 (22.22%, SFRE x APO) and 24 (23.08%, SFRE x JAG). As expected, the percentage of shared species was higher between the receptor basins of the MNCE, varying from 41.67% (20 spp., APO x PAR) to 64.81%, (35, JAG x PIA). It is worth mentioning that 49 (62.82%) of the SFRE native species do not occur in any receptor basin, and 33 (54.10%) species occur in the MNCE basin but are absent in SFRE (Tab. 1).
There are also some systematic differences in the ichthyofaunal composition of the SFR-IWT ecoregions, mainly referring to the families Loricariidae, Anostomidae, Pimelodidae, and Characidae. Only three loricariid species occur in both ecoregions (Hypostomus pusarum, Parotocinclus cearensis, and P. jumbo), while three are only found in SFRE (Hypostomus macrops, Pterygoplichthys etentaculatus, and Rineloricaria sp.), and seven only in MNCE (Hypostomus sertanejo, Loricariichthys derbyi, Loricariichthys sp., Parotocinclus seridoensis, P. spilosoma, P. spilurus, and Pseudancistrus genisetiger) (Fig. 3E-F). Only two Anostomidae species were recorded in both ecoregions (Leporinus piau and L. taeniatus), while five are exclusive to SFRE (Leporellus vittatus, Leporinus friderici, Megaleporinus obtusidens, M. reinhardti, and Schizodon knerii), and one to MNCE (Schizodon dissimilis). All pimelodid species (five) were only recorded in SFRE, besides Conorhynchos conirostris, previously allocated in this family, and also endemic to the SFRE. Lastly, of the 26 characid species, only seven were found simultaneously in SFRE and MNCE (Compsura heterura, Hemigrammus marginatus, Moenkhausia costae, Psalidodon fasciatus, Psellogrammus kennedyi, Serrapinnus heterodon, and S. piaba), while 11 were recorded only in SFRE, and eight in MNCE (Tab. 1, Fig. 3B-C).
Endemic species. Among the 111 native freshwater fish species recorded in the basins of the SFR-IWT, 36 (32.43%) species were considered endemic of the SFRE or MNCE ecoregions, some of them occurring in a single basin of the latter, 23 in SFRE (Apareiodon hasemani, Prochilodus argenteus, P. costatus, Megaleporinus reinhardti, Schizodon knerii, Hemigrammus brevis, Orthospinus franciscensis, Phenacogaster julliae, Roeboides xenodon, Triportheus guentheri, Tatia bockmanni, Lophiosilurus alexandri, Pimelodella laurenti, Corydoras garbei, Crenicichla sp., Salminus franciscanus, Duopalatinus emarginatus, Pimelodus pohli, Pimelodella robinsoni, Pterygoplichthys etentaculatus, Conorhynchos conirostris, Cynolebias porosus, Hypsolebias flavicaudatus), and 13 in MNCE (Apareiodon davisi, Psectrogaster saguiru, Cheirodon jaguaribensis, Corydoras sp., Hypostomus sertanejo, Pimelodella dorseyi, P. enochi, Cynolebias microphthalmus, Hypsolebias antenori, some of them restricted to a single basin, such as Aspidoras menezesi, and Parotocinclus spilurus, in JAG, P. seridoensis in PIA, and P. spilosoma in PAR) (Fig. 3A, D-G). Some families must be highlighted: Loricariidae and Characidae with five endemic species each (38.46% and 19.23% of the species of this families, respectively), and Heptapteridae with four species (80.00%). Even though the total number of species is smaller, Rivulidae (4) and Parodontidae (2) included only endemic species (Tabs. 1, 3).
DISCUSSION
São Francisco IWT freshwater fish richness.Langeani et al. (2009)Langeani F, Buckup PA, Malabarba LR, Py-Daniel LHR, Lucena CAS, Rosa RS et al. Peixes de água doce. In: Rocha RM, Boeger WA, editors. Estado da arte e perspectivas para a Zoologia no Brasil. Curitiba: EDUFPR; 2009. p.211-30. mentioned the absence of comparative data between ichthyofauna of donor and receptor basins of the SFR-IWT. According to our updated database, 49 species (62.82%, considering only native and 56.98%, all species) of the SFRE do not occur in any MNCE receptor basin and might be potentially invasive species. It is important to note that, for species shared among donating and receiving basins, many are still in need of taxonomic revisions, which might change this number. Besides the taxonomic composition, the biological diversity expressed through populational variations are, certainly, present and could be addressed through phylogeographic studies (Craig et al., 2017Craig JM, Crampton WGR, Albert JS. Revision of the polytypic electric fish Gymnotus carapo (Gymnotiformes, Teleostei), with descriptions of seven subspecies. Zootaxa. 2017; 4318(3):401-38. https://doi.org/10.11646/zootaxa.4318.3.1
https://doi.org/10.11646/zootaxa.4318.3....
; Berbel-Filho et al., 2018Berbel-Filho WM, Ramos TPA, Jacobina UP, Maia DJG, Torres RA, Lima SMQ. Updated checklist and DNA barcode-based species delimitations reveal taxonomic uncertainties among freshwater fishes from the mid-north-eastern Caatinga ecoregion, north-eastern Brazil. J Fish Biol. 2018; 93(2):311-23. https://doi.org/10.1111/jfb.13758
https://doi.org/10.1111/jfb.13758...
). Moreover, in 2019 the team responsible for monitoring fish species during implementation of the SFR-IWT had already identified that three species from the SFRE must have arrived in PAR via the SFR-IWT canal. However, the scientific names of these species were not mentioned (Domingues, 2019Domingues F. Pelos canais do rio São Francisco ao menos três espécies de peixes chegam a lugares onde não existiam [Internet]. Globo; 2019. Available from: https://g1.globo.com/natureza/desafio-natureza/noticia/2019/12/22/pelos-canais-do-rio-sao-francisco-ao-menos-tres-especies-de-peixes-chegam-a-lugares-onde-nao-existiam.ghtml
https://g1.globo.com/natureza/desafio-na...
), corroborating the potential invasion risk of SFRE species in MNCE basins.
The SFR-IWT environmental impacts assessment (EIA) of the project pointed out possible environmental impacts, such as dissimilar fauna, supposedly, with only twenty-eight (23.93%, of 117 species listed in EIA) shared species between SFRE and MNCE, and possible quali-quantitative alterations in the ichthyofaunal composition of the receptor basin. However, the document minimizes the impacts that might be caused by the existence of eight non-native species (Astronotus ocellatus, Cichla monoculus, Lebistes reticulatus (= Poecilia reticulata), Leporinus “elongatus”, Oreochromis niloticus, Plagioscion squamosissimus, Poecilia latipinna, Poecilia vivipara (= considered as native in our study)) in the receptor basins and the low fish diversity (61 species, including non-native) considering its total drainage area (Brasil, 2004Brasil. Relatório de Impacto Ambiental-RIMA do Projeto de Integração do rio São Francisco com as Bacias Hidrográficas do Nordeste Setentrional. Brasília: Ministério da Integração Nacional/Ecology and Environment do Brasil; 2004.). Although the SFR-IWT EIA provides useful information about the fish fauna from the receptor basins, mainly considering the technical and systematics limitations of its time, it did not provide any species comparison among the drainage areas involved, and the taxonomy is outdated (Brasil, 2004Brasil. Relatório de Impacto Ambiental-RIMA do Projeto de Integração do rio São Francisco com as Bacias Hidrográficas do Nordeste Setentrional. Brasília: Ministério da Integração Nacional/Ecology and Environment do Brasil; 2004.).
Freshwater fish fauna basin surveys provide valuable information to detect changes in the species composition, mainly the input of species from the donor basin, such as those present in the lower-middle São Francisco basin in the SFR-IWT. The advances in the knowledge of SFR-IWT freshwater fish diversity were primarily due to the extensive surveys in the last decade, mainly conducted by regional research groups, whose fish collections are not yet online (Lima et al., 2017Lima SMQ, Ramos TPA, da Silva MJ, Rosa RS. Diversity, distribution, and conservation of the Caatinga fishes: advances and challenges. In: Silva, JMC, Leal IR, Tabarelli M, editors. Caatinga the largest tropical dry forest region in South America. Springer; 2017. p.97-131. https://doi.org/10.1007/978-3-319-68339-3_4
https://doi.org/10.1007/978-3-319-68339-...
). To make this database available, this systematic and geographic reference work on the freshwater fishes of the SFR-IWT, including sheets and shapefiles, will be available to fill this spatial gap in future regional or global analysis. This baseline might be useful to detect the establishment of non-native species, including those native to the SFRE via SFR-IWT canals.
Rosa (2004)Rosa RS. Diversidade e conservação dos peixes da caatinga. In: Silva JMC, Tabarelli M, Fonseca MT, Lins LV, editors. Biodiversidade da Caatinga: Áreas e Ações Prioritárias para a Conservação. Brasília: Ministério do Meio Ambiente; 2004. p. 149-61. conducted a comprehensive study of the Caatinga fishes based on literature data and showed species richness information for the main drainage areas of this biome, including three receptor ones from SFR-IWT: JAG (45 species), PIA (24), and PAR (23). These numbers included introduced species, but no species lists were provided for each basin. Another document that reports species richness in the SFR-IWT receptor basins is the SFR-IWT EIA (Brasil, 2004Brasil. Relatório de Impacto Ambiental-RIMA do Projeto de Integração do rio São Francisco com as Bacias Hidrográficas do Nordeste Setentrional. Brasília: Ministério da Integração Nacional/Ecology and Environment do Brasil; 2004.). The ichthyological list included primary data of 50 localities in the receptor basins surveyed between October and December 1998, secondary data (literature) from the donor basin, and interviews with riverine communities. It listed 86 species assigned to the lower-middle São Francisco basin, 52 to JAG, 21 to APO, 29 to PIA, and 34 to PAR, but considering only the native species (except for Poecilia vivipara, herein considered as native), they reduced it to 78 species in SFR, 47 in JAG, 17 in APO, 25 in PIA and 30 in PAR, numbers that are up to 46.88% lower than those currently listed (78 in SFR, 50 in JAG, 32 in APO, 39 in PIA and 36 in PAR) (Tab. 1). The updated values are also higher than those presented by a recent global database of freshwater fishes by basins (Tedesco et al., 2017Tedesco PA, Beauchard O, Bigorne R, Blanchet S, Buisson L, Conti L, et al. A global database on freshwater fish species occurrence in drainage basins. Scientific Data. 2017; 4:170141. https://doi.org/10.1038/sdata.2017.141
https://doi.org/10.1038/sdata.2017.141...
), which listed 31 species in JAG, 14 in APO, 17 in PIA and 18 in PAR. Recently, Ramos et al. (2018)Ramos TPA, Barros-Neto LF, Britski HA, Lima SMQ. Parotocinclus seridoensis, a new hypoptopomatine catfish (Siluriformes: Loricariidae) from the upper rio Piranhas-Açu basin, northeastern Brazil. Neotrop Ichthyol. 2013; 11(4):787-96. https://doi.org/10.1590/S1679-62252013000400006
https://doi.org/10.1590/S1679-6225201300...
and Rodrigues-Filho et al. (2019)Rodrigues-Filho CAS, Gurgel-Lourenço RC, Ramos EA, Novaes JLC, Garcez DS, Costa RS, Sánchez-Botero JI. Metacommunity organization in an intermittent river in Brazil: the importance of riverine networks for regional biodiversity. Aquat Ecol. 2019; 54:145-61. https://doi.org/10.1007/s10452-019-09732-1
https://doi.org/10.1007/s10452-019-09732...
provided inventories of continental fish species from PAR and JAG, respectively. The first, including vicarious division families, listed 47 species, but all freshwater species were also recorded in our study. The second, focused on the organization of fish metacommunities, lists 34 species, but contains misidentifications. Additionally, another 24 species were added in JAG. Considering the SFRE, in our study only the species from the lower-middle stretch were listed, resulting in 86 (78 native), which corresponds to 28.28% of the 304 species (241 native) of the whole ecoregion (Barbosa et al., 2017Barbosa JM, Soares EC, Cintra IHA, Hermann M, Araújo ARR. Perfil da ictiofauna da bacia do rio São Francisco. Acta Fish Aquat Res. 2017; 5(1):70-90. https://doi.org/10.2312/ActaFish.2017.5.1.70-90
https://doi.org/10.2312/ActaFish.2017.5....
). The upper and middle SFRE is highly different from the downstream stretches, with several highland taxa not present in the lowlands and semiarid.
Endemic and endangered species. Studies on the distribution of endemic species have important conservationist aims, mainly in the definition of protected areas, including their biotypes (Myers et al., 2000Myers N, Mittermeier RA, Mittermeier CG, Fonseca GAB da, Kent J. Biodiversity hotspots for conservation priorities. Nature. 2000; 403:853-8. https://doi.org/10.1038/35002501
https://doi.org/10.1038/35002501...
). The presence of 36 endemic species (32.43% of natives) in the studied basins corroborates the high endemism of Caatinga fishes (Lima et al., 2017Lima SMQ, Ramos TPA, da Silva MJ, Rosa RS. Diversity, distribution, and conservation of the Caatinga fishes: advances and challenges. In: Silva, JMC, Leal IR, Tabarelli M, editors. Caatinga the largest tropical dry forest region in South America. Springer; 2017. p.97-131. https://doi.org/10.1007/978-3-319-68339-3_4
https://doi.org/10.1007/978-3-319-68339-...
). The presence of endemic and endangered species in the receptor basins is worrying and deserves special attention, considering the ecological impacts caused by species introduction, which might range from behavioral shifts to complete alterations in the food web, extirpation of native species, and biotic homogenization (Kadye, Booth, 2012Kadye WT, Booth AJ. Detecting impacts of invasive non-native sharptooth catfish, Clarias gariepinus, within invaded and non-invaded rivers. Biodivers Conserv. 2012; 21:1997-2015. https://doi.org/10.1007/s10531-012-0291-5
https://doi.org/10.1007/s10531-012-0291-...
; Brito et al., 2020Brito MFG, Daga VS, Vitule JR. Fisheries and biotic homogenization of freshwater fish in the Brazilian semiarid region. Hydrobiologia. 2020; 847:3877-95. https://doi.org/10.1007/s10750-020-04236-8
https://doi.org/10.1007/s10750-020-04236...
). Habitat changes, added to invasion of predator or competitor non-native species, may reduce their abundance or distribution range (Shelton et al., 2017Shelton JM, Clark BM, Sephaka T, Turpie JK. Population crash in Lesotho’s endemic Maloti minnow Pseudobarbus quathlambae following invasion by translocated small mouth yellow fish Labeobarbus aeneus. Aquat Conserv. 2017; 27(1):65-77. https://doi.org/10.1002/aqc.2633
https://doi.org/10.1002/aqc.2633...
). Even endemic and threatened species from the São Francisco basin might suffer from lower hydrological levels, benefiting generalist species over specialists, and might also affect the seasonal killifish species inhabiting the floodplains of the basin (Costa, 2002Costa WJEM. Peixes Anuais Brasileiros: Diversidade e Conservação. Curitiba: Editora da UFPR; 2002.).
Leporinus taeniatus is referred in the literature as endemic to the SFRE (Reis et al., 2003Reis RE, Kullander SO, Ferraris-Jr CJ, editors. Check list of the freshwater fishes of South and Central America. Porto Alegre: EDIPUCRS; 2003.; Buckup et al., 2007Buckup PA, Menezes NA, Ghazzi MS. Catálogo das Espécies de Peixes de Água Doce do Brasil. Rio de Janeiro: Museu Nacional; 2007. ; Fricke et al., 2020Fricke R, Eschmeyer WN, Van-Der-Laan R, editors. Eschmeyer’s Catalog of Fishes: genera, species, references [Internet]. San Francisco: California Academy of Science; 2020. Available from: https://www.calacademy.org/scientists/projects/catalog-of-fishes
https://www.calacademy.org/scientists/pr...
). However, this species was collected in two out of the four MNCE receptor basins (JAG and APO). Even though there are no records of introduction of this species in the consulted source’s reports (Mattheus, 2005Mattheus S. América do Sul invadida: a crescente ameaça das espécies exóticas invasoras. Programa Global de Espécies Invasoras; 2005.; Leão et al., 2011Leão TCC, Almeida WR, Dechoum M, Ziller SR. Espécies exóticas invasoras no nordeste do Brasil: contextualização, manejo e políticas públicas. Recife: CEPAN; 2011.; Paiva, Mesquita, 2013Paiva MP, Mesquita PEC. Uma semente fecunda: commissão téchnica de piscicultura do Nordeste (1932-1945). Revista do Instituto do Ceará. 2013; 127:9-40.), the hypothesis of introduction cannot be disregarded, given that it is of commercial interest in SFRE (Bazzoli, 2003Bazzoli N. Parâmetros reprodutivos de peixes de interesse comercial do rio São Francisco, região de Pirapora, MG. In: Godinho HP, Godinho AL, editors. Águas, peixes e pescadores do São Francisco das Minas Gerais. Belo Horizonte: PUC Minas; 2003. p.286-300.), and that Megaleporinus obtusidens had already been introduced to the MNCE (Gurgel, Oliveira, 1987Gurgel JJS, Oliveira AG. Efeitos da introdução de peixes e crustáceos no Semi-árido do Nordeste brasileiro. Coleção Mossoroense. 1987; 453:1-32.).
Three Parotocinclus species are endemic to SFR-IWT receptor basins: P. spilurus in JAG, P. seridoensis in PIA, and P. spilosoma in PAR, and this number could be higher considering putative undescribed species (Lima et al., 2017Lima SMQ, Ramos TPA, da Silva MJ, Rosa RS. Diversity, distribution, and conservation of the Caatinga fishes: advances and challenges. In: Silva, JMC, Leal IR, Tabarelli M, editors. Caatinga the largest tropical dry forest region in South America. Springer; 2017. p.97-131. https://doi.org/10.1007/978-3-319-68339-3_4
https://doi.org/10.1007/978-3-319-68339-...
) (Fig. 3F). This might also be the case of some species not identified at species level (Loricariichthys sp. in JAG, Corydoras sp. in PIA, and Aspidoras sp. in SFRE) which can be putative undescribed species, confirming that ichthyofaunal taxonomic knowledge from the region is still incomplete, and strengthening the necessity for systematic research (Lima et al., 2017Lima SMQ, Ramos TPA, da Silva MJ, Rosa RS. Diversity, distribution, and conservation of the Caatinga fishes: advances and challenges. In: Silva, JMC, Leal IR, Tabarelli M, editors. Caatinga the largest tropical dry forest region in South America. Springer; 2017. p.97-131. https://doi.org/10.1007/978-3-319-68339-3_4
https://doi.org/10.1007/978-3-319-68339-...
). The Parotocinclus and Aspidoras catfishes are usually restricted to intermittent headwaters and could also be impacted by accumulation of fine sedimentary particles carried through the canals.
Final considerations and prospects. IWT projects are considered as one of the main threats to freshwater biodiversity, mainly to fish (Dudgeon, 2019Dudgeon D. Multiple threats imperil freshwater biodiversity in the Anthropocene. Curr Biol. 2019; 29(19):960-7. https://doi.org/10.1016/j.cub.2019.08.002
https://doi.org/10.1016/j.cub.2019.08.00...
). Furthermore, the modifications of the hydrological regime of the SFR-IWT basins can influence the seasonality, flow, availability and quality of the Caatinga rivers (Jardine et al., 2015Jardine TD, Woods R, Marshall J, Fawcett J, Lobegeiger J, Valdez D, Kainz MJ. Reconciling the role of organic matter pathways in aquatic food webs by measuring multiple tracers in individuals. Ecology. 2015; 96(12):3257-69. https://doi.org/10.1890/14-2153.1
https://doi.org/10.1890/14-2153.1...
). Consequently, these changes can lead to a reduction in fish diversity and inland fishery production (Sabo et al., 2017Sabo JL, Ruhi A, Holtgrieve GW, Elliott V, Arias ME, Ngor PB, et al. Designing river flows to improve food security futures in the Lower Mekong Basin. Science. 2017; 358(6368): eaao1053. https://doi.org/10.1126/science.aao1053
https://doi.org/10.1126/science.aao1053...
).
Deleterious anthropogenic alterations to the aquatic environment caused by large engineering projects are indisputable worldwide; however, obtaining reliable species richness lists before artificial IWT canals are implemented is difficult, albeit essential to detect species invasions (Johnson et al., 2008Johnson PTJ, Olden JD, Vander Zanden MJ. Dam invaders: impoundments facilitate biological invasions into freshwaters. Front Ecol Environ. 2008; 6(7):357-63. https://doi.org/10.1890/070156
https://doi.org/10.1890/070156...
). Deficiency in the pre-construction fish species baseline of the Lesotho Highland Water Project in Africa must have contributed to the disappearance of the endangered minnow Pseudobarbus quathlambae (Barnard 1938) in some areas, which was the only endemic species of the Lesotho highlands (Shelton et al., 2017Shelton JM, Clark BM, Sephaka T, Turpie JK. Population crash in Lesotho’s endemic Maloti minnow Pseudobarbus quathlambae following invasion by translocated small mouth yellow fish Labeobarbus aeneus. Aquat Conserv. 2017; 27(1):65-77. https://doi.org/10.1002/aqc.2633
https://doi.org/10.1002/aqc.2633...
). In the SFR-IWT basins, it was aggravated by the lack of an updated fish list by basins, which might hamper detection of the changes in richness due to the arrival of invasive species by the artificial canals. This study provides original lists of the freshwater fish species in each SFR-IWT basin, which should not considered as a final list, but as a baseline to detect changes after its operation.
This baseline of species richness and endemism (including endangered species) presents values higher than those previously stated (Rosa, 2004Rosa RS. Diversidade e conservação dos peixes da caatinga. In: Silva JMC, Tabarelli M, Fonseca MT, Lins LV, editors. Biodiversidade da Caatinga: Áreas e Ações Prioritárias para a Conservação. Brasília: Ministério do Meio Ambiente; 2004. p. 149-61.; Brasil, 2004Brasil. Relatório de Impacto Ambiental-RIMA do Projeto de Integração do rio São Francisco com as Bacias Hidrográficas do Nordeste Setentrional. Brasília: Ministério da Integração Nacional/Ecology and Environment do Brasil; 2004.), as well as making the first comparisons of shared species among donor and receptor SFR-IWT basins. These results cause concern, due to the potential risk of non-native species invading the receptor basins. To the extent that invasive species settle down, the faunal homogenization process will begin (Brito et al., 2020Brito MFG, Daga VS, Vitule JR. Fisheries and biotic homogenization of freshwater fish in the Brazilian semiarid region. Hydrobiologia. 2020; 847:3877-95. https://doi.org/10.1007/s10750-020-04236-8
https://doi.org/10.1007/s10750-020-04236...
); and possible local and even total extinctions of endemic species might occur in this ichthyofauna adapted to the temporary hydrological regimen (Silva, 2017Silva MJ. Diversidade e conservação da ictiofauna das bacias envolvidas no Projeto de Transposição do rio São Francisco. [PhD Thesis]. Natal: Universidade Federal do Rio Grande do Norte; 2017. Available from: https://repositorio.ufrn.br/jspui/handle/123456789/23669
https://repositorio.ufrn.br/jspui/handle...
).
The SFR-IWT canals are already functioning, and special attention should be given to implementing ichthyofaunal protection measures, such as aquatic biota block barriers, mainly to avoid the faunal exchange between basins. The priority actions for the SFR-IWT basins should include the (1) sustainable limits of freshwater withdrawal, and (2) halting expansion of IWT projects, and (3) constant monitoring, especially of threatened and endemic freshwater fish species of the semiarid drainage areas.
To avoid invasive species, it is necessary to build efficient barriers to restrain faunal exchange among the SFR-IWT basins (Brasil, 2020Brasil. Licença de Instalação [Internet]. Brasília: Ministério do Desenvolvimento Regional; 2020. Available from: https://www.mdr.gov.br/seguranca-hidrica/projeto-rio-sao-francisco/o-que-e-o-projeto/documentos-tecnicos
https://www.mdr.gov.br/seguranca-hidrica...
). Behavioral and physical barriers should be adequate to block migrant and carnivore fish that occur in the catchment area (e.g. Leporinus, Megaleporinus, Pygocentrus, and Plagioscion species), which may see their chance of establishment in modified environments rise up to eight times in relation to natural conditions (Johnson et al., 2008Johnson PTJ, Olden JD, Vander Zanden MJ. Dam invaders: impoundments facilitate biological invasions into freshwaters. Front Ecol Environ. 2008; 6(7):357-63. https://doi.org/10.1890/070156
https://doi.org/10.1890/070156...
). Of the 11 non-native species listed herein, Xiphophorus helleri and Parachromis managuensis are considered as recently introduced (Rodrigues-Filho et al., 2016Rodrigues-Filho CAS, Gurgel-Lourenço RC, Bezerra LAV, Sousa WA, Garcez DS, Lima SMQ et al. Ichthyofauna of the humid forest enclaves in the tablelands of Ibiapaba and Araripe, Northeastern Brazil. Biota Neotrop. 2016; 16(4):e20160273. https://doi.org/10.1590/1676-0611-BN-2016-0273
https://doi.org/10.1590/1676-0611-BN-201...
; França et al., 2017França EJ, Almeida CAC, Almeida-Neto MS, Santos RE, Magalhães ALB, El-Deir ACA, Severi W. Novelty on the market, novelty in the environment: The invasion of non-native fish jaguar guapote (Perciformes) in northeastern Brazil. Neotropical Biol Conserv. 2017; 12(1):12-8. https://doi.org/10.4013/nbc.2017.121.02
https://doi.org/10.4013/nbc.2017.121.02...
). However, the second, an exotic predator cichlid, was recorded in the SFRE and in PAR among the receptor basins, and could invade the other basins throughout the canals (França et al., 2017França EJ, Almeida CAC, Almeida-Neto MS, Santos RE, Magalhães ALB, El-Deir ACA, Severi W. Novelty on the market, novelty in the environment: The invasion of non-native fish jaguar guapote (Perciformes) in northeastern Brazil. Neotropical Biol Conserv. 2017; 12(1):12-8. https://doi.org/10.4013/nbc.2017.121.02
https://doi.org/10.4013/nbc.2017.121.02...
; Ramos et al., 2018Ramos TPA, Lima JAS, Costa SYL, Silva MJ, Avellar RC, Oliveira-Silva L. Continental ichthyofauna from the Paraíba do Norte River basin pre-transposition of the São Francisco River, Northeastern Brazil. Biota Neotrop. 2018; 18(4):e20170471. https://doi.org/10.1590/1676-0611-bn-2017-0471
https://doi.org/10.1590/1676-0611-bn-201...
) (Fig. 3K-L).
Environmental changes can force the substitution of native species populations by those from the donor basin, which might be more adapted to the perennial regime resulting from water transfer (Shelton et al., 2017Shelton JM, Clark BM, Sephaka T, Turpie JK. Population crash in Lesotho’s endemic Maloti minnow Pseudobarbus quathlambae following invasion by translocated small mouth yellow fish Labeobarbus aeneus. Aquat Conserv. 2017; 27(1):65-77. https://doi.org/10.1002/aqc.2633
https://doi.org/10.1002/aqc.2633...
). Nevertheless, Lynch et al. (2011)Lynch HJ, Grant EHC, Muneepeerakul R, Arunachalam M, Rodriguez-Iturbe I, Fagan WF. How restructuring river connectivity changes freshwater fish biodiversity and biogeography. Water Resour Res. 2011; 47(5):1-10. https://doi.org/10.1029/2010WR010330
https://doi.org/10.1029/2010WR010330...
recognized the higher possibility of faunal composition changes among distinct hydrographic basins in the Par Tapi Narmada Link Project canals in India, and that in long canals (over 550 km) there is a reduction in the probability of species invasion, as only some can run those long distances. In the case of SFR-IWT, it is worrying taking into account the canals’ intermediate size (357 km in North Axis and 220 in East Axis), in which it is possible that migratory species such as Prochilodus, Pimelodus, Megaleporinus and other medium and large-sized-species from the SFRE may reach the receptor basins. Other studies verified the establishment of invasive species in rivers with altered abiotic conditions and river flow, including IWT canals (Kadye, Booth, 2012Kadye WT, Booth AJ. Detecting impacts of invasive non-native sharptooth catfish, Clarias gariepinus, within invaded and non-invaded rivers. Biodivers Conserv. 2012; 21:1997-2015. https://doi.org/10.1007/s10531-012-0291-5
https://doi.org/10.1007/s10531-012-0291-...
). We hope, however, that this baseline is a starting point for predictive assessments of non-native species (e.g. ecological niche modeling) in the SFR-IWT receptor basins.
ACKNOWLEDGEMENTS
This study was funded by CNPq/ICMBio (Conselho Nacional de Desenvolvimento Científico e Tecnológico/Instituto Chico Mendes de Conservação da Biodiversidade; process number: 552086/2011-8). Márcio Silva was supported by a doctoral scholarship from CAPES (Coordenação de Aperfeiçoamento de Pessoal de Nível Superior). Telton Ramos received a DCR (Desenvolvimento Científico Regional) post-doctoral fellowship from CNPq/FAPERN (Fundação de Apoio à Pesquisa do Estado do Rio Grande do Norte, Proc. 350674/2012-4). Sergio Lima and Marcelo Brito received CNPq’s productivity research grants (process numbers: 313644/2018-7; 306851/2018-0, respectively). All authors are grateful to the students of Laboratório de Ictiologia Sistemática e Evolutiva and Grupo de Estudos de Ecologia e Fisiologia de Animais Aquáticos from the Universidade Federal do Rio Grande do Norte for the fieldwork assistance. Additionally, the authors thank MMA/ICMBio/SISBIO (Ministério Brasileiro do Meio Ambiente/Instituto Chico Mendes de Conservação da Biodiversidade/Sistema de Autorização e Informação em Biodiversidade) for collection permits. Finally, the authors are grateful to the anonymous reviewers who kindly made valuable suggestions throughout the peer review process of this paper.
REFERENCES
- Abell R, Thieme ML, Revenga C, Bryer M, Kottelat M, Bogutskaya N et al. Freshwater ecoregions of the world: a new map of biogeographic units for freshwater biodiversity conservation. Bioscience. 2008; 58(5):403-14. https://doi.org/10.1641/B580507
» https://doi.org/10.1641/B580507 - Albert JS, Carvalho TP. Neogene assembly of modern faunas. In: Albert JS, Reis RE, editors. Historical biogeography of Neotropical freshwater fishes. Berkeley and Los Angeles, California: University of California Press; 2011. p.119-36.
- Albert JS, Destouni G, Duke-Sylvester SM, Magurran AE, Oberdorff T, Reis RE et al. Scientist's warning to humanity on the freshwater biodiversity crisis. Ambio. 2020. https://doi.org/10.1007/s13280-020-01318-8
» https://doi.org/10.1007/s13280-020-01318-8 - Albert JS, Petry P, Reis RE. Major biogeographic and phylogenetic patterns. In: Albert JS, Reis RE, editors. Historical biogeography of Neotropical freshwater fishes. Berkeley and Los Angeles, California: University of California Press; 2011. p.21-57.
- Albuquerque UP, Araújo EL, El-Deir ACA, Lima ALA, Souto A, Bezerra BM et al. Caatinga revisited: ecology and conservation of an important seasonal dry forest. Sci World J. 2012; 205182:1-18. https://doi.org/10.1100/2012/205182
» https://doi.org/10.1100/2012/205182 - Alves CBM, Vieira F, Pompeu PS. Ictiofauna da bacia hidrográfica do rio São Francisco. Diagnóstico do macrozoneamento ecológico-econômico da Bacia Hidrográfica do Rio São Francisco. Brasília: Ministério do Meio Ambiente; 2011. p.226-41.
- Andrade JGP, Barbosa PSF, Souza LCA, Makino DL. Interbasin water transfers: The Brazilian experience and international case comparisons. Water Resour Manag. 2011; 25(8):1915-34. https://doi.org/10.1007/s11269-011-9781-6
» https://doi.org/10.1007/s11269-011-9781-6 - Arrhenius O. Species and area. J Ecol. 1921; 9:95-99. https://doi.org/10.2307/2255763
» https://doi.org/10.2307/2255763 - Barbosa JM, Soares EC, Cintra IHA, Hermann M, Araújo ARR. Perfil da ictiofauna da bacia do rio São Francisco. Acta Fish Aquat Res. 2017; 5(1):70-90. https://doi.org/10.2312/ActaFish.2017.5.1.70-90
» https://doi.org/10.2312/ActaFish.2017.5.1.70-90 - Barletta M, Jaureguizar AJ, Baigun C, Fontoura NF, Agostinho AA, Almeida-Val VMF et al. Fish and aquatic habitat conservation in South America: a continental overview with emphasis on Neotropical systems. J Fish Biol. 2010; 76(9):2118-76. https://doi.org/10.1111/j.1095-8649.2010.02684.x
» https://doi.org/10.1111/j.1095-8649.2010.02684.x - Bazzoli N. Parâmetros reprodutivos de peixes de interesse comercial do rio São Francisco, região de Pirapora, MG. In: Godinho HP, Godinho AL, editors. Águas, peixes e pescadores do São Francisco das Minas Gerais. Belo Horizonte: PUC Minas; 2003. p.286-300.
- Berbel-Filho WM, Ramos TPA, Jacobina UP, Maia DJG, Torres RA, Lima SMQ. Updated checklist and DNA barcode-based species delimitations reveal taxonomic uncertainties among freshwater fishes from the mid-north-eastern Caatinga ecoregion, north-eastern Brazil. J Fish Biol. 2018; 93(2):311-23. https://doi.org/10.1111/jfb.13758
» https://doi.org/10.1111/jfb.13758 - Brasil. Relatório de Impacto Ambiental-RIMA do Projeto de Integração do rio São Francisco com as Bacias Hidrográficas do Nordeste Setentrional. Brasília: Ministério da Integração Nacional/Ecology and Environment do Brasil; 2004.
- Brasil. Metadados [Internet]. Agência Nacional de Águas; 2014a. Available from: http://metadados.ana.gov.br/geonetwork/srv/pt/main.home
» http://metadados.ana.gov.br/geonetwork/srv/pt/main.home - Brasil. Dados georreferenciados [Internet]. Ministério do Meio Ambiente; 2014b. Available from: http://www.mma.gov.br/areas-protegidas/cadastro-nacional-de-ucs/dados-georreferenciados
» http://www.mma.gov.br/areas-protegidas/cadastro-nacional-de-ucs/dados-georreferenciados - Brasil. Portaria Ministério do Meio Ambiente n° 445, de 17 de Dezembro de 2014. Lista Nacional Oficial de Espécies da Fauna Ameaçada de Extinção: Peixes e Invertebrados Aquáticos. Brasília: Diário Oficial da União; 2014c. p.126-30
- Brasil. Projeto de Integração do Rio São Francisco: o andamento das obras [Internet]. Brasília: Ministério da Integração Nacional; 2019. Available from: http://www.mi.gov.br/web/projeto-sao-francisco/o-andamento-das-obras
» http://www.mi.gov.br/web/projeto-sao-francisco/o-andamento-das-obras - Brasil. Licença de Instalação [Internet]. Brasília: Ministério do Desenvolvimento Regional; 2020. Available from: https://www.mdr.gov.br/seguranca-hidrica/projeto-rio-sao-francisco/o-que-e-o-projeto/documentos-tecnicos
» https://www.mdr.gov.br/seguranca-hidrica/projeto-rio-sao-francisco/o-que-e-o-projeto/documentos-tecnicos - Brito MFG, Daga VS, Vitule JR. Fisheries and biotic homogenization of freshwater fish in the Brazilian semiarid region. Hydrobiologia. 2020; 847:3877-95. https://doi.org/10.1007/s10750-020-04236-8
» https://doi.org/10.1007/s10750-020-04236-8 - Buckup PA, Menezes NA, Ghazzi MS. Catálogo das Espécies de Peixes de Água Doce do Brasil. Rio de Janeiro: Museu Nacional; 2007.
- Castro RMC, Polaz CNM. Small-sized fish: the largest and most threatened portion of the megadiverse neotropical freshwater fish fauna. Biota Neotrop. 2020; 20(1):e20180683. https://doi.org/10.1590/1676-0611-bn-2018-0683
» https://doi.org/10.1590/1676-0611-bn-2018-0683 - CBHSF (Comitê da Bacia Hidrográfica do Rio São Francisco). Shapefiles [Internet]. CBHSF; 2014. Available from: http://www.saofrancisco.cbh.gov.br/Shapes/SHAPE.sigeo.Transposicao.SF.zip
» http://www.saofrancisco.cbh.gov.br/Shapes/SHAPE.sigeo.Transposicao.SF.zip - Connor EF, McCoy ED. The statistics and biology of the species area relationship. Am Nat. 1979; 113:791-833. Available from: http://www.jstor.org/stable/2460305
» http://www.jstor.org/stable/2460305 - Costa WJEM. Peixes Anuais Brasileiros: Diversidade e Conservação. Curitiba: Editora da UFPR; 2002.
- Craig JM, Crampton WGR, Albert JS. Revision of the polytypic electric fish Gymnotus carapo (Gymnotiformes, Teleostei), with descriptions of seven subspecies. Zootaxa. 2017; 4318(3):401-38. https://doi.org/10.11646/zootaxa.4318.3.1
» https://doi.org/10.11646/zootaxa.4318.3.1 - Dengler J. Which function describes the species-area relationship best? A review and empirical evaluation. J Biogeogr. 2009; 36(4):728-44. https://doi.org/10.1111/j.1365-2699.2008.02038.x
» https://doi.org/10.1111/j.1365-2699.2008.02038.x - Dias MS, Zuanon J, Couto TBA, Carvalho M, Carvalho LN, Espirito-Santo HMV et al. Trends in studies of Brazilian stream fish assemblages. Nat Conserv. 2016; 14(2):106-11. https://doi.org/10.1016/j.ncon.2016.06.003
» https://doi.org/10.1016/j.ncon.2016.06.003 - DNOCS (Departamento Nacional de Obras Contra a Seca). Relatório das atividades desenvolvidas pela coordenação de pesca e aquicultura, durante o ano de 2002. Coordenação de Pesca e Aquicultura; 2002.
- Domingues F. Pelos canais do rio São Francisco ao menos três espécies de peixes chegam a lugares onde não existiam [Internet]. Globo; 2019. Available from: https://g1.globo.com/natureza/desafio-natureza/noticia/2019/12/22/pelos-canais-do-rio-sao-francisco-ao-menos-tres-especies-de-peixes-chegam-a-lugares-onde-nao-existiam.ghtml
» https://g1.globo.com/natureza/desafio-natureza/noticia/2019/12/22/pelos-canais-do-rio-sao-francisco-ao-menos-tres-especies-de-peixes-chegam-a-lugares-onde-nao-existiam.ghtml - Dudgeon D. Multiple threats imperil freshwater biodiversity in the Anthropocene. Curr Biol. 2019; 29(19):960-7. https://doi.org/10.1016/j.cub.2019.08.002
» https://doi.org/10.1016/j.cub.2019.08.002 - Dyrnes GV, Vatn A. Who owns the water? A study of a water conflict in the Valley of Ixtlahuaca, México. Water Policy. 2005; 7(3):295-312. https://doi.org/10.2166/wp.2005.0019
» https://doi.org/10.2166/wp.2005.0019 - França EJ, Almeida CAC, Almeida-Neto MS, Santos RE, Magalhães ALB, El-Deir ACA, Severi W. Novelty on the market, novelty in the environment: The invasion of non-native fish jaguar guapote (Perciformes) in northeastern Brazil. Neotropical Biol Conserv. 2017; 12(1):12-8. https://doi.org/10.4013/nbc.2017.121.02
» https://doi.org/10.4013/nbc.2017.121.02 - Fricke R, Eschmeyer WN, Van-Der-Laan R, editors. Eschmeyer’s Catalog of Fishes: genera, species, references [Internet]. San Francisco: California Academy of Science; 2020. Available from: https://www.calacademy.org/scientists/projects/catalog-of-fishes
» https://www.calacademy.org/scientists/projects/catalog-of-fishes - Gupta J, Van der Zaag P. Interbasin water transfers and integrated water resources management: Where engineering, science and politics interlock. Phys Chem Earth. 2008; 33(1-2):28-40. https://doi.org/10.1016/j.pce.2007.04.003
» https://doi.org/10.1016/j.pce.2007.04.003 - Gurgel JJS, Fernando CH. Fisheries in Semi-Arid Northeast Brazil with special reference to the role of Tilapias. Int Rev Hydrobiol. 1994; 79(1):77-94. https://doi.org/10.1002/iroh.19940790109
» https://doi.org/10.1002/iroh.19940790109 - Gurgel JJS, Oliveira AG. Efeitos da introdução de peixes e crustáceos no Semi-árido do Nordeste brasileiro. Coleção Mossoroense. 1987; 453:1-32.
- India (National Water Development Agency). Par Tapi Narmada Link Project. India: Ministry of Jal Shakti; 2017. p.844–926. Available from: http:// nwda.gov.in/content/innerpage/par-tapinarmada-link-project.php
» http:// nwda.gov.in/content/innerpage/par-tapinarmada-link-project.php - Jardine TD, Woods R, Marshall J, Fawcett J, Lobegeiger J, Valdez D, Kainz MJ. Reconciling the role of organic matter pathways in aquatic food webs by measuring multiple tracers in individuals. Ecology. 2015; 96(12):3257-69. https://doi.org/10.1890/14-2153.1
» https://doi.org/10.1890/14-2153.1 - Johnson PTJ, Olden JD, Vander Zanden MJ. Dam invaders: impoundments facilitate biological invasions into freshwaters. Front Ecol Environ. 2008; 6(7):357-63. https://doi.org/10.1890/070156
» https://doi.org/10.1890/070156 - Kadye WT, Booth AJ. Detecting impacts of invasive non-native sharptooth catfish, Clarias gariepinus, within invaded and non-invaded rivers. Biodivers Conserv. 2012; 21:1997-2015. https://doi.org/10.1007/s10531-012-0291-5
» https://doi.org/10.1007/s10531-012-0291-5 - Langeani F, Buckup PA, Malabarba LR, Py-Daniel LHR, Lucena CAS, Rosa RS et al. Peixes de água doce. In: Rocha RM, Boeger WA, editors. Estado da arte e perspectivas para a Zoologia no Brasil. Curitiba: EDUFPR; 2009. p.211-30.
- Leão TCC, Almeida WR, Dechoum M, Ziller SR. Espécies exóticas invasoras no nordeste do Brasil: contextualização, manejo e políticas públicas. Recife: CEPAN; 2011.
- Lesotho (Lesotho Highlands Development Authority). Project description [Internet]. Lesotho; 2020. Available from: http://www.lhda.org.ls/lhdaweb
» http://www.lhda.org.ls/lhdaweb - Lévêque C, Balian EV, Martens K. An assessment of animal species diversity in continental waters. Hydrobiologia. 2005; 542:39-67. https://doi.org/10.1007/s10750-004-5522-7
» https://doi.org/10.1007/s10750-004-5522-7 - Lévêque C, Oberdorff T, Paugy D, Stiassny MLJ, Tedesco PA. Global diversity of fish (Pisces) in freshwater. Hydrobiologia. 2008; 595:545-67. https://doi.org/10.1007/978-1-4020-8259-7_53
» https://doi.org/10.1007/978-1-4020-8259-7_53 - Li J, Dong S, Peng M, Yang Z, Liu S, Li X et al. Effects of damming on the biological integrity of fish assemblages in the middle Lancang-Mekong River basin. Ecol Indic. 2013; 34:94-102. https://doi.org/10.1016/j.ecolind.2013.04.016
» https://doi.org/10.1016/j.ecolind.2013.04.016 - Lima LC. Além das águas, a discussão no nordeste do rio São Francisco. Revista do Departamento de Geografia. 2005; 17:94-100. https://doi.org/10.7154/RDG.2005.0017.0007
» https://doi.org/10.7154/RDG.2005.0017.0007 - Lima FCT, Britski HA. Salminus franciscanus, a new species from the Rio São Francisco basin, Brazil (Ostariophysi: Characiformes: Characidae). Neotrop Ichthyol. 2007; 5(3):237-44. https://doi.org/10.1590/S1679-62252007000300001
» https://doi.org/10.1590/S1679-62252007000300001 - Lima SMQ, Ramos TPA, da Silva MJ, Rosa RS. Diversity, distribution, and conservation of the Caatinga fishes: advances and challenges. In: Silva, JMC, Leal IR, Tabarelli M, editors. Caatinga the largest tropical dry forest region in South America. Springer; 2017. p.97-131. https://doi.org/10.1007/978-3-319-68339-3_4
» https://doi.org/10.1007/978-3-319-68339-3_4 - Long D, Yang W, Scanlon BR, Zhao J, Liu D, Burek P et al. South-to-North Water Diversion stabilizing Beijing’s groundwater levels. Nat Commun. 2020; 11(3665). https://doi.org/10.1038/s41467-020-17428-6
» https://doi.org/10.1038/s41467-020-17428-6 - Lucena CAS. Revisão taxonômica das espécies do gênero Roeboides grupo-affinis (Ostariophysi, Characiformes, Characidae). Iheringia Ser Zool. 2007; 97(2):117-36. https://doi.org/10.1590/S0073-47212007000200001
» https://doi.org/10.1590/S0073-47212007000200001 - Luz SCS, Lima HC, Severi W. Composição da ictiofauna em ambientes marginais e tributários do médio-submédio rio São Francisco. Braz J Agricult Sci. 2012; 7(2):358-66. https://doi.org/10.5039/agraria.v7i2a1436
» https://doi.org/10.5039/agraria.v7i2a1436 - L’vovich MI, White GF. Use and transformation of terrestrial water systems. In: Turner-II BL, Clark WC, Kates RW, Richards JF, Mathews JT, Meyer WB, editors. The Earth as transformed by human action. University Cambridge Press; 1990. p.235-52.
- Lynch HJ, Grant EHC, Muneepeerakul R, Arunachalam M, Rodriguez-Iturbe I, Fagan WF. How restructuring river connectivity changes freshwater fish biodiversity and biogeography. Water Resour Res. 2011; 47(5):1-10. https://doi.org/10.1029/2010WR010330
» https://doi.org/10.1029/2010WR010330 - Mattheus S. América do Sul invadida: a crescente ameaça das espécies exóticas invasoras. Programa Global de Espécies Invasoras; 2005.
- Meador MR. Inter-basin water transfer: Ecological concerns. Fisheries. 1992; 17(2):17-22. https://doi.org/10.1577/1548-8446(1992)017<0017:IWTEC>2.0.CO;2
» https://doi.org/10.1577/1548-8446(1992)017<0017:IWTEC>2.0.CO;2 - Meybeck M. Global analysis of river systems: from Earth system controls to Anthropocene syndromes. Philos Trans R Soc Lond B Biol Sci. 2003; 358(1440):1935-55. https://doi.org/10.1098/rstb.2003.1379
» https://doi.org/10.1098/rstb.2003.1379 - Moreira-Filho O, Buckup PA. A poorly known case of watershed transposition between the São Francisco and upper Paraná river basins. Neotrop Ichthyol. 2005; 3(3):449-52. https://doi.org/10.1590/S1679-62252005000300015
» https://doi.org/10.1590/S1679-62252005000300015 - Muller M. Interbasin water sharing: A South African perspective. In: Proceedings of the International Workshop. Paris: UNESCO, Technical Documents in Hydrology 28, International Hydrological Program - V; 1999. p.61-70.
- Myers GS. Fresh-water fishes and west Indian zoogeography. Annu Rep Board Regents Smithson Inst. 1937; 3(465):339-64.
- Myers N, Mittermeier RA, Mittermeier CG, Fonseca GAB da, Kent J. Biodiversity hotspots for conservation priorities. Nature. 2000; 403:853-8. https://doi.org/10.1038/35002501
» https://doi.org/10.1038/35002501 - Nascimento WS, Barros NHC, Araújo AS, Gurgel LL, Canan B, Molina WF, et al. Composição da ictiofauna das bacias hidrográficas do Rio Grande do Norte, Brasil. Biota Amazônia. 2014; 4(1):126-31. https://doi.org/10.18561/2179-5746/biotaamazonia.v4n1p126-131
» https://doi.org/10.18561/2179-5746/biotaamazonia.v4n1p126-131 - Paiva MP, Mesquita PEC. Uma semente fecunda: commissão téchnica de piscicultura do Nordeste (1932-1945). Revista do Instituto do Ceará. 2013; 127:9-40.
- Pittock J, Meng JH, Geiger M, Chapagain AK. Interbasin water transfers and water scarcity in a changing world - a solution or a pipedream?. Germany: WWF; 2009.
- Ramos TPA, Barros-Neto LF, Britski HA, Lima SMQ. Parotocinclus seridoensis, a new hypoptopomatine catfish (Siluriformes: Loricariidae) from the upper rio Piranhas-Açu basin, northeastern Brazil. Neotrop Ichthyol. 2013; 11(4):787-96. https://doi.org/10.1590/S1679-62252013000400006
» https://doi.org/10.1590/S1679-62252013000400006 - Ramos TPA, Lima JAS, Costa SYL, Silva MJ, Avellar RC, Oliveira-Silva L. Continental ichthyofauna from the Paraíba do Norte River basin pre-transposition of the São Francisco River, Northeastern Brazil. Biota Neotrop. 2018; 18(4):e20170471. https://doi.org/10.1590/1676-0611-bn-2017-0471
» https://doi.org/10.1590/1676-0611-bn-2017-0471 - Ramos TPA, Ramos RTC, Ramos SAQA. Ichthyofauna of the Parnaíba river basin, Northeastern Brazil. Biota Neotrop. 2014; 14(1):1-8. https://doi.org/10.1590/S1676-06020140039
» https://doi.org/10.1590/S1676-06020140039 - Reis RE, Albert JS, Di Dario F, Mincarone MM, Petry P, Rocha LA. Fish biodiversity and conservation in South America. J Fish Biol. 2016; 89(1):12-47. https://doi.org/10.1111/jfb.13016
» https://doi.org/10.1111/jfb.13016 - Reis RE, Kullander SO, Ferraris-Jr CJ, editors. Check list of the freshwater fishes of South and Central America. Porto Alegre: EDIPUCRS; 2003.
- Revenga C, Campbell I, Abell R, Villiers P, Bryer M. Prospects for monitoring freshwater ecosystems towards the 2010 targets. Philos Trans R Soc Lond B Biol Sci. 2005; 360(1454):397-413. https://doi.org/10.1098/rstb.2004.1595
» https://doi.org/10.1098/rstb.2004.1595 - Ribeiro FRV, Lucena CAS. A new species of Pimelodus LaCépède, 1803 (Siluriformes: Pimelodidae) from the rio São Francisco drainage, Brazil. Neotrop Ichthyol. 2006; 4(4):411-8. https://doi.org/10.1590/S1679-62252006000400004
» https://doi.org/10.1590/S1679-62252006000400004 - Rodrigues-Filho CAS, Gurgel-Lourenço RC, Bezerra LAV, Sousa WA, Garcez DS, Lima SMQ et al. Ichthyofauna of the humid forest enclaves in the tablelands of Ibiapaba and Araripe, Northeastern Brazil. Biota Neotrop. 2016; 16(4):e20160273. https://doi.org/10.1590/1676-0611-BN-2016-0273
» https://doi.org/10.1590/1676-0611-BN-2016-0273 - Rodrigues-Filho CAS, Gurgel-Lourenço RC, Ramos EA, Novaes JLC, Garcez DS, Costa RS, Sánchez-Botero JI. Metacommunity organization in an intermittent river in Brazil: the importance of riverine networks for regional biodiversity. Aquat Ecol. 2019; 54:145-61. https://doi.org/10.1007/s10452-019-09732-1
» https://doi.org/10.1007/s10452-019-09732-1 - Rosa RS. Diversidade e conservação dos peixes da caatinga. In: Silva JMC, Tabarelli M, Fonseca MT, Lins LV, editors. Biodiversidade da Caatinga: Áreas e Ações Prioritárias para a Conservação. Brasília: Ministério do Meio Ambiente; 2004. p. 149-61.
- Rosa RS, Gomes-Filho G, Menezes NA, Shibatta OA, Costa WJEM. Biota aquática: áreas e ações prioritárias para conservação da caatinga. In: Silva JMC, Tabarelli M, Fonseca MT, Lins LV, editors. Biodiversidade da caatinga: áreas e ações prioritárias para a conservação. Brasília: Ministério do Meio Ambiente; 2004. p.163-71.
- Rosa RS, Menezes NA, Britski HA, Costa WJEM, Groth F. Diversidade, padrões de distribuição e conservação dos peixes da Caatinga. In: Leal IR, Silva JMC, Tabarelli M, editors. Ecologia e Conservação da Caatinga. Recife: EDUFPE; 2003. p.135-81.
- Sabo JL, Ruhi A, Holtgrieve GW, Elliott V, Arias ME, Ngor PB, et al. Designing river flows to improve food security futures in the Lower Mekong Basin. Science. 2017; 358(6368): eaao1053. https://doi.org/10.1126/science.aao1053
» https://doi.org/10.1126/science.aao1053 - Shelton JM, Clark BM, Sephaka T, Turpie JK. Population crash in Lesotho’s endemic Maloti minnow Pseudobarbus quathlambae following invasion by translocated small mouth yellow fish Labeobarbus aeneus Aquat Conserv. 2017; 27(1):65-77. https://doi.org/10.1002/aqc.2633
» https://doi.org/10.1002/aqc.2633 - Silva MJ. Diversidade e conservação da ictiofauna das bacias envolvidas no Projeto de Transposição do rio São Francisco. [PhD Thesis]. Natal: Universidade Federal do Rio Grande do Norte; 2017. Available from: https://repositorio.ufrn.br/jspui/handle/123456789/23669
» https://repositorio.ufrn.br/jspui/handle/123456789/23669 - Silva MJ, Costa BG, Ramos TPA, Auricchio P, Lima SMQ. Ichthyofauna of the Gurgueia river, Parnaíba river basin, Northeastern Brazil. Check List. 2015; 11(5):1-8. https://doi.org/10.15560/11.5.1765
» https://doi.org/10.15560/11.5.1765 - Silva JMC, Tabarelli M, Fonseca MT, Lins LV, editors. Biodiversidade da Caatinga: áreas e ações prioritárias para a conservação. Brasília: Ministério do Meio Ambiente; 2004.
- Tedesco PA, Beauchard O, Bigorne R, Blanchet S, Buisson L, Conti L, et al. A global database on freshwater fish species occurrence in drainage basins. Scientific Data. 2017; 4:170141. https://doi.org/10.1038/sdata.2017.141
» https://doi.org/10.1038/sdata.2017.141 - 22 Uieda VS, Castro RMC. Coleta e fixação de peixes de riachos. In: Caramaschi EP, Mazzoni R, Peres-Neto PR, editors. Ecologia de Peixes de Riachos. Rio de Janeiro: Oecologia Brasiliensis; 1999. p.22.
- Van der Zaag P. Asymmetry and equity in water resources management; critical governance issues for Southern Africa. Water Resour Manag. 2007; 21(12):1993-2004. https://doi.org/10.1007/s11269-006-9124-1
» https://doi.org/10.1007/s11269-006-9124-1 - Zawadzki CH, Ramos TPA, Sabaj M. Hypostomus sertanejo (Siluriformes: Loricariidae), new armoured catfish species from north-eastern Brazil. J Fish Biol. 2017; 91(1):317-30. https://doi.org/10.1111/jfb.13349
» https://doi.org/10.1111/jfb.13349
ADDITIONAL NOTES
-
HOW TO CITE THIS ARTICLE
Silva MJ, Ramos TPA, Carvalho FR, Brito MFG, Ramos RTC, Rosa RS, Sánchez-Botero JI, Novaes JLC, Costa RS, Lima SMQ. Freshwater fish richness baseline from the São Francisco Interbasin Water Transfer Project in the Brazilian Semiarid. Neotrop Ichthyol. 2020; 18(4):e200063. https://doi.org/10.1590/1982-0224-2020-0063
Edited by
Data availability
Data citations
Brasil. Metadados [Internet]. Agência Nacional de Águas; 2014a. Available from: http://metadados.ana.gov.br/geonetwork/srv/pt/main.home
Brasil. Dados georreferenciados [Internet]. Ministério do Meio Ambiente; 2014b. Available from: http://www.mma.gov.br/areas-protegidas/cadastro-nacional-de-ucs/dados-georreferenciados
CBHSF (Comitê da Bacia Hidrográfica do Rio São Francisco). Shapefiles [Internet]. CBHSF; 2014. Available from: http://www.saofrancisco.cbh.gov.br/Shapes/SHAPE.sigeo.Transposicao.SF.zip
Publication Dates
-
Publication in this collection
27 Nov 2020 -
Date of issue
2020
History
-
Received
14 July 2020 -
Accepted
7 Oct 2020