Resumos
Cinco genótipos de amendoim de diferentes hábitos de crescimento foram submetidos a estresse hídrico moderado durante 7 dias em casa de vegetação. O plantio foi feito em vasos de 15 litros contendo solo e substrato (Plantmax®), otimizados para a cultura. O delineamento experimental adotado foi inteiramente casualizado, com esquema bi-fatorial 5 × 2 (5 genótipos × 2 tratamentos hídricos), com cinco repetições. As atividades enzimáticas da SOD, CAT, APX e GPX foram analisadas, além do acúmulo de carboidratos, amido e prolina. Em função do déficit hídrico submetido, verificou-se aumento nas atividades da SOD e da CAT em todos os genótipos, tanto nas folhas quanto na raiz, destacando-se as cv. 55 437 e BR 1 e a linhagem LBM Branco. O acúmulo de prolina e redução do teor de amido foram verificados em todos os genótipos, em resposta ao déficit hídrico, porém o teor de carboidratos foi elevado só nos materiais mais tolerantes. Em função das respostas obtidas, destacam-se como materiais mais tolerantes as cultivares 55 437 e BR 1, seguidas por LBM Branco. LBR Branco tem comportamento intermediário e LViPE 06 é o material mais sensível.
amendoim; estresse hídrico; espécies reativas de oxigênio
Five peanut genotypes, with different growth habits were submitted to moderate water stress in a greenhouse for seven days. Seeds were planted in 15-litre pots containing soil and a substrate (Plantmax®), both optimized for the crop. The adopted experimental design was completely randomized, using a 5 x 2 bi-factorial scheme(5 genotypes × 2 water treatments) with five replications. The enzymatic activities of SOD, CAT, APX and GPX were analyzed, as well as the carbohydrate, starch and proline. An increased activity was shown in SOD and CAT enzymes for all genotypes submitted to water stress, in both leaf and root, especially in the 55 437 and BR1 cultivars and also in LBM Branco line. Proline accumulation and reduced starch were verified in all stressed genotypes, but carbohydrate content was increased just in tolerant materials. Based on results, 55 437 and BR1 were more tolerant to water stress, followed by LBM Branco. The LBR Branco shows intermediate behavior and LViPE- 06 was more sensitive.
peanut; water stress; reactive oxygen species
ARTIGO CIENTÍFICO
Mudanças bioquímicas em genótipos de amendoim submetidos a déficit hídrico moderado
Biochemical changes in peanut genotypes submitted to moderate water stress
Jacqueline Wanessa de Lima PereiraI; Péricles de Albuquerque Melo FilhoI; Manoel Bandeira AlbuquerqueII; Rejane Jurema Mansur Custódio NogueiraIII; Roseane Cavalcanti SantosIV,* * Autor para correspondência
IDepartamento de Agronomia/Laboratório de Expressão Gênica/UFRPE, Rua: Dom Manoel de Medeiros, s/n, Dois Irmãos, Recife-PE, Brasil, jacquelinewlp@gmail.com, péricles@depa.ufrpe.br
IIDepartamento de Fitotecnia e Ciências Ambientais/Laboratório de Ecologia Vegetal/Centro de Ciências Agrárias/UFPB/Campus II, Areia-PB, Brasil, 58.397-000, mbalbuquerque@yahoo.com.br
IIIDepartamento de Biologia/Laboratório de Fisiologia Vegetal/UFRPE, rjmansur1@gmail.com
IVEmbrapa Algodão, Caixa Postal 174, Campina Grande-PB, Brasil, 58.107-720, caval@cnpa.embrapa.br
RESUMO
Cinco genótipos de amendoim de diferentes hábitos de crescimento foram submetidos a estresse hídrico moderado durante 7 dias em casa de vegetação. O plantio foi feito em vasos de 15 litros contendo solo e substrato (Plantmax®), otimizados para a cultura. O delineamento experimental adotado foi inteiramente casualizado, com esquema bi-fatorial 5 × 2 (5 genótipos × 2 tratamentos hídricos), com cinco repetições. As atividades enzimáticas da SOD, CAT, APX e GPX foram analisadas, além do acúmulo de carboidratos, amido e prolina. Em função do déficit hídrico submetido, verificou-se aumento nas atividades da SOD e da CAT em todos os genótipos, tanto nas folhas quanto na raiz, destacando-se as cv. 55 437 e BR 1 e a linhagem LBM Branco. O acúmulo de prolina e redução do teor de amido foram verificados em todos os genótipos, em resposta ao déficit hídrico, porém o teor de carboidratos foi elevado só nos materiais mais tolerantes. Em função das respostas obtidas, destacam-se como materiais mais tolerantes as cultivares 55 437 e BR 1, seguidas por LBM Branco. LBR Branco tem comportamento intermediário e LViPE 06 é o material mais sensível.
Palavras-chave: amendoim. estresse hídrico. espécies reativas de oxigênio.
ABSTRACT
Five peanut genotypes, with different growth habits were submitted to moderate water stress in a greenhouse for seven days. Seeds were planted in 15-litre pots containing soil and a substrate (Plantmax®), both optimized for the crop. The adopted experimental design was completely randomized, using a 5 x 2 bi-factorial scheme(5 genotypes × 2 water treatments) with five replications. The enzymatic activities of SOD, CAT, APX and GPX were analyzed, as well as the carbohydrate, starch and proline. An increased activity was shown in SOD and CAT enzymes for all genotypes submitted to water stress, in both leaf and root, especially in the 55 437 and BR1 cultivars and also in LBM Branco line. Proline accumulation and reduced starch were verified in all stressed genotypes, but carbohydrate content was increased just in tolerant materials. Based on results, 55 437 and BR1 were more tolerant to water stress, followed by LBM Branco. The LBR Branco shows intermediate behavior and LViPE- 06 was more sensitive.
Key words: peanut. water stress. reactive oxygen species.
INTRODUÇÃO
O déficit hídrico é um dos tipos de estresse abiótico mais crítico e comum que acometem as lavouras conduzidas em ambientes semiáridos. Seus efeitos são evidentes em qualquer estágio fenológico da planta, podendo variar de acordo com a severidade e duração do estresse (FAROOQ et al., 2009).
Em resposta à falta de água, as plantas desengatilham vários eventos fisiológicos sendo mais comum o ajustamento osmótico onde elas se ajustam para manter o potencial hídrico e a turgescência das células próximos ao nível adequado. Tais processos são conseguidos por meio do acúmulo de solutos orgânicos de baixo peso molecular no citosol, destacando-se açúcares solúveis, prolina e aminoácidos livres (NEPOMUCENO et al., 2001).
Independentemente do tipo de metabolismo, a falta de água leva à redução na atividade fotossintética e aumento na respiração das plantas o que, por sua vez, leva a uma produção excessiva de espécies reativas de oxigênio (Reactive Oxygen Species - ROS). Moléculas de ROS, como o oxigênio singlet (1O2), peróxido de hidrogênio (H2O2), superóxido (O2-), e radicais hidroxila (HO) produzidos nas mitocôndrias, cloroplastos e peroxissomas são capazes de causar danos oxidativos aos lípidios, proteínas e ao DNA (APEL; HIRT, 2004; MØLLER; JENSEN; HANSSON, 2007).
Para minimizar os efeitos citotóxicos dos ROS, as plantas desengatilham um complexo sistema antioxidativo onde enzimas específicas atuam neutralizando a ação desses radicais, iniciando com a superóxido dismutase (SOD. EC 1.15.1.1), que desmuta o radical O2- para H2O2; este, por sua vez, sofre ação de várias enzimas como: a catalase (CAT. EC 1.11.1.6), responsável pela converção do H2O2 a H2O e O2, e as peroxidases, ascorbato peroxidase (APX. EC 1.11.1.11) e guaiacol peroxidase (GPX. EC 1.11.1.7) que reduzem o H2O2 a H2O (APEL; HIRT, 2004).
O amendoim (Arachis hypogaea L.) é uma leguminosa conhecida por sua larga adaptação ambiental, conferida pela plasticidade genética das cultivares, especialmente as desenvolvidas para o ambiente semiárido (SANTOS; GODOY; FAVERO, 2005). Para a região Nordeste do Brasil, onde a agricultura familiar é o modelo agrícola mais expressivo, o manejo do amendoim configura-se em uma atraente alternativa, especialmente por ser uma oleaginosa e poder agregar mais um produto no emergente mercado de óleos vegetais para o segmento de biocombustíveis.
Para contribuir com o estabelecimento da lavoura na região, contudo, torna-se necessária a identificação de genótipos que possam responder estavelmente no aspecto de produção, mesmo quando submetidos a situações de baixa e irregular disponibilidade hídrica.
Vários procedimentos são adotados por pesquisadores para identificação de genótipos tolerantes ao estresse hídrico, sendo mais comum a seleção por meio de descritores agronômicos associados a descritores fisiológicos e bioquímicos. Dentre esses últimos, os relacionados ao sistema antioxidativo e ajustamento osmótico têm contribuído substancialmente na identificação de genótipos promissores, e conseqüentemente, no progresso dos trabalhos de melhoramento da cultura para resistência à seca (AZEVEDO NETO et al., 2009; SANTOS; GODOY; FAVERO, 2005).
Baseado neste aspecto apresenta-se neste trabalho um estudo sobre a atividade de alguns descritores bioquímicos em cinco genótipos de amendoim submetidos ao déficit hídrico moderado, usando-se como base as cultivares BR 1 e 55 437, que são tolerantes ao estresse hídrico.
MATERIAL E MÉTODOS
Cinco genótipos de amendoim, com níveis diferenciados de tolerância ao estresse hídrico, foram cultivados em vasos (15 L) contendo o substrato Plantmax® complementado com 15 g de Superfosfato simples e 8 g de Cloreto de potássio, em casa de vegetação. As regas foram realizadas diariamente, mantendo-se a umidade próxima à capacidade de campo. Aos 15 dias após a emergência, os tratamentos foram diferenciados sendo considerado controle (C) as plantas mantidas com rega diária e estresse (E), as plantas submetidas a suspensão total de rega durante 7 dias.
As análises bioquímicas foram iniciadas quando as plantas sob restrição hídrica atingiram 50% do fechamento estomático (7 dias), estimado com o porômetro de equilíbrio dinâmico (LI 1600, Licor Inc.) nas faces abaxial e adaxial das folhas (DUARTE, 2008).
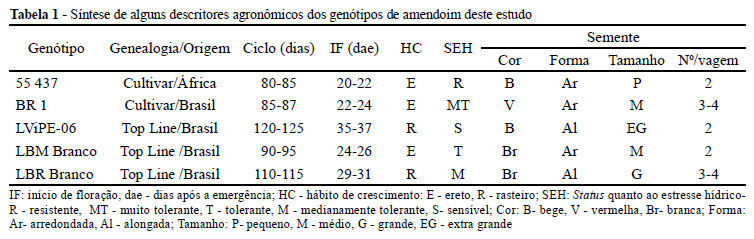
O delineamento experimental adotado foi inteiramente casualizado, com esquema bi-fatorial 5 x 2, com cinco repetições. A unidade experimental consistiu de um vaso contendo duas plantas de cada genótipo. Cada análise foi feita em folhas e raízes das plantas e em duplicada; a média de cada repetição foi utilizada para as análises estatísticas. Uma síntese dos descritores agronômicos dos materiais estudados encontra-se na Tabela 1.
Atividade enzimática
A atividade da SOD foi determinada como descrita em Giannopolitis e Ries (1977). A reação (1,5 mL) consistiu de 50 mM tampão fosfato monobásico (pH = 7,8), 0,1 mM EDTA, 13 mM metionina, 75 mM de NBT, 2 mM de riboflavina e 50 mL do extrato bruto das amostras. A reação foi exposta à luz por 15 minutos. A leitura foi feita a 560 nm, sendo que uma unidade de atividade de SOD (U) foi definida como a quantidade de enzima necessária para inibir 50% da redução do NBT e a atividade foi expressa em U g-1 MF min-1. Para análise da CAT seguiu-se a metodologia de Beers Júnior e Sizer (1952). A reação (1,5 mL) consistiu de 100 mM de tampão fosfato monobásico e 0,1 mM de EDTA (pH = 7,0), 20 mM de H2O2 e 50 µL do extrato. A atividade foi determinada pela degradação do H2O2 no intervalo de 1 minuto, a 240 nm. A quantificação foi feita adotando-se o coeficiente molar de extinção de 36 M-1 cm-1 e expressa em µmol H2O2 g-1 MF min-1.
Para APX seguiu-se a metodologia de Nakano e Asada (1981). A reação (1,5 mL) consistiu de 50 mM do tampão fosfato monobásico e 0,1 µM de EDTA (pH 6.0), 0,5 mM de ascorbato, 1 mM de H2O2 e 75 µL do extrato. A atividade foi determinada pela oxidação do ascorbato no intervalo de 1 minuto a 290 nm. Para sua quantificação utilizou-se o coeficiente molar de extinção 2,8 mM-1 cm-1 do ascorbato e expressa como µmol ascorbato g-1 MF min-1.
A atividade da GPX foi determinada como descrito por Urbanek, Kuzniak-Gebarowska e Herka (1991). A reação (1,5 mL) consistiu de 100 mM de tampão fosfato monobásico e 0,1 mM de EDTA (pH = 7,0), 5 mM de guaiacol, 15 mM de H2O2 e 50 µL do extrato. A leitura foi feita a 470 nm, no intervalo de 1 minuto. A atividade enzimática foi quantificada pela quantidade de tetraguaiacol formado usando o coeficiente extinção de 26.6 mM-1 cm-1 e os resultados foram expressos em mmol H2O2 min-1 mg-1 de matéria fresca.
Determinação de prolina
O teor de prolina foi determinado segundo metodologia de Bates (1973) em folhas e raízes. As leituras foram feitas em espectrofotômetro a 520 nm. O extrato bruto das amostras foi obtido através da maceração de 1 g de folhas completamente expandidas em 4 mL de tampão fosfato monobásico (100 mM) e EDTA (0,1 mM) (pH = 7,0). Esse extrato foi utilizado para determinação de prolina e para as análises da atividade enzimática.
Carboidratos Solúveis e Amido
Para extração de carboidratos solúveis, um grama de folhas completamente expandidas foi triturado e transferido para um tubo de ensaio contendo 5 mL de etanol 80% e aquecido a 95 ºC por 10 minutos (3 vezes). As amostras foram armazenadas em estufa (52 ºC) até a evaporação de todo líquido, sendo a seguir ressuspendidas em 10 mL de água destilada. O amido foi extraído a partir do pellet da extração de carboidratos, seguindo metodologia descrita em McCready et al. (1950). As reações para as análises de amido e carboidrato seguiram metodologia do fenol-ácido sulfúrico, descrita em Dubois et al. (1956). As leituras foram procedidas no comprimento de onda de 490 nm e a concentração dos açúcares expressa em µmol carboidratos ou amido g-1 matéria fresca.
Análise estatística
Os dados obtidos foram submetidos a ANOVA de duas vias e as médias comparadas entre si pelo teste de Tukey (P < 0,05). As análises foram realizadas com o auxílio do programa estatístico Assistat beta 7.5.
RESULTADOS E DISCUSSÃO
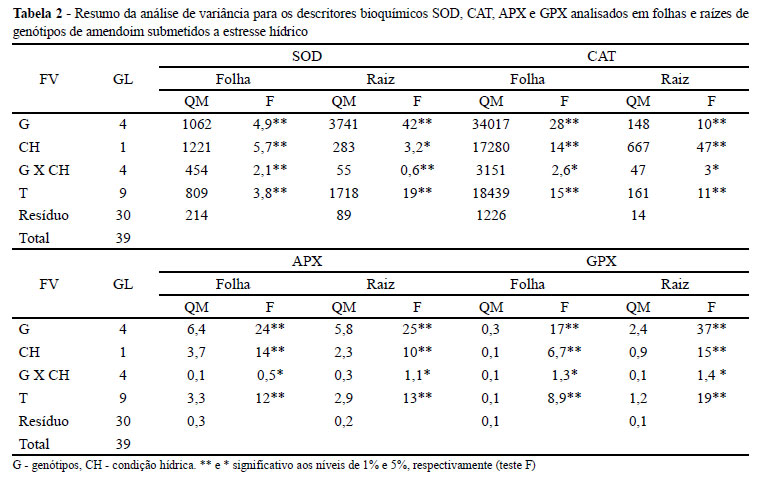
Uma síntese das análises de variância obtidas a partir dos dados das enzimas oxidativas em folhas e raízes, encontra-se na Tabela 2. Verificou-se diferença estatística significativa para todos os tratamentos e para as interações, indicando que a atividade dessas enzimas sofre alteração em decorrência do sinal de estresse hídrico engatilhado pela planta em nível celular.
A média de atividade de cada enzima nos diferentes tratamentos é apresentada nas Tabelas 3A e B. De forma generalizada, percebe-se que após a ativação da SOD nos genótipos 55 437 e BR 1, respectivamente resistente e tolerante a seca, há um aumento expressivo da CAT em ambos tecidos, quando na condição de estresse hídrico. Essa tendência, conjuntamente associada aos descritores de precocidade e ao hábito de crescimento, permite ao melhorista presumir o comportamento da planta quando a adaptação ao ambiente semiárido. Como pode ser observado na Tabela 1, ambas as cultivares são de hábito de crescimento eretos e muito precoces para iniciar a floração e finalizar o ciclo.
Akcay et al. (2010), que estudaram a atividade da CAT em acessos de amendoim rasteiro e ereto, encontraram que a atividade dessa enzima aumentou consideravelmente no acesso ereto quando submetido ao mais elevado nível de estresse, corroborando com os resultados deste trabalho. Para esses autores, a CAT é uma das enzimas mais eficazes na defesa de processos oxidativos, uma vez que, na planta resistente, possibilita a integridade da célula mesmo quando o estresse encontra-se em um estágio mais rigoroso. De acordo com Sankar et al. (2007), os níveis de expressão da CAT variam também entre os acessos eretos, sendo, contudo, sempre maior nos mais precoces, conforme pode ser visto em seu trabalho, onde um incremento médio de até 230% de atividade foi obtido no material mais precoce, quando submetido a 10 dias de supressão hídrica.
A linhagem ereta LBM também acompanha o padrão do genitor materno, denotando ter herdado o caráter de tolerância da BR 1, ao contrário da rasteira LBR Branco, também derivada do mesmo cruzamento, que apresentou um perfil intermediário entre os progenitores.
No genótipo tardio LViPE-06, as diferenças observadas entre os tratamentos hídricos para estas duas enzimas foram pouco expressivas. Tal resposta pode estar associada à baixa tolerância deste material a ambientes de baixa disponibilidade hídrica, o que é condizente com seu ciclo que, por ser mais longo, entre 120 a 125 dias, necessita de maior requerimento de água para atender todas as demandas fenológicas que envolvem as fases de crescimento e desenvolvimento.
Em trabalhos desenvolvidos por Santos et al. (2010), que avaliaram a produtividade de amendoins precoces e tardios em vários municípios do Nordeste, os autores reportaram que os genótipos tardios (hábito de crescimento rasteiro), foram de menor adaptação ao cultivo de sequeiro, cujo período chuvoso é frequentemente irregular e mal distribuído. Como tem, contudo, elevado potencial para produção de grãos, são excelentes candidatos para trabalhos de melhoramento via hibridação, utilizando-se como genitores maternos, genótipos precoces e adaptados ao semiárido.
A SOD é a primeira enzima envolvida no sistema de defesa das plantas quando a célula detecta um sinal de estresse, sendo ela responsável pela formação direta de diferentes espécies reativas de oxigênio (APEL; HIRT, 2004; SANKAR et al., 2007). Em resposta à atividade da SOD, o nível de H2O2 aumenta, engatilhando, a partir daí, a ação de enzimas secundárias de neutralização, tais como, CAT e as peroxidases, APX e GPX, que são responsáveis pela conversão do H2O2 a H2O + ½ O2 e H2O2 a H2O + R(O)2, respectivamente.
A resposta expressiva da CAT observada nos genótipos tolerantes, especialmente nas raízes, evidencia sua adoção como um descritor bioquímico nos trabalhos de seleção de materiais para tolerância ao estresse hídrico. Uma das vantagens no aspecto operacional é que a CAT exerce sua função independente da presença de agentes redutores, com atividade rápida, aproximadamente 107 min-1, capaz de neutralizar o H2O2 antes mesmo da sua difusão pela célula (AKCAY et al., 2010; SCANDALIOS, 2005). A adoção desta enzima em resposta ao estresse hídrico tem sido feita por vários autores como Terzi et al. (2010), em feijão, Sairam e Srivastava (2000), em trigo, Fazeli, Ghorbanli e Niknam (2007), em gergilim e em Sankar et al. (2007) e Akcay et al. (2010), em amendoim.
Com relação às peroxidases, outra classe de enzimas responsáveis pela neutralização do H2O2, ao contrário da CAT, elas necessitam de agentes redutores para sua atividade. A APX utiliza um elétron doado pelo ascorbato para converter o H2O2 a H2O e monodehidro-ascorbato (MDA) (APEL; HIRT, 2004), enquanto que a GPX atua oxidando uma molécula de guaiacol e reduzindo quatro moléculas de H2O2; por meio desta reação, forma-se uma molécula de tetraguaiacol com a liberação de H2O (FATIBELLO-FILHO; VIEIRA, 2002).
No presente trabalho, a atividade da GPX nas plantas sob estresse foi mais expressiva do que a APX. Nesta última, a propósito, a atividade nas folhas e raízes da LViPE-06 foi estatisticamente semelhante, podendo a APX ser utilizada como diferenciador de genótipos sensíveis ao estresse hídrico. Para confirmar tal conclusão, torna-se necessário analisar outros genótipos sensíveis, na mesma condição deste trabalho, embora Akcay et al. (2010) reportem que o aumento da atividade da CAT associado ao aumento da atividade da APX confere melhor resposta antioxidativa ao amendoim. Isso pode ser observado em todos os materiais tolerantes (55 437 e BR1) e ainda nas linhagens derivadas da BR 1, LBM Branco e LBR Branco, (Tabela 3).
Com relação aos resultados de açúcares e prolina, os valores obtidos neste trabalho encontram-se na Tabela 4. As análises foram realizadas apenas nas folhas devido às diferenças serem mais expressivas neste tecido do que em raiz, tanto em espécies de amendoim cultivado quanto silvestre, segundo trabalho realizado por Azevedo Neto et al. (2009).
Neste trabalho, verificou-se que as concentrações de carboidratos diferiram entre as condições hídricas dos tratamentos havendo ganho apenas nos genótipos 55 437, BR 1 e LBM Branco. Na análise de amido, também verificou-se diferença entre os tratamentos, contudo, todos os genótipos apresentaram expressiva redução quando as plantas estavam em condição de estresse hídrico. Na literatura, essa tendência de comportamento também ocorre em outras culturas (MAGALHÃES FILHO et al., 2008). Segundo Kramer e Boyer (1995), várias espécies quando são submetidas à seca apresentam redução na concentração de amido para formar açúcares solúveis, ou seja, a diminuição na concentração de amido é acompanhada pelo aumento da concentração de carboidratos. Melo et al. (2007), contudo, reportam que uma diminuição no teor de amido não acompanhada pelo aumento no teor de açúcares solúveis pode indicar que esteja ocorrendo o consumo imediato dos açúcares para a manutenção da sobrevivência das plantas. Pelos resultados vistos neste trabalho, essa afirmação é mais compatível com o comportamento das rasteiras LViPE-06 e da LBR Branco, que são sensíveis e medianamente sensíveis ao déficit hídrico, respectivamente (PEREIRA, 2010). Para os demais genótipos, que são mais tolerantes, a elevação nos teores de carboidratos deve está relacionada com o melhor ajustamento da regulação osmótica para manutenção dos níveis de água nas folhas visando o equilíbrio das células (MENDES et al., 2007).
Com relação à concentração de prolina livre verificou-se aumento em todas as plantas sob a condição de estresse, sendo o acúmulo maior observado na cultivar resistente 55 437 (Tabela 4), na ordem de 105%. Na cultivar BR 1 e em seus descendentes, LBM Branco e LBR Branco, os aumentos foram mais discretos, de 49; 29 e 45%, respectivamente. No genótipo mais sensível, LViPE-06, o acumulo situou-se na faixa de 36%.
Os valores obtidos para a maioria dos genótipos não foram tão expressivos como os encontrados para a cultivar 55 437, um material conhecidamente resistente à seca (AZEVEDO NETO et al., 2009; NOGUEIRA et al., 1998; PEREIRA, 2010). No trabalho de Nogueira et al. (1998), que testou esta cultivar por um período de apenas 10 dias de estresse hídrico, o acúmulo nas folhas das plantas estressadas foi de 15 vezes maior do que nas plantas controle. Esta cultivar apresentou também maior resistência difusiva e menor potencial hídrico foliar do que a cultivar a sensível, avaliada no trabalho, a IAC Tupã. Azevedo Neto et al. (2009), que também avaliaram folhas e raízes da cultivar 55 437 (Arachis hypogaea), juntamente com outras três espécies selvagens (A. duranensis, A. ipaensis e A. stenosperma), submetidas a 45 dias de estresse hídrico, encontraram acúmulo de prolina nas folhas na ordem de 233%. Segundo os autores, nas raízes também houve acúmulo porém, menos expressivo do que nas folhas.
O programa de melhoramento de amendoim desenvolvido pela Embrapa para a região semiárida utiliza frequentemente a cultivar 55 437 como progenitor nos trabalhos de hibridação visando transferência do caráter de tolerância a seca para os materiais brasileiros (SANTOS; GODOY; FAVERO, 2005). A cultivar BRS 151 L7, a mais precoce desenvolvida pela Embrapa e de elevada tolerância ao ambiente semiárido, tem a cultivar africana como um de seus progenitores (GOMES et al., 2007). O rendimento da 55 437, que pertence ao grupo botânico Spanish, não é elevado, variando entre 1.400 e 1.600 kg ha-1 (SANTOS et al., 1999). Entretanto, as linhagens dela oriundas, produzem, em média, 1.800 kg ha-1, fornecendo um incremento médio de 26% na produtividade, aliado a herança de tolerância ao ambiente semiárido (SANTOS et al., 2010).
Entre os demais genótipos avaliados neste estudo, destaca-se a BR 1, cultivar do grupo Valencia, de alta tolerância ao semiárido e de larga aceitação pelos agricultores regionais (SANTOS et al., 2010). Os genótipos dela derivadas, LBM Branco e LBR Branco, obtidos por meio de cruzamento com a rasteira LViPE-06 demonstraram certa habilidade em tolerar a condição de estresse a que foram submetidas. Contudo, o comportamento observado na rasteira LBR Branco, foi bastante expressivo quando se considera se tratar de um material rasteiro. Percebe-se neste estudo que esta linhagem apresentou maior habilidade de se ajustar ao déficit hídrico do que seu progenitor LVi-PE-06, que é mais sensível. Tal habilidade, adquirida pela herança da BR 1, a condiciona a melhor adaptação ao manejo em condições de disponibilidade hídrica limitada, o que também lhe é possibilitado devido ao ciclo mais curto, de apenas 115 dias, em comparação com as cultivares rasteiras tradicionais (130 dias). Tal precocidade permite, comparativamente, que a LBR Branco se ajuste melhor a situação de déficit hídrico, antes que uma condição de estresse mais severa se estabeleça.
Para a região Nordeste, onde há uma demanda crescente por cultivares rasteiras, esses dados são importantes, uma vez que as atuais cultivares disponíveis no mercado foram desenvolvidas para a região Sudeste e tem baixa adaptação ao ambiente semiárido (SANTOS et al., 2005). A adoção de um material do tipo da LBR Branco permite melhor competitividade para o segmento de grãos grandes, com alto teor de óleo.
CONCLUSÕES
As cultivares 55 437 e BR 1 destacaram-se como materiais mais resistentes à seca. A LBM Branco, uma linhagem gerada por cruzamento entre a BR 1 e a LViPE-06, tem comportamento semelhante a precoce BR 1; a outra isolinha, LBR Branco, tem comportamento intermediário e a tardia LViPE-06 é o material mais sensível a condição de déficit hídrico, nas condições deste trabalho.
Recebido para publicação em 04/08/2011; aprovado em 23/03/2012
pesquisa financiada pela Embrapa Algodão
Parte de Dissertação de Mestrado em Melhoramento Genético, na UFRPE
- AKCAY, U. C. et al Drought-induced oxidative damage and antioxidant responses in peanut (Arachis hypogaea L.) seedlings. Plant Growth Regulation, v. 61, n. 01, p. 21-28, 2010.
- APEL, K.; HIRT, H. Reactive oxygen species: Metabolism, oxidative stress and signal transduction. Annual Review of Plant Biology, v. 55, p. 373-399, 2004.
- AZEVEDO NETO, A. D. et al Physiological and biochemical responses of peanut genotypes to water deficit, Journal of Plant Interactions, v. 05, n. 01, p. 01-10, 2009.
- BATES, L. S. Rapid determination of free proline for water-stress studies. Plant and Soil, v. 39, n. 01, p. 205-207, 1973.
- BEERS JUNIOR, R. F.; SIZER, I. W. A spectrophotometric method for measuring the breakdown of hydrogen peroxidase by catalase. Journal of Biological Chemistry, v. 195, n. 02, p. 133-140, 1952.
- DUARTE, E. A. A. Identificação de genes diferencialmente expressos em amendoim submetido a estresse hídrico 2008. 54 f. Dissertação (Mestrado em Melhoramento Genético de Plantas) - Universidade Federal Rural de Pernambuco, Recife, 2008.
- DUBOIS, M. et al Colorimetric methods for determination of sugars and substances. Analytical Chemistry, v. 28, n. 03, p. 350-356, 1956.
- FAROOQ, M. et al Plant drought Stress: Effects, mechanisms and management. Agronomy for Sustainable and Development, v. 29, n. 01, p. 185-212, 2009.
- FATIBELLO-FILHO, O.; VIEIRA, I. C. Uso analítico de tecidos e de extratos brutos vegetais como fonte enzimática. Química Nova, v. 25, n. 03, p. 455-464, 2002.
- FAZELI, F.; GHORBANLI, M.; NIKNAM, V. Effect of drought on biomass, protein content, lipid peroxidation and antioxidant enzymes in two sesame cultivars. Biologia Plantarum v. 51, n. 01, p. 98-103, 2007.
- GIANNOPOLITIS, C. N.; RIES, S. K. Superoxide dismutases. I. Occurrence in higher plants. Plant Physiology, v. 59, n. 02, p. 309-314, 1977.
- GOMES, L. R. et al Adaptabilidade e estabilidade fenotípica de genótipos de amendoim de porte ereto. Pesquisa Agropecuária Brasileira, v. 42, n. 07, p. 985-989, 2007.
- KRAMER, P. J.; BOYER, J. S. Water relations of plants and soils Academic Press, 1995. 495 p.
- MAGALHÃES FILHO, J. R. et al Deficiência hídrica, trocas gasosas e crescimento de raízes em laranjeira "Valência" sobre dois tipos de porta enxerto. Bragantia, v. 67, n. 01, p. 75-82, 2008.
- MCCREADY, R. M. et al Determination of starch and amylose in vegetables. Analytical Chemistry v. 22, n. 09, p. 1156-1158, 1950.
- MELO, H. C. et al Alterações anatômicas e fisiológicas em Setaria anceps Stapf ex Massey e Paspalum paniculatum L. sob condições de déficit hídrico. Hoehnea, v. 34, n. 02, p. 145-153, 2007.
- MENDES, R. M. S. et al Relações fonte-dreno em feijão-de-corda submetido a deficiência hídrica. Revista Ciência Agronômica, v. 38, n. 01, p. 95-103, 2007.
- MØLLER, I. M.; JENSEN, P. E.; HANSSON, A. Oxidative modifications to cellular components in plants. Annual Review of Plant Biology, v. 58, p. 459-481, 2007.
- NAKANO, Y.; ASADA, K. Hydrogen peroxide is scavenged by ascorbate-specific peroxidases in spinach chloroplast. Plant Cell Physiology, v. 22, n. 05, p. 867-880, 1981.
- NEPOMUCENO, A. L. et al Tolerância à seca em plantas: Mecanismos fisiológicos e moleculares. Biotecnologia, Ciência & Desenvolvimento, v. 04, n. 23, p. 12-18, 2001.
- NOGUEIRA, R. J. M. C. et al Comportamento fisiológico de duas cultivares de amendoim submetidas a diferentes regimes hídricos. Pesquisa Agropecuária Brasileira, v. 33, n. 12, p. 1963-1969, 1998.
- PEREIRA, J. W. L. Respostas fisiológica e agronômica de genótipos de amendoim sob condição de estresse hídrico 2010. 64 p. Dissertação (Mestrado em Melhoramento Genético de Plantas), Universidade Federal Rural de Pernambuco, Recife, 2010.
- SAIRAM, R. K; SRIVASTAVA, G. C. Water stress tolerance of wheat (Triticum aestivum L.): Variations in hydrogen peroxide accumulation and antioxidant activity in tolerant and susceptible genotypes. Journal Agronomy & Crop Science v. 186, n. 01, p. 63-70, 2001.
- SANKAR, B. et al Effect of paclobutrazol on water stress amelioration through antioxidants and free radical scavenging enzymes in Arachis hypogaea L. Colloids and Surfaces B: Biointerfaces. v. 60, n. 2, p. 229-235, 2007.
- SANTOS, R. C. et al Estabilidade fenotípica de cultivares de amendoim avaliadas na região Nordeste do Brasil. Ciência Agrotecnologia, v. 23, n. 04, p. 808-812, 1999.
- SANTOS, R. C.; GODOY, J. I.; FÁVERO, A. P. Melhoramento do Amendoim. In: SANTOS, R. C. O Agronegócio do Amendoim no Brasil Campina Grande: Embrapa Algodão, 2005, cap. 4, p. 124-192.
- SANTOS, R. C. et al Produtividade de linhagens avançadas de amendoim em condições de sequeiro no Nordeste brasileiro. Revista Brasileira de Engenharia Agrícola e Ambiental, v. 14, n. 06, p. 589-593, 2010.
- SCANDALIOS, J. G. Oxidative stress: molecular perception and transduction of signals triggering antioxidant gene defenses. Brazilian Journal of Medical and Biological Research v. 38, n. 07, p. 995-1014, 2005.
- TERZİ, R. et al Impact of soil drought stress on photochemical efficiency of photosystem II and antioxidant enzyme activities of Phaseolus vulgaris cultivars. Tubitak v. 34, p. 01-10, 2010.
- URBANEK, H.; KUZNIAK-GEBAROWSKA, E.; HERKA, K. Elicitation of defense responses in bean leaves by Botrytis cinerea polygalacturonase. Acta Physiologiae Plantarum, v. 13, n. 01, p. 43-50, 1991.
Datas de Publicação
-
Publicação nesta coleção
25 Jun 2012 -
Data do Fascículo
Dez 2012
Histórico
-
Recebido
04 Ago 2011 -
Aceito
23 Mar 2012