ABSTRACT
This study aimed to evaluate the tolerance of C. leprosum seedlings to salt stress during the emergence and early seedling development. The experiment was carried out between the months of August and September, in a greenhouse, and the substrate used was the commercial coconut fiber wetted with aqueous solutions of sodium chloride (NaCl), calcium chloride (CaCl2) and potassium chloride (KCl) with the electrical conductivities of 0.0, 1.5, 2.5, 3.5, 4.5, 5.5 and 6.5 dS m-1. We used a completely randomized design in a 3 x 7 factorial scheme (three salts x seven levels of electrical conductivity). We evaluated seedling emergence, emergence speed index, root length, shoot length, dry matter of roots and shoots. The results indicated significant interaction between both factors analyzed. The emergence and initial growth of C. leprosum plants decrease gradually due to the increase of the electrical conductivity levels, with root length being less affected by waters salinized with CaCl and KCl. C. leprosum seedlings show sensitivity to the three types of salts and are highly sensitive to KCl and NaCl.
Key words:
Combretaceae; Salinity; Forest seeds
RESUMO
Dentre os fatores que limitam a propagação de plantas, principalmente no semiárido brasileiro, destaca-se o problema de salinidade que afeta os solos em função da dissolução e intemperismo dos minerais e do solo. Objetivou-se avaliar a emergência e crescimento inicial de plantas de C. leprosum irrigadas com águas salinas de diferentes naturezas catiônicas. O experimento foi realizado em casa de vegetação utilizando o substrato fibra de coco comercial umedecido com águas salinizadas com Na, Ca e K nas condutividades elétricas de: 0,0; 1,5; 2,5; 3,5; 4,5; 5,5 e 6,5 dS m-1. Utilizou-se o delineamento inteiramente casualizado em esquema fatorial 3 x 7 (três sais x sete níveis de condutividade elétrica). Durante vinte dias avaliou-se a emergência, índice de velocidade de emergência de plântulas, comprimento da raiz, comprimento da parte aérea, massas secas da raiz e da parte aérea. Os resultados indicaram interação significativa entre os dois fatores analisados. A emergência e crescimento inicial de plantas de C. leprosum apresentam diminuição gradativa em função do aumento dos níveis de condutividade elétrica, com o comprimento de raiz sendo menos afetado pelas águas salinizadas com CaCl e KCl. As plântulas de C. leprosum apresentam sensibilidade aos três tipos de sais, sendo verificado alta sensibilidade a KCl e NaCl.
Palavras-chave:
Combretaceae; Salinidade; Sementes florestais
INTRODUCTION
‘Mofumbo’ (Combretum leprosum Mart.) is a neotropical species with exclusive distribution in South America (LOIOLA, 2009LOIOLA, M. I. B. et al. Flora da Paraíba, Brasil: Combretaceae. Acta Botânica Brasílica, v. 23, n. 2, p. 330-342, 2009.), which is important to the whole Brazil and especially to the Caatinga biome. However, there are few studies focusing on this species and those available correspond to floristic and ethnobotanic surveys.
In relation to the agricultural exploitation of C. leprosum, as well as for most medicinal, aromatic and spice plants, information is very scarce, especially regarding production techniques (PRAVUSCHI et al., 2010PRAVUSCHI, P. R. et al. Efeito de diferentes lâminas de irrigação na produção de óleo essencial do manjericão (Ocimum basilicum L.). Acta Scientiarum. Agronomy, v. 32, n. 4, p. 687-693, 2010.).
Although the cultivation of medicinal plants has evolved in the last years, there is a limited number of species whose means of propagation are known and, therefore, preference has been given to those for which there are more advanced studies in the fields of chemistry and pharmacology. In this context, seedling production becomes one of the greatest obstacles when rational cultivation is aimed (MOREIRA et al., 2007MOREIRA, F. J. C. et al. Tratamentos pré-germinativos em sementes de Luffa cylindrica Roemer. Revista Ciência Agronômica, v. 38, n. 2, p. 233-238, 2007.). Thus, studies on propagation and conservation of plant species are of great importance, mainly those which undergo irrational exploitation.
The factors limiting propagation, particularly in the Brazilian semi-arid region, include the problem of salinity, which affects soils due to the dissolution and weathering of minerals and soil, as well as salts from irrigation water, factors that are accelerated by low rainfall and high evapotranspiration. Thus, knowing the tolerance of each species in relation to cultivation under such conditions is primordial for their propagation and perpetuation.
Tolerance to salinity is described as the ability to prevent excessive amounts of salts from the substrate from reaching the protoplasm and also to withstand toxic and osmotic effects associated with the increase in their concentration (LARCHER, 2000LARCHER, W. Ecofisiologia vegetal. Tradução de C. H. B. A. Prado. São Carlos: Rima, 2000. 531 p.). One of the most widespread methods to determine plant tolerance to excess salts is the observation of seed germination in saline substrate. Reduction in germination power, as emphasized by Andréo-Souza et al. (2010)ANDRÉO-SOUZA, Y. et al. Efeito da salinidade na germinação de sementes e no crescimento inicial de mudas de pinhão-manso. Revista Brasileira de Sementes, v. 32, n. 2, p. 83-92, 2010., occurs due to the high concentration of salts which cause stress on plants, because it reduces the osmotic potential and leads to the action of ions on cell protoplasm, negatively influencing water absorption by seeds and plants, and cause toxicity when accumulated inside plant cells (LOPES; MACÊDO, 2008LOPES, J. C.; MACÊDO, C. M. P. Germinação de sementes de couve chinesa sob influência do teor de água, substrato e estresse salino. Revista Brasileira de Sementes, v. 30, n. 3, p. 79-85, 2008.). Thus, the ability to germinate under salt stress conditions may be an indication of tolerance to salinity in the initial stage, which is one of the most critical in plant development (TAIZ; ZEIGER, 2013TAIZ, L.; ZEIGER, E. Fisiologia vegetal. 5. ed. Porto Alegre: Artmed, 2013. 918 p.).
In the literature there are no indications of studies reporting the influence of saline environments on C. leprosum, not even with different cationic nature of irrigation water. However, there are studies with other species, such as Lima; Torres (2009)LIMA, B. G.; TORRES, S. B. Estresses hídrico e salino na germinação de sementes de Zizyphus joazeiro Mart. (Rhamnaceae). Revista Caatinga, v. 22, n. 4, p. 93-99, 2009. with Zizyphus joazeiro Mart., Betoni, Scalon and Mussury (2011)BETONI, R.; SCALON, S. P. Q. E; MUSSURY, R. M. Salinidade e temperatura na germinação e vigor de sementes de mutambo (Guazuma ulmifolia lam.) (sterculiaceae). Revista Árvore, v. 35, n. 3, p. 605-616, 2011. with Guazuma ulmifolia Lam., Nogueira et al. (2012)NOGUEIRA, N. W. et al. Efeito da salinidade na emergência e crescimento inicial de plântulas de flamboyant. Revista Brasileira de Sementes, v. 34, n. 3, p. 466-472, 2012. with Delonix regia (Bojer ex Hook. Raf.), Leal et al. (2015)LEAL, C. C. P. et al. Emergence and early development of seedlings of Mimosa ophthalmocentra Mart. ex Benth. irrigated with brackish water. Bioscience Journal, v. 31, n. 3, p. 759-766, 2015. with Mimosa ophthalmocentra Mart. ex Benth. and Oliveira et al. (2018)OLIVEIRA, F. I. F. et al. Biomass and chloroplast pigments in jackfruit seedlings under saline stress and nitrogen fertilization. Revista Caatinga, v. 31, n. 3, p. 622-631, 2018. with Artocarpus heterophyllus L., in which these authors confirmed the negative influence of sodium chloride (NaCl). These studies are limited to evaluating initial development under saline water with different levels of electrical conductivity. It is necessary to study different cationic natures of irrigation water because sodium chloride is not the only salt causing negative effect on germination and initial development of seedlings, as can be observed in the study of Ferreira et al. (2013)FERREIRA, E. G. B. S. et al. Processo germinativo e vigor de sementes de Cedrela odorata L. sob estresse salino. Ciência Florestal, v. 23, n. 1, p. 99-105, 2013., who evaluated the effects of not only NaCl, but also potassium chloride (KCl) and calcium chloride (CaCl2), on the germination and vigor of Cederela odorata L.
Water of different cationic composition affects, due to the excess of salts, the external water potential and causes ionic effect due to the accumulation of ions in plant tissues (MUNNS; TESTER, 2008MUNNS, R.; TESTER, M. Mechanism of salinity tolerance. Annual Review of Plant Biology, v. 59, n. 1, p. 651-681, 2008.). When it occurs, stomatal conductance limits CO2 influx to the leaves, causing decline in the rate of assimilation of this compound (DALLAGNOL et al., 2011DALLAGNOL, L. J. et al. Alterations on rice leaf physiology during infection by Bipolaris oryzae. Australasian Plant Pathology, v. 40, n. 4, p. 360-365, 2011.). Therefore, reduction in CO2 supply to the leaves causes disorders in water relations, absorption and use of nutrients, and accumulation of potentially toxic ions, such as Na+ and Cl- in plant metabolism (AMORIM et al., 2010AMORIM, V. et al. Respostas fisiológicas de cajueiro anão precoce à salinidade. Revista Ciência Agronômica, v. 41, n. 1, p. 113-121, 2010.). The results of Lima et al. (2017)LIMA, G. S. et al. Trocas gasosas, pigmentos cloroplastídicos e dano celular na mamoneira sob diferentes composições catiônica da água. Irriga, v. 22, n. 4, p. 757-774, 2017. with castor beans, under different cationic compositions of irrigation water, highlight that potassium is the cation which leads to highest values of CO2 assimilation rate and instantaneous water use efficiency, but causes the highest percentage of damage to the cell membrane of the studied species.
Given the above, this study aimed to evaluate the effects of different cationic natures of irrigation water on the initial growth of C. leprosum plants.
MATERIAL AND METHODS
C. leprosum seeds were obtained from ripe fruits harvested from different mother plants in the municipality of Mossoró, RN, Brazil (5°12’14” S, 37°19’26” W and mean altitude of 23 m). According to Köppen’s classification, the local climate is BSwh’, dry and very hot, with two seasons: a dry one, which generally encompasses the period from June to January and rainy one, between the months of February and May (CARMO FILHO; ESPÍNOLA SOBRINHO; MAIA NETO, 1991CARMO FILHO, F.; ESPÍNOLA SOBRINHO, J.; MAIA NETO, J. M. Dados climatológicos de Mossoró: um município semi-árido nordestino. Mossoró: ESAM, 1991. 121 p. (Coleção Mossoroense. Série C, 30).).
After harvest, fruits were dried in the shade and taken to the Laboratory of Seed Analysis of the Federal Rural University of the Semi-Arid Region (UFERSA), where they were manually opened to remove the seeds. The seeds were placed in glass containers and stored in controlled environment (10 ± 2 °C and 40% relative humidity) along the entire experimental period.
The experiment was conducted in greenhouse (50% shade cloth) from August to September 2015, using polypropylene trays with 128 cells. Treatments were formed by using the substrate coconut fiber (pH = 5.4; electrical conductivity (EC) = 1.8 dS m-1; cation exchange capacity (CEC) = 92; C/N ratio = 132; d = 70 g L-1; total porosity = 95.6%; water retention = 538 mL L-1; aeration capacity = 45.5% and easily assimilable water = 19.8%), moistened with aqueous solutions of sodium (NaCl), calcium chloride (CaCl2) and potassium chloride (KCl), at the following levels of electrical conductivity: 0.0 (control), 1.5, 2.5, 3.5, 4.5, 5.5 and 6.5 dS m-1. The following evaluations were carried out:
Emergence - determined by counting the number of seedlings emerged at the end of the 20th day, considered as those whose cotyledons were exposed on substrate surface.
Emergence speed index - determined by daily counting the number of emerged seedlings, at the same time, from the first day to emergence stabilization. The index was calculated using the formula proposed by Maguire (1962)MAGUIRE, J. D. Speed of germination: aid in selection and evaluation for seedling emergence and vigour. Crop Science, v. 2, n. 2, p. 176-177, 1962..
Length of main root and hypocotyl - at the end of the emergence test (20th day), the primary root and hypocotyl of normal plants of each replicate were measured using a ruler graduated in millimeter, with results expressed in cm.
Dry matter of roots and hypocotyl of seedlings - at the end of the emergence test (20th day), roots and hypocotyl of normal seedlings were placed in Kraft paper bags and dried in a forced air circulation oven at 65 ºC until constant weight. Then, the material was weighed on analytical scale (0.001 g), and the results were expressed in mg seedling-1 (NAKAGAWA, 1999NAKAGAWA, J. Testes de vigor baseados no desempenho das plântulas. In: KRZYZANOWSKI, F. C.; VIEIRA, R. D.; FRANÇA NETO, J. B. Vigor de sementes: conceitos e testes. Londrina: ABRATES, 1999. p. 21-24.).
The experimental design was completely randomized in a 3 x 7 factorial scheme (three types of salts x seven levels of electrical conductivity), with four replicates of 25 seeds each. The results obtained were firstly subjected to exploratory analyses to evaluate the assumptions of normality of residuals, homogeneity of variances of the treatments and model additivity, and then to analysis of variance. The obtained means were compared by Tukey test (p ≤ 0.05) using the program SISVAR 5.3 (FERREIRA, 2011FERREIRA, D. O. Sisvar: a computer statistical analysis system. Ciência e Agrotecnologia, v. 35, n. 6, p. 1039-1042, 2011.). Nonlinear and polynomial regression curves, used to estimate the performance of the variables evaluated, as a function of the salt concentrations, were fitted using the software program Sigmaplot®.
RESULTS AND DISCUSSION
For all variables analyzed, there was significant interaction between factors (three types of salts and seven levels of electrical conductivity) (Table 1).
Summary of analysis of variance for emergence and the other variables of vigor of Combretum leprosum Mart. seeds, as a function of different salts and levels of electrical conductivity in irrigation water
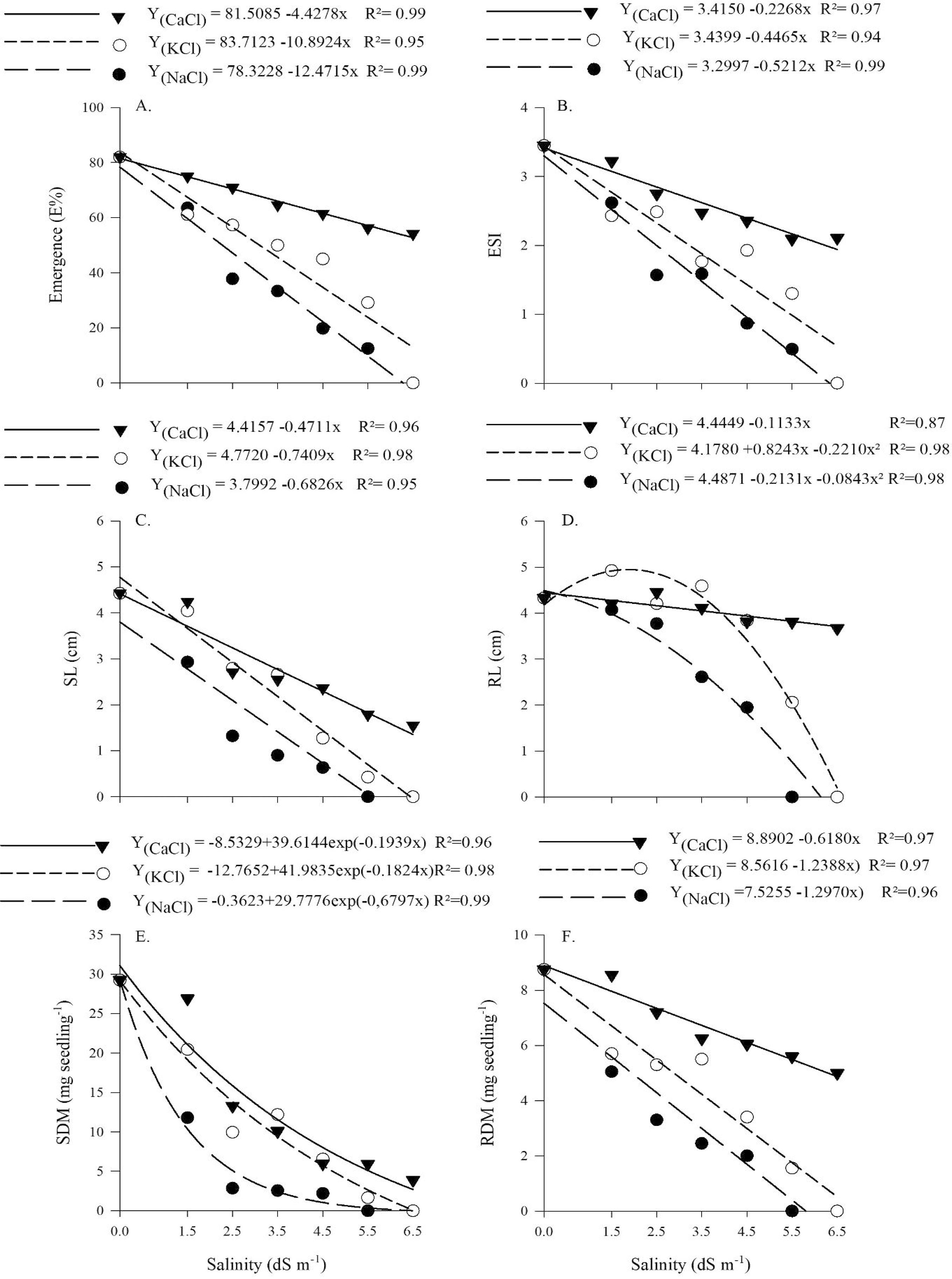
In general, according to the behavior of the regression curves (Figure 1), the water salinized with calcium chloride (CaCl2) caused smaller negative effect on the variables evaluated, whereas those salinized with NaCl resulted in greater negative effect and the one salinized with KCl led to intermediate values, compared with the other salts studied. These results do not agree with those reported by Ferreira et al. (2013)FERREIRA, E. G. B. S. et al. Processo germinativo e vigor de sementes de Cedrela odorata L. sob estresse salino. Ciência Florestal, v. 23, n. 1, p. 99-105, 2013. in Cedrela odorata L, since these authors observed more harmful effects of the concentrations of CaCl2. This demonstrates the different effect that each salt has on different species, indicating the need to evaluate separately, and not in a generalized manner, the tolerance to the cationic nature of the water based on results of different species.
Emergence (A), emergence speed index (B), shoot length (C), root length (D), shoot dry matter (E) and root dry matter (F) of Combretum leprosum Mart. seedlings subjected to three types of salts at different levels of salinity
Emergence percentage was linearly affected by the different cationic natures studied, decreasing, for all of them, with the increase in irrigation water electrical conductivity, signaling the harmful effects that the three types of salt caused on the seed. The water salinized with CaCl2 was less harmful to C. leprosum seedlings than that prepared with the other two salts (KCl and NaCl), leading to a reduction in seedling emergence from 80% to 60% between the highest and lowest levels of electrical conductivity, whereas KCl and NaCl reduced from 80 to less than 20% and 0% at the highest electrical conductivity, respectively (Figure 1A). These results corroborate those of Ferreira et al. (2013)FERREIRA, E. G. B. S. et al. Processo germinativo e vigor de sementes de Cedrela odorata L. sob estresse salino. Ciência Florestal, v. 23, n. 1, p. 99-105, 2013., in which the three salts caused negative effect on the germination and vigor of C. odorata; however, in the same study, the water salinized with CaCl2 resulted in greater negative effect compared with waters salinized with KCl and NaCl, highlighting that the negative action of the saline environments varies with species and type of salt. Therefore, successive irrigations with saline water, along with the effect of high temperature of the environment, promotes higher evaporation and results in greater accumulation of salts (MEDEIROS; SILVA; DUARTE, 2010MEDEIROS, P. R. F.; SILVA, E. F. F.; DUARTE, S. N. Salinidade em ambiente protegido. In: GHEYI, H. R.; LACERDA, C. F.; DIAS, N. S. (Ed.). Manejo da salinidade na agricultura: estudos básicos e aplicados. Fortaleza: INCTSal, 2010. p. 83-92.).
Increase in salt concentration in the substrate causes reduction in the osmotic potential, resulting in lower capacity of water absorption by seeds, which usually influences germination capacity and seedling development (LIMA et al., 2015LIMA, M. F. P. et al. Emergência e crescimento inicial de plântulas de albízia submetidas à irrigação com água salina. Revista Brasileira de Engenharia Agrícola e Ambiental, v. 19, n. 2, p. 106-112, 2015.). This toxic effect caused by the high concentration of salts leads to the action of ions on cell protoplasm, negatively influencing water absorption by seeds, and triggers the toxicity process when these salts accumulate inside plant cells (LOPES; MACÊDO, 2008LOPES, J. C.; MACÊDO, C. M. P. Germinação de sementes de couve chinesa sob influência do teor de água, substrato e estresse salino. Revista Brasileira de Sementes, v. 30, n. 3, p. 79-85, 2008.).
Studies about the effect of salts on seed germination are more common using NaCl and always show its negative effect on both emergence and initial growth, as can be observed in the studies of Leal et al. (2015)LEAL, C. C. P. et al. Emergence and early development of seedlings of Mimosa ophthalmocentra Mart. ex Benth. irrigated with brackish water. Bioscience Journal, v. 31, n. 3, p. 759-766, 2015. with Mimosa ophthalmocentra Mart. ex Benth., with accentuated reductions in seedling emergence from 1.5 dS m-1, and Nogueira et al. (2012)NOGUEIRA, N. W. et al. Efeito da salinidade na emergência e crescimento inicial de plântulas de flamboyant. Revista Brasileira de Sementes, v. 34, n. 3, p. 466-472, 2012. with Delonix regia (Bojer ex Hook.) Raf., with higher tolerance to NaCl up to the concentration of 4.5 dS m-1. Likewise, Freitas et al. (2010)FREITAS, R. M. O. et al. Efeito da irrigação com água salina na emergência e crescimento inicial de plântulas de jucá. Revista Caatinga, v. 23, n. 3, p. 54-58, 2010. observed in Caesalpinia ferrea Mart. seedlings reduction in emergence percentage, which was proportional to the increase in the levels of irrigation water electrical conductivity and with greater reduction from the NaCl concentration of 3.0 dS m-1.
Such difference in emergence percentage, obtained when seeds germinated under irrigation water of different cationic natures, occurs because as the seedlings absorb this type of water, they also end up absorbing the excess of salts which, dissolved in the saline solution, cause different damages according to the ions contained in this solution.
The emergence speed index (ESI) exhibited a similar behavior to that of seedling emergence, which was linear for the three cationic natures of the irrigation water, decreasing as the electrical conductivity increased. The control treatment showed ESI values around 3.5 with reductions of 43% (2.0) at electrical conductivity of 6.5 dS m-1 in plants irrigated using water salinized with CaCl2, and 86% (0.5) and 100% (0.0) for the cationic natures with KCl and NaCl, respectively (Figure 1B). ESI data are totally linked to emergence speed, so that higher ESI values are attributed to higher emergence speed, a desirable characteristic in the field (FERREIRA et al., 2013FERREIRA, E. G. B. S. et al. Processo germinativo e vigor de sementes de Cedrela odorata L. sob estresse salino. Ciência Florestal, v. 23, n. 1, p. 99-105, 2013.). Such reduction with increasing electrical conductivity is attributed to the increment of salt concentration in the substrate, caused by the salts dissolved in the irrigation water, which reduce the osmotic potential of the substrate, reducing water availability to the seeds, leading to decrease in water absorption capacity, and thus compromising their physiological processes and reducing emergence speed.
For shoot length, the regression curve was linear, with decrease as electrical conductivity increased. In the three cationic natures used, the best values for CaCl2 concentrations decreased from 4.5 to 1.35 cm (70% reduction) between the control treatment and the highest level of electrical conductivity. With NaCl, the highest values of length were approximately 3.8 cm, decreasing to 0 cm already at conductivity of 5.5 dS m-1, the treatment in which germination occurred but plants were stunted. The same occurred for the KCl solution of 6.5 dS m-1, in which the highest value was 4.77 in the control treatment (Figure 1C).
In study conducted with Caesalpiiniea ferrea Benth., Freitas et al. (2010)FREITAS, R. M. O. et al. Efeito da irrigação com água salina na emergência e crescimento inicial de plântulas de jucá. Revista Caatinga, v. 23, n. 3, p. 54-58, 2010. observed that seedlings irrigated with NaCl solution (0.5 dS m-1) reached average height of 6.7 cm, whereas those in the treatment with 4.5 dS m-1 were on average 5 cm tall. This demonstrates the higher tolerance to salinity exhibited by this species, which managed to maintain good seedling height even under high-salinity solutions, with a reduction of only 1.7 cm.
Although some species have good tolerance to salinity (halophytes), it is a consensus among authors that saline solutions will always cause reduction, even if it is minimal, compared to plants irrigated with normal water. This fact was verified by Holanda et al. (2007)HOLANDA, A. C. et al. Desenvolvimento inicial de espécies arbóreas em ambientes degradados por sais. Revista de Biologia e Ciências da Terra, v. 7, n. 1, p. 39-50, 2007., evaluating the initial growth of the tree species Anadenanthera colubrina, Myracroduon urundeuva, Amburana cearensis and Gliricidia sepium subjected to different levels of irrigation water salinity.
In regard to root length, it is possible to note a difference from what occurred with seedling emergence and ESI. Linear behavior was observed with a slight reduction (3%) in root length from 4.5 to 3.70 cm for waters salinized with CaCl2. The regression curve for water salinized with KCl was quadratic, with increments of 17% in root length, reaching 4.91 cm at the concentration of 1.5 dS m-1, compared with the control. For water salinized with NaCl, a similar growth was observed in the other variables, and root length decreased with increasing electrical conductivity, reaching zero at conductivity of 5.5 dS m-1, because there were no normal plants under such condition. These results indicate that C. leprosum is able to normally develop its root system and deepen the main root even in soils salinized with CaCl2, and that it may also occur in soils with KCl concentration up to 1.5 dS m-1. On the other hand, Leal et al. (2015)LEAL, C. C. P. et al. Emergence and early development of seedlings of Mimosa ophthalmocentra Mart. ex Benth. irrigated with brackish water. Bioscience Journal, v. 31, n. 3, p. 759-766, 2015., with M. ophthalmocentra, observed greater root length when seedlings were irrigated with saline solution containing NaCl at concentration of 2.5 dS m-1.
Root dry matter accumulation decreased gradually and in a more severe manner compared with the other variables. For this variable, less harmful effect was again caused by the water salinized with CaCl2, with values that reached less than 5 mg plant-1, which are much lower than those above 30 mg plant-1 found in the control treatment. For waters salinized with KCl and NaCl, dry matter values were equal to zero already at the electrical conductivity of 5.5 dSm-1, evidencing once more the toxic effect of the saline solutions on the initial growth of C. leprosum seedlings (Figure 1E and F). For shoot dry matter, an exponential effect was caused by the three cationic natures of the irrigation water, with reductions in dry matter accumulation of 46% and 99.9% for waters salinized with CaCl and KCl, respectively, at electrical conductivity of 6.5 dS m-1. In water salinized with NaCl, there was a reduction of 100% in the dry matter between the control treatment and the electrical conductivity of 6.5 dS m-1. This was also observed by Oliveira et al. (2009)OLIVEIRA, F. R. A. et al. Emergência de plântulas de moringa irrigada com água de diferentes níveis de salinidade. Bioscience Journal, v. 25, n. 5, p. 66-74, 2009., working with Moringa oleifera Lam. seedlings irrigated using saline solutions with NaCl concentrations from 0 to 5.0 dS m-1. These authors found that dry matter accumulations in the seedlings were reduced and statistically different when the level of irrigation water salinity increased.
However, Nogueira et al. (2012)NOGUEIRA, N. W. et al. Efeito da salinidade na emergência e crescimento inicial de plântulas de flamboyant. Revista Brasileira de Sementes, v. 34, n. 3, p. 466-472, 2012. in a similar study, conducted with D. regia seeds and saline solutions with NaCl concentrations ranging from zero to 6.0 dS m-1, found no difference between the values obtained in treatments in which seedlings were irrigated using solutions with NaCl concentrations of 0.5, 1.5 and 3.0 dS m-1. These authors also found that the reduction in dry matter accumulation occurred from the concentration of 4.5 dS m-1 and that the lowest value for this variable was obtained in the treatment with 6.0 dS m-1 irrigation solution.
Similarly, Silva et al. (2005)SILVA, M. B. R. et al. Crescimento de plantas jovens da espécie florestal favela (Cnidosculus phyllacanthus Pax & K. Hoffm) em diferentes níveis de salinidade da água. Revista de Biologia e Ciências da Terra, v. 5, n. 2, p. 1-13, 2005., in a study conducted with Cnidoscolus phyllacanthus (Müll. Arg.) Pax & L. Hoffm. seedlings irrigated using saline solutions with NaCl concentrations between 1.0 dS m-1 and 6.0 dS m-1, also found reduction of 63.4% in shoot dry matter when irrigation was carried out using saline solutions with concentrations between the lowest and highest levels.
In studies conducted with M. oleifera seedlings, Miranda et al. (2007)MIRANDA, J. R. P. et al. Produção de massa seca e acúmulo de nutrientes por plantas de moringa (Moringa oleifera Lam.) cultivadas em solução nutritiva com diferentes níveis de NaCl. Revista de Ciências Agrárias, n. 47, p. 187-198, 2007. observed that the gradual increase of salt concentration in the irrigation solution (0 to 150 mol m-1) caused a proportional reduction in root dry matter accumulation and that, although soil salinity was more adequate for this species at the concentration of up to 30 mol m-1, the irrigation solution with NaCl concentration of 60 mol m-1 did not cause toxic effect on either root system or normal growth of seedlings. This fact was observed in the present study in C. leprosum seedlings subjected to CaCl2 solution of 1.5 dS m-1.
CONCLUSIONS
-
C. leprosum emergence and initial growth gradually decrease with increasing levels of electrical conductivity, and root length is less affected by waters salinized with CaCl2 and KCl;
-
C. leprosum seedlings are sensitive to the three types of salts and highly sensitive to KCl and NaCl.
-
1
Parte da Tese de Doutorado do primeiro autor apresentada no Programa de Pós-Graduação em Fitotecnia da Universidade Federal Rural do Semi-Árido
ACKNOWLEDGMENTS
To the Rio Grande do Norte Research Support Foundation (FAPERN) and to the Coordination for the Improvement of Higher Education Personnel (CAPES) for granting the scholarship to the first author.
REFERENCES
- AMORIM, V. et al Respostas fisiológicas de cajueiro anão precoce à salinidade. Revista Ciência Agronômica, v. 41, n. 1, p. 113-121, 2010.
- ANDRÉO-SOUZA, Y. et al Efeito da salinidade na germinação de sementes e no crescimento inicial de mudas de pinhão-manso. Revista Brasileira de Sementes, v. 32, n. 2, p. 83-92, 2010.
- BETONI, R.; SCALON, S. P. Q. E; MUSSURY, R. M. Salinidade e temperatura na germinação e vigor de sementes de mutambo (Guazuma ulmifolia lam.) (sterculiaceae). Revista Árvore, v. 35, n. 3, p. 605-616, 2011.
- CARMO FILHO, F.; ESPÍNOLA SOBRINHO, J.; MAIA NETO, J. M. Dados climatológicos de Mossoró: um município semi-árido nordestino. Mossoró: ESAM, 1991. 121 p. (Coleção Mossoroense. Série C, 30).
- DALLAGNOL, L. J. et al Alterations on rice leaf physiology during infection by Bipolaris oryzae Australasian Plant Pathology, v. 40, n. 4, p. 360-365, 2011.
- FERREIRA, D. O. Sisvar: a computer statistical analysis system. Ciência e Agrotecnologia, v. 35, n. 6, p. 1039-1042, 2011.
- FERREIRA, E. G. B. S. et al Processo germinativo e vigor de sementes de Cedrela odorata L. sob estresse salino. Ciência Florestal, v. 23, n. 1, p. 99-105, 2013.
- FREITAS, R. M. O. et al Efeito da irrigação com água salina na emergência e crescimento inicial de plântulas de jucá. Revista Caatinga, v. 23, n. 3, p. 54-58, 2010.
- HOLANDA, A. C. et al Desenvolvimento inicial de espécies arbóreas em ambientes degradados por sais. Revista de Biologia e Ciências da Terra, v. 7, n. 1, p. 39-50, 2007.
- LARCHER, W. Ecofisiologia vegetal Tradução de C. H. B. A. Prado. São Carlos: Rima, 2000. 531 p.
- LEAL, C. C. P. et al Emergence and early development of seedlings of Mimosa ophthalmocentra Mart. ex Benth. irrigated with brackish water. Bioscience Journal, v. 31, n. 3, p. 759-766, 2015.
- LIMA, B. G.; TORRES, S. B. Estresses hídrico e salino na germinação de sementes de Zizyphus joazeiro Mart. (Rhamnaceae). Revista Caatinga, v. 22, n. 4, p. 93-99, 2009.
- LIMA, G. S. et al Trocas gasosas, pigmentos cloroplastídicos e dano celular na mamoneira sob diferentes composições catiônica da água. Irriga, v. 22, n. 4, p. 757-774, 2017.
- LIMA, M. F. P. et al Emergência e crescimento inicial de plântulas de albízia submetidas à irrigação com água salina. Revista Brasileira de Engenharia Agrícola e Ambiental, v. 19, n. 2, p. 106-112, 2015.
- LOIOLA, M. I. B. et al Flora da Paraíba, Brasil: Combretaceae. Acta Botânica Brasílica, v. 23, n. 2, p. 330-342, 2009.
- LOPES, J. C.; MACÊDO, C. M. P. Germinação de sementes de couve chinesa sob influência do teor de água, substrato e estresse salino. Revista Brasileira de Sementes, v. 30, n. 3, p. 79-85, 2008.
- MAGUIRE, J. D. Speed of germination: aid in selection and evaluation for seedling emergence and vigour. Crop Science, v. 2, n. 2, p. 176-177, 1962.
- MEDEIROS, P. R. F.; SILVA, E. F. F.; DUARTE, S. N. Salinidade em ambiente protegido. In: GHEYI, H. R.; LACERDA, C. F.; DIAS, N. S. (Ed.). Manejo da salinidade na agricultura: estudos básicos e aplicados. Fortaleza: INCTSal, 2010. p. 83-92.
- MIRANDA, J. R. P. et al Produção de massa seca e acúmulo de nutrientes por plantas de moringa (Moringa oleifera Lam.) cultivadas em solução nutritiva com diferentes níveis de NaCl. Revista de Ciências Agrárias, n. 47, p. 187-198, 2007.
- MOREIRA, F. J. C. et al Tratamentos pré-germinativos em sementes de Luffa cylindrica Roemer. Revista Ciência Agronômica, v. 38, n. 2, p. 233-238, 2007.
- MUNNS, R.; TESTER, M. Mechanism of salinity tolerance. Annual Review of Plant Biology, v. 59, n. 1, p. 651-681, 2008.
- NAKAGAWA, J. Testes de vigor baseados no desempenho das plântulas. In: KRZYZANOWSKI, F. C.; VIEIRA, R. D.; FRANÇA NETO, J. B. Vigor de sementes: conceitos e testes. Londrina: ABRATES, 1999. p. 21-24.
- NOGUEIRA, N. W. et al Efeito da salinidade na emergência e crescimento inicial de plântulas de flamboyant. Revista Brasileira de Sementes, v. 34, n. 3, p. 466-472, 2012.
- OLIVEIRA, F. I. F. et al Biomass and chloroplast pigments in jackfruit seedlings under saline stress and nitrogen fertilization. Revista Caatinga, v. 31, n. 3, p. 622-631, 2018.
- OLIVEIRA, F. R. A. et al Emergência de plântulas de moringa irrigada com água de diferentes níveis de salinidade. Bioscience Journal, v. 25, n. 5, p. 66-74, 2009.
- PRAVUSCHI, P. R. et al Efeito de diferentes lâminas de irrigação na produção de óleo essencial do manjericão (Ocimum basilicum L.). Acta Scientiarum. Agronomy, v. 32, n. 4, p. 687-693, 2010.
- SILVA, M. B. R. et al Crescimento de plantas jovens da espécie florestal favela (Cnidosculus phyllacanthus Pax & K. Hoffm) em diferentes níveis de salinidade da água. Revista de Biologia e Ciências da Terra, v. 5, n. 2, p. 1-13, 2005.
- TAIZ, L.; ZEIGER, E. Fisiologia vegetal 5. ed. Porto Alegre: Artmed, 2013. 918 p.
Publication Dates
-
Publication in this collection
Apr-Jun 2019
History
-
Received
15 Aug 2016 -
Accepted
07 June 2018