Abstract
INTRODUCTION: Honey is a common household product with many medicinal uses described in traditional medicine. Only recently has its antioxidant properties and preventive effects against disease been highlighted. Chrysin is a natural flavone commonly found in honey that has been shown to be an antioxidant agent. In this study, we investigated the antiproliferative and apoptotic effects of honey and chrysin on cultured human prostate cancer cells. METHODS: Cells were cultured in RPMI medium and treated with different concentrations of honey and chrysin for three consecutive days. Cell viability was quantitated by the 3-(4, 5-Dimethylthiazol-2-yl)-2, 5-diphenyl tetrazolium bromide (MTT) assay. The percentage of apoptotic cells was determined by flow cytometry using Annexin V-fluorescein isothiocyanate. RESULTS: The MTT assay revealed that both compounds had an antiproliferative effect on PC-3 cells in a dose- and time-dependent manner. The IC50 values for honey and chrysin against PC-3 cells were 2.5% and 24.5% after 48 h and 1.8% and 8.5% after 72 h, respectively. Chrysin induced apoptosis in PC-3 cells, as determined by flow cytometry. CONCLUSION: Our results suggest that honey has anti-proliferative effects on prostate cancer cells and the effects are mainly due to chrysin. Therefore, chrysin may be a potential compound for both cancer prevention and treatment. Further in vivo investigation is needed to support the use of chrysin in cancer therapy.
Apoptosis; Cytotoxicity; Chrysin; Honey; PC-3
BASIC RESEARCH
IDepartment of Physiology, School of Medicine, Mashhad University Medical Sciences, Mashhad, Iran
IIImmunology Research Center, BuAli Research Institute, Mashh University of Medical Sciences, Mashhad, Iran
ABSTRACT
INTRODUCTION: Honey is a common household product with many medicinal uses described in traditional medicine. Only recently has its antioxidant properties and preventive effects against disease been highlighted. Chrysin is a natural flavone commonly found in honey that has been shown to be an antioxidant agent. In this study, we investigated the antiproliferative and apoptotic effects of honey and chrysin on cultured human prostate cancer cells.
METHODS: Cells were cultured in RPMI medium and treated with different concentrations of honey and chrysin for three consecutive days. Cell viability was quantitated by the 3-(4, 5-Dimethylthiazol-2-yl)-2, 5-diphenyl tetrazolium bromide (MTT) assay. The percentage of apoptotic cells was determined by flow cytometry using Annexin V-fluorescein isothiocyanate.
RESULTS: The MTT assay revealed that both compounds had an antiproliferative effect on PC-3 cells in a dose- and time-dependent manner. The IC50 values for honey and chrysin against PC-3 cells were 2.5% and 24.5% after 48 h and 1.8% and 8.5% after 72 h, respectively. Chrysin induced apoptosis in PC-3 cells, as determined by flow cytometry.
CONCLUSION: Our results suggest that honey has anti-proliferative effects on prostate cancer cells and the effects are mainly due to chrysin. Therefore, chrysin may be a potential compound for both cancer prevention and treatment. Further in vivo investigation is needed to support the use of chrysin in cancer therapy.
Keywords: Apoptosis; Cytotoxicity; Chrysin; Honey; PC-3.
INTRODUCTION
Prostate cancer is the second most prevalent malignancy and the second leading cause of cancer-related deaths among men in the United States, with similar trends in many western countries.1 Prostate cancer is usually diagnosed in men over 50 years of age and with life expectancy increasing worldwide the incidence of prostate cancer is also likely to increase.1,2 The fine balance between cell proliferation and apoptotic death is lost in cancer, contributing to increased cellular mass and tumor progression. In terms of prostate cancer chemoprevention, there is considerable emphasis on identifying novel natural products that selectively induce apoptosis and growth arrest in prostate cancer cells without cytotoxic effects in normal cells.3,4
Foods rich in antioxidants have preventive effects against cancer, coronary disease, inflammatory disorders, and neurological degeneration.5,6 One such food includes honey,7,8 which has been used as a traditional food source since ancient times. Honey is made when nectar and sweet deposits from plants are gathered, modified, and stored in a honeycomb by honeybees. Its major components are fructose and glucose and it also consists of carbohydrates, proteins, amino acids, vitamins, water, minerals, and enzymes. Honey exhibits a broad spectrum of therapeutic properties, including antibacterial, antifungal, cytostatic,9,10 wound healing,11 and anti-inflammatory activities.12 There are also many reports in the medical literature of honey being effective as a dressing for wounds,13,14 burns,15-17 and ulcers.18,19 Honey has been to treat Fournier's gangrene, abdominal wound disruption, gastric ulcers,20 gastroenteritis, and burns and to store skin grafts.21 Honey seems to have no harmful effects and actually enables faster healing of wounds by promoting the formation of new tissues.
It was recently shown that honey could induce apoptosis in T24, RT4, 253J, and MBT-2 bladder cancer cell lines. In addition, honey significantly inhibition proliferation in T24 and MBT-2 cell lines by 1-25% and in RT4 and 253J cell lines by 6-25%.22 Honey possesses moderate antitumor and pronounced antimetastatic effects, as demonstrated in five different strains of tumors in rat and mouse models.23
Some minor constituents of honey are believed to have antioxidant properties,24 including flavonoids and phenolic acids,25,26 certain enzymes (glucose oxidase, catalase), ascorbic acid,27 carotenoid-like substances,28 amino acids, and proteins.29 Therefore, it is possible that the antitumor properties of honey are attributable to its polyphenols. With the evolution of extraction procedures for various polyphenols, it is possible to concentrate and study the polyphenolic compounds extracted from honey rather than crude honey itself.
Phenolic compounds, or polyphenols, are an important group of compounds that occur in plants and comprise at least 8,000 different known structures.30 They are also produced by plants as secondary metabolites and are present in honey. Phenolic compounds can be divided into at least 10 different types based upon their structure: simple phenols, phenolic acids, coumarins, isocoumarins, naphthoquinones, xanthones, stilbenes, anthaquinones, flavonoids, and lignins. Flavonoids are the most important polyphenolic class, with more than 5,000 compounds already described. Flavonoids are natural antioxidants that exhibit a wide range of biological effects, including antibacterial, anti-inflammatory, antiallergic, antithrombotic, and vasodilatory actions.31 Chrysin (5,7-dihydroxyflavone) is a natural and biologically active flavone extracted from honey, plants, and propolis. It has potent anti-inflammatory and antioxidant properties and promotes cell death by perturbing cell cycle progression.
Since honey is a common food for humans, we investigated whether it is a potential candidate for prostate cancer treatment by analyzing the cytotoxic and apoptotic effects of it and one of its phenolic compounds, chrysin, on prostate cancer cell lines.
MATERIAL AND METHODS
Chemicals and reagents
MTT (3-(4, 5-Dimethylthiazol-2-yl)-2, 5-diphenyltetrazolium bromide) was purchased from Amerso (USA). RPMI 1640 was purchased from Gibco BRL (Grand Island, NY, USA). Annexin V/FITC (fluorescein isothiocyanate) was obtained from Invitrogen Corporation (USA). Fetal bovine serum was purchased from PAA Laboratories GmbH, Austria. Chrysin (5,7-dihydroxyflavone), was purchased from Sigma-Aldrich (St. Louis, MO).
Cell culture
The human prostate cancer cell line PC-3 was obtained from the Pasteur Institute of Iran and cultured in RPMI medium supplemented with 10% FBS, 100 U/ml penicillin, and 100 μg/ml streptomycin. PC-3 cells were cultured in a CO2 incubator (MCO-17AI, Sanyo Electric Co., Ltd., Japan) at 37°C in a humidified atmosphere enriched by 5% CO2 and subcultured every 3-4 days.
Cell viability assay
Cell viability was measured using the MTT assay, which is based on the conversion of MTT to formazan crystals by mitochondrial dehydrogenases. Briefly, PC-3 cells were plated at a density of 1x103 cells/ml in 96-well plates and allowed to attach for 24 h, resulting in log phase growth at the time of drug treatment. Honey (2.5, 5, 10, and 20%) and chrysin (10, 20, 30, and 40 |mM) were added to the wells for 24, 48, and 72 hours. After treatment, 10 ml MTT was added to each well. After 4 h of incubation at 37°C, the solution was removed and the produced formazan was solubilized in 100 ml of dimethyl sulfoxide (DMSO). Absorbance was measured at 550 nm using an automated microplate reader (Bio-Rad 550). Cell viability was expressed as a percentage of the value for control cultures. The cytotoxic effects of honey and chrysin on PC-3 were expressed as IC50 values (the drug concentration that reduced the absorbance of treated cells by 50% compared to untreated cells). All experiments were carried out in triplicate.
Morphological studies
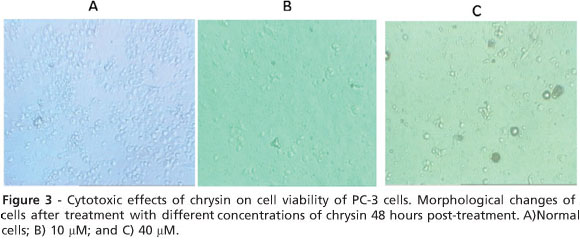
Morphological changes consistent with cell death were observed by using a normal inverted microscope. Cells were treated for 48 h with chrysin (10 and 40 mM). Untreated cells were used for a negative control. The morphological changes of the cells were observed 48 h post-treatment.
Assessment of apoptosis by Annexin V-FITC
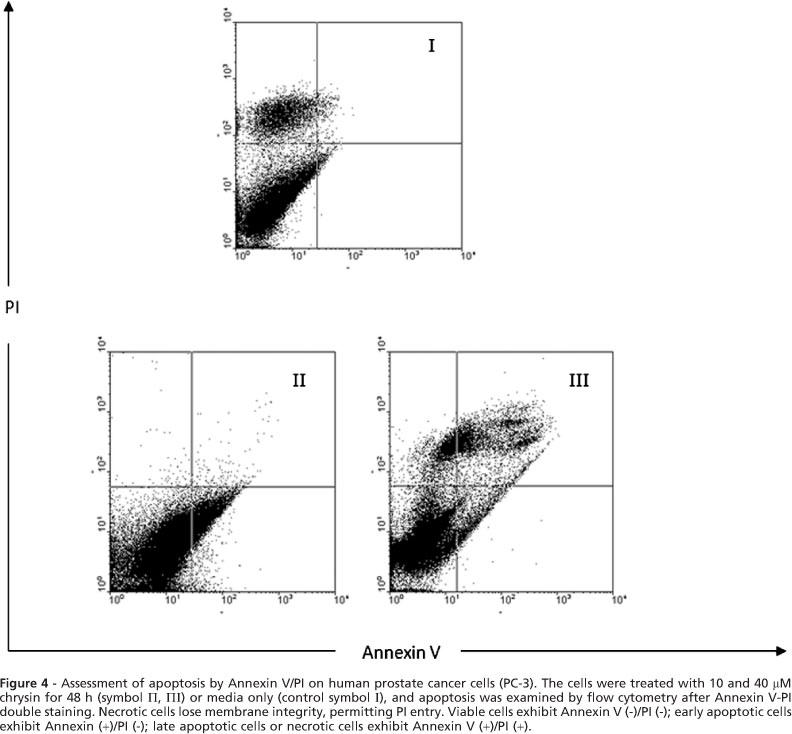
Apoptotic cell death was measured using a flouresin isothiocynate (FITC)-conjugated Annexin V/PI assay kit by flow cytometry. Briefly, 5x 105 cells were washed with ice-cold PBS, resuspended in 100 μl binding buffer, and stained with 5 μl of FITC- conjugated Annexin V (10 mg/ml) and 10 μl of PI (50 mg/ml). The cells were incubated for 15 min at room temperature in the dark, 400 μl of binding buffer was added, and the cells were analyzed (FACScan, Becton-Dickinson, USA). The PC-3 cells were gated separately according to their granularity and size on forward scatter (FSC) versus Side Scatter (SSC) plots. Early and late apoptosis was evaluated on fluorescence 2 (FL2 for propidium iodide) versus fluorescence 1 (FL1 for Annexin) plots. Cells stained with only annexin V were evaluated as being in early apoptosis; cells stained with both annexin V and propidium iodide were evaluated as being in late apoptosis or in a necrotic stage.
Statistical analysis
All results were expressed as mean + SEM. Significance was evaluated using ANOVA and Bonferroni's test. A probability level of p<0.05 was considered statistically significant.
RESULTS
Effects of honey on cell viability
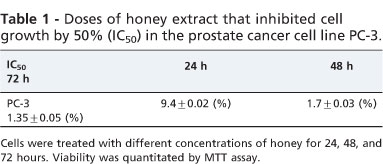
PC-3 cells were incubated with various concentrations of honey for 24, 48, and 72 h. The impact of honey on cell viability was quantitated 48 h post-exposure by the MTT assay. Exposure of PC-3 cells to honey significantly inhibited growth in a dose- and time-dependent manner (p<0.001). While there was no significant effect from the low concentration of honey after 24 hours (2.5%), there were significant decreases in viability at concentrations of 5%, 10%, and 20% (p<0.05, p<0.01, and p<0.001, respectively, Fig. 1). Treatment of PC-3 cells for 48 and 72 h resulted in significant reductions in the number of viable cells at all doses (2.5, 5,10, and 20%, p<0.001, Fig. 1). The IC50 dose for PC-3 cells was determined to be 2.5+ 0.02% and 1.8+ 0.03% at 48 and 72 h, respectively (Table 1).
Effects of chrysin on cell viability
The PC-3 carcinoma cell line was incubated with various concentrations of chrysin for 24, 48, and 72 h. The cytotoxicity of chrysin on cell viability was quantitated by MTT assay. After 24 h of exposure to chrysin, significant inhibitory effects on proliferation were only observed for the highest concentration (40 μM, p<0.05). However, there were significant decreases in viability for the concentrations of 10, 20, 30, and 40 μM after 48 and 72 h (p<0.001, Fig. 2). The IC50 dose was determined to be 24.5+0.08 | M and 8.5+0.01 μM at 48 and 72 h, respectively (Table 2).
Morphological evaluation
Although there were no significant morphological changes after 24 h of incubation with the low dose of chrysin (10 μ M, data not shown), there were significant changes after 48 h of incubation with concentrations of 10 and 40 μ M of chrysin (Fig. 3). Cells treated with chrysin (40 μ M) showed more prominent growth inhibition and cell shrinkage compared to cells treated with the lower dose of chrysin (10 μ M), consistent with the MTT results (Fig. 3).
Quantification of apoptosis by chrysin
PC-3 cells were treated with 10 and 40 μ M chrysin for 48 h to determine whether chrysin induced apoptosis. After treatment, the cells were harvested and apoptosis was examined by flow cytometry (Fig. 4). Quantitative analysis using the Annexin V/PI assay showed that the proportion of early stage apoptotic cells (Annexin V+/PI-) increased significantly from 1.6% to 28.32% and the proportion of late stage apoptotic cells (Annexin V+/PI+) increased significantly from 1.5% to 36.7% (Fig. 5). Apoptosis induced by 10 and 40 μ M of chrysin was significantly greater than in untreated cells and the percentage of early and late apoptotic cells significantly increased with chrysin concentration (p<0.001).
DISCUSSION
In recent years, considerable efforts have been made to identify naturally occurring compounds and related synthetic agents that can prevent the development and recurrence of cancer. Several naturally occurring compounds, such as phenols, indoles, inositol-6-phosphate, aromatic isothiocyanates, and dithiolethiones, have been shown to inhibit several types of cancer. In addition, a wide variety of natural food and food products can induce apoptosis in various tumor cells. There is strong evidence supporting the positive role of some natural materials and medicinal plants in oncology and their ability affect all phases of the tumorigenic process.32 Therefore, it is important to screen natural products, either as crude extracts or as isolated components, for apoptotic properties to identify potential anticancer compounds. Over 60% anticancer agents currently used are derived from natural sources, including plants, marine organisms, and microorganisms33 and they offer an opportunity to study the molecular mechanisms of tumorigenesis.34
Considerable attention is currently focused on the consumption of functional foods for the protection of human health. In particular, there is intense interest in the role of dietary antioxidants, which are capable of scavenging the oxidants and free radicals responsible for initiating various diseases.35 Systematic investigations of the antioxidant properties of various foods, beverages, spices, and herbs have been performed36 and the number of papers addressing the health-protective and antioxidant characteristics of honey is increasing. Honey has been used as a natural source of sugars for a long time, as well as an important ingredient in traditional medicine due to its antimicrobial and anti-inflammatory properties.37 The health-protective and therapeutic effects of honey has been attributed to the presence of various antioxidant components, such as flavonoids, phenolic acids, organic acids, enzymes, and vitamins,38,39 while the main sugar constituents of honey, i.e., glucose and fructose, are either inhibitors or accelerators of mutagenic activity.40
Caffeic acid (3,4-dihydroxycinnamic acid) ester derivatives, which are present in honey at levels of 20-25%, are thought to exhibit a broad spectrum of activities that include tumor inhibition.41 The mechanism of the antitumor effect shown in this study is unclear. It may be related to the inhibitory effect of caffeic acid esters and flavonoid glycones on tyrosine protein kinase (TPK), lipoxygenase, and cyclooxygenase pathway metabolites. Several cellular components associated with cell proliferation, such as poly-amines and polyamine synthetic enzymes including ornithine decarboxylase, are present at high levels in normal and neoplastic cells. In addition, many kinases, such as TPK, mediate proliferative as well as metabolic signals in cells. Eicosanoids, the metabolites of arachidonic acid from the lipoxygenase and cyclooxygenase pathways, exert a variety of biological activities. Honey also contains various poly-phones that have significant antiproliferative effects on various cancer cells in vitro.41 The apoptosis-inducing potential of honey could be attributed to its phenolic constituents. Various signaling pathways, including stimulation of TNF-alpha (tumor necrosis factor-alpha) release, inhibition of cell proliferation, induction of apoptosis and cell cycle arrest, as well as inhibition of lipoprotein oxidation, mediate the beneficial effects exerted by honey and its major components, such as chrysin and other flavonoids.42
Flavonoids are a diverse family of natural phenolic compounds commonly found in fruits, vegetables, and honey. Flavonoids are generally safe and are associated with low toxicity, making them ideal candidates for cancer chemopreventive agents. Chrysin (5,7-dihydroxyflavone) is a natural flavonoid present in honey. Recently, a number of studies have shown that chrysin has multiple biological activities, such as anti-inflammatory, anti-oxidant, and anti-cancer effects.43 In our study, we found chrysin induced cytotoxicity and apoptosis in a prostate cancer cell line. Chrysin has also been shown to inhibit tumor angiogenesis in vivo, which is a key step in metastasis.44 Chrysin may inhibit chymotrypsin-like and trypsin-like proteasomes, which play an important role in regulating apoptosis and the cell cycle.45 Chrysin suppresses expression of HIF-1a in tumor cells in vitro and inhibits tumor cell-induced angiogenesis in vivo.44 Furthermore, we have shown that chrysin induces cell cycle arrest and apoptosis in a prostate cancer cell line. Li et al. attempted to further address the anti-cancer potential of chrysin by assessing the sensitization effect of chrysin on TNFα-mediated apoptotic cell death and the molecular mechanisms involved.46 Such sensitization is closely associated with its inhibitory effect on NF-kB activation, which in turn leads to reduced expression of the anti-apoptotic NFkB target gene, c-FLIP-L, one of the key anti-apoptotic genes capable of blocking caspase-8 activity. Their reveal a novel function of chrysin and enhance the value of chrysin as a useful anti-cancer agent. In addition, the sensitization effect of chrysin on TNFa-induced cell death is mainly achieved through enhanced activation of caspase 8, the initial caspase in the death receptor signaling pathway that typically induces apoptosis.46 NF-kB, one of the main molecular targets of chemopreventive phytochemicals,47 is a transcription factor involved in multiple cellular processes, including apoptosis. Chrysin also induces apoptosis in association with activation of caspase-3, which involves the inactivation of Akt or Protein kinase B (PKB) signaling and down-regulation of X-linked inhibitor of apoptosis protein (XIAP). Other studies have shown that chrysin can potentiate the cytotoxicity of anti-cancer drugs by depleting cellular GSH (glutathione), an important factor in anti-oxidant defense.48 Chrysin potentially induces p38, activating NFkB/p65 in HeLa cells.49 The MAPK p38 pathway has been implicated in the regulation of a wide spectrum of cellular processes, including cell-cycle arrest and apoptosis. Chrysin and other flavonoids extracted from Scutellaria plants (apigenin, baicalein, baicalin, scutellarein, and wogonin) showed dose-dependent inhibition of U87-MG proliferation.
While our data shows that both honey and chrysin induce cytotoxicity and apoptosis in a prostate cancer cell line, honey is more potent. It seems that the variety of polyphenols in honey may have stronger anti-proliferative effects than chrysin alone. This is the first report on honey-induced and chrysin-induced cytotoxicity and apoptosis in human prostate cancer cells. Our data confirm that honey has cytotoxic activity against human prostate cancer cells, which is consistent with previous studies that indicate that honey and its ingredients possess antitumor and antic-arcinogenic properties. The ability to induce tumor cell apoptosis is an important property in an anticancer drug that discriminates it from a toxic compound.50
Our study showed that honey and chrysin significantly inhibit proliferation in PC-3 cells in a dose-dependent manner (Fig. 1 and Fig. 2). Apoptosis induced by chrysin was confirmed by Annexin V-FITC (Fig. 4). In the present study chrysin-induced apoptosis was involved in cell death.
Apoptosis plays an essential role in the development and maintenance of homeostasis and protection against viral infection.51 Apoptosis is characterized by distinct morphological features, including chromatic condensation, cell and nuclear shrinkage, membrane blebbing, and oligo nucleosomal DNA fragmentation.52 We found that honey contains bioactive compounds that inhibit the proliferation of a human prostate cell line (PC-3) through induction of apoptosis or programmed cell death. Our results also suggest that the anti-proliferative effects of honey are mainly due to chrysin. Chrysin may therefore be considered a potential compound for both cancer prevention and treatment.
Our results provide important insight into the molecular mechanisms involved in the anti-cancer activity of chrysin. Although further studies are needed to fully elucidate the mechanism involved in cell death, honey and chrysin are promising chemotherapeutic agents in prostate cancer treatment.
ACKNOWLEDGEMENTS
The authors would like to thank Research Affairs of Mashhad University of Medical Sciences for their financial support.
Received for publication on December 23, 2010; First review completed on January 18, 2011; Accepted for publication on March 10, 2011
E-mail: samarghandians@mums.ac.ir Tel.: 98-511-8002228
- 1. Jemal A, Siegel R, Xu J, Ward E. Cancer statistics. CA Cancer J Clin. 2010;60:277-300, doi: 10.3322/caac.20073.
- 2. Rini BI, Small EJ. Prostate cancer update. Curr. Opin. Oncol. 2002;14: 28691, doi: 10.1097/00001622-200205000-00006.
- 3. Tsao AS, Kim ES, Hong WK. Chemoprevention of cancer. CA Cancer J Clin. 2004;54:150-80, doi: 10.3322/canjclin.54.3.150.
- 4. Jain MG, Hislop GT, Howe GR, Ghadirian P. Plant foods, antioxidants, and prostate cancer risk: findings from case-control studies in Canada. Nutr Cancer. 1999;34:173-84, doi: 10.1207/S15327914NC3402_8.
- 5. Wollgast J, Anklam E. Review on polyphenols in Theobroma cacao: changes in composition during the manufacture of chocolate and methodology for identification and quantification. Food Res Int. 2000; 33:423-47, doi: 10.1016/S0963-9969(00)00068-5.
- 6. Madhavi DV, Despande SS, Salunkhe DK. Food antioxidants. Dekker: New York; 1996.
- 7. Mohamed AI-Mamary. Antioxidant activity and total phenolic of different types of honey. Nutririon Research. 2002;22:1041-7, doi: 10. 1016/S0271-5317(02)00406-2.
- 8. Estevinho L, Pereira AP, Moreira L, Dias LG, Pereira E. Antioxidant and antimicrobial effects of phenolic compounds extracts of Northeast Portugal honey. Food Chem Toxicol. 2008;46:3774-9, doi: 10.1016/j.fct. 2008.09.062.
- 9. Dustmann JH. Antibacterial effect of honey. Apiacta. 1979;14:7-11.
- 10. Molan P. The antibacterial activity of honey. The nature of the antibacterial activity. Bee World. 1992;1:5-28.
- 11. Effem SEE. Clinical observations on the wound healing properties of honey. Br J Surg. 1988;75:679-81, doi: 10.1002/bjs.1800750718.
- 12. Jeddar A, Khassany A, Ramsaroop VG, Bhamjei IE, Moosa A. The antibacterial action of honey: an in vitro study. S Afr Med J. 1985;67: 257-9.
- 13. Tovey F. Honey and sugar as a dressing for wounds and ulcers. Tropical Doctor. 2000;30:1-9.
- 14. Molan PC. Potential of honey in the treatment of wounds and burns. Am J Clin Dermatol. 2001;2:13-9, doi: 10.2165/00128071-200102010-00003.
- 15. Subrahmanyam M. Topical application of honey in treatment of burns. Br J Surg. 1991;78:497-8, doi: 10.1002/bjs.1800780435.
- 16. Philips CE. Honey for burns. Gleaning in Bee Culture. 1993;61:284.
- 17. Subrahmanyam M, Sahapure AG, Nagane NS, Bhagwat VR, Ganu JV. Effects of topical application of honey on burn wound healing. Ann Burns and Fire Disasters. 2001;14:143-5.
- 18. Jull A, Walker N, Parag V, Molan P, Rodgers A. Randomized clinical trial of honey-impregnated dressings for venous leg ulcers. Br J Surg. 2008; 95:175-82, doi: 10.1002/bjs.6059.
- 19. Molan PC, Betts JA. Using honey to heal diabetic foot ulcers. Advances in Skin and Wound Care. 2008;21:313-6, doi: 10.1097/01.ASW.0000323523. 38800.9f.
- 20. Ali AT, Al-Swayeh OA, Al-Humayed MS. Natural honey prevents ischemia-reperfusion-induced gastric mucosal lesions and increased vascular permeability in rats. Eur J Gastroentero Hepatol. 1997;9:1101-5.
- 21. Subrahmanyan M. Storage of skin grafts in honey. Lancet. 1993;341: 63-6, doi: 10.1016/0140-6736(93)92547-7.
- 22. Swellam T, Miyanaga N, Onozawa M, Hattori K, Kawai K, Shimazui T, et al. Antineoplastic activity of honey in an experimental bladder cancer implantation model: in vivo and in vitro studies. Int J Urol. 2003;10:213-9, doi: 10.1046/j.0919-8172.2003.00602.x.
- 23. Gribel NV, Pashiniski VG. Antitumor properties of honey. Vopr Onkol. 1990;36:704-7.
- 24. Antony SM, Han IY, Rieck JR, Dawson PL. Antioxidative effect of maillard reaction products formed from honey at different reaction times. J Agric Food Chem. 2000;48:3985-9, doi: 10.1021/jf000305x.
- 25. Cherchi A, Spanedda L, Tuberoso C, Cabras P. Solidphase extraction and HPLC determination of organic acid in Honey. J Chromatogr B Analyt Technol Biomed Life Sci. 2002;770:77-82, doi: 10.1016/S1570-0232(01)00583-9.
- 26. Davies AMC, Harris RG. Free amino acid analysis of honeys from England and of the geographical origin of Honeys. Journal of Apicultural Research. 1982;21:168-73.
- 27. White JW, Crane E. Honey a Comprehensive Survey. Eds. Heinemann: London: UK; 1975.
- 28. Tan ST, Wilkins AL, Holland PT, McGhie TK. Geographical discrimination of honeys though the employment of sugar patterns and common chemical quality parameters. Journal of Agricultural and Food Chemistry. 1989;37:1217-21, doi: 10.1021/jf00089a004.
- 29. White JW. The protein content of honey. Journal of Apicultural Research.vol. 1978;17:234-8.
- 30. Bravo L. Polyphenols: chemistry, dietary sources, metabolism, and nutritional significance. Nutrition Reviews. 1998;56:317-33, doi: 10.1111/ j.1753-4887.1998.tb01670.x.
- 31. Cook NC, Samman S. Flavonoids-chemistry, metabolism, cardioprotective effects, and dietary sources. Journal of Nutritional Biochemistry. 1996;7:66-76, doi: 10.1016/0955-2863(95)00168-9.
- 32. Treasure T, Fallowfield L, Farewell V, Ferry D, Lees B, Leonard P, et al. Pulmonary metastasectomy in colorectal cancer: time for a trial. Eur J Surg Oncol. 2009;35:686-9.
- 33. Cragg L, Fox A, Nation K, Reid C, Anderson M. Neural correlates of successful and partial inhibitions in children: an ERP study. Dev Psychobiol. 2009;51:533-43, doi: 10.1002/dev.20391.
- 34. Amit S, Tomoo I. Mutant p53 Gain of Oncogenic Function: In Vivo Evidence, Mechanism ofAction and Its Clinical Implications. Fukuoka Acta Med. 2009;100:217-28.
- 35. Rice-Evans C A, Miller N J, Paganga G. Structure-antioxidant activity relationships of flavonoids and phenolic acids. Free Radical Biology & Medicine. 1996;20:933-56, doi: 10.1016/0891-5849(95)02227-9.
- 36. Stasko A, Polovka M, Brezova V, Biskupic S, Malik F. Tokay wines as scavengers of free radicals (an EPR study). Food Chemistry. 2006;96: 18596, doi: 10.1016/j.foodchem.2005.02.022.
- 37. Kücük M, Kolayli S, Karaoglu S, Ulusoy E, Baltaci C, Candan F. Biological activities and chemical composition of three honeys of different types from Anatolia. Food Chemistry. 2007;100:526-34, doi: 10.1016/j.foodchem.2005.10.010.
- 38. Inoue K, Murayama S, Seshimo F, Takeba K, Yoshimura Y, Nakazawa H. Identification of phenolic compound in manuka honey as specific superoxide anion radical scavenger using electron spin resonance (ESR) and liquid chromatography with coulometric array detection. Journal of the Science of Food and Agriculture. 2005;85:872-8, doi: 10.1002/jsfa. 1952.
- 39. Wang X-H, Gheldof N, Engeseth NJ. Effect of processing and storage on antioxidant capacity of honey. Journal of Food Science. 2004;69:96-101, doi: 10.1111/j.1365-2621.2004.tb09894.x.
- 40. Busserolles J, Gueux E, Rock E, Mazur A, Rayssiguier Y. Substituting honey for refined carbohydrates protects rats from hypertriglyceridemic and prooxidative effects of fructose. Nutrient Metabolism-Research Communication Journal of Nutrition. 2002;132:3379-82.
- 41. Jaganathan a SK, Mahitosh M. Honey constituents and its apoptotic effect in colon cancer cells. JAAS. 2009;1:29-36.
- 42. Woo KJ, Jeong YJ, Park JW, Kwon TK. Chrysin-induced apoptosis is mediated through caspase activation and Akt inactivation in U937 leukemia cells. Bioch Bioph Res Com. 2004;325:1215-22, doi: 10.1016/j. bbrc.2004.09.225.
- 43. Cho H, Yun CW, Park WK, Kong JY, Kim KS, Park Y, et al. Modulationof the activity of pro-inflammatory enzymes, COX-2 and iNOS, by chrysin derivatives. Pharmacol Res. 2004;49:37-43, doi: 10.1016/S1043-6618(03)00248-2.
- 44. Fu B, Xue J, Li Z, Shi X, Jiang BH, Fang J. Chrysin inhibits expression of hypoxia-inducible factor-1alpha through reducing hypoxia-inducible factor-1alpha stability and inhibiting its protein synthesis, Mol. Cancer Ther. 2007;6:220-6, doi: 10.1158/1535-7163.MCT-06-0526.
- 45. Wu YX, Fang X. Apigenin, chrysin, and luteolin selectively inhibit chymotrypsin-like and trypsin-like proteasome catalytic activities in tumor cells. Planta Med. 2010;76:128-32, doi: 10.1055/s-0029-1186004.
- 46. Li X, Huang Q, Ong CN, Yang XF, Shen HM. Chrysin sensitizes tumor necrosis factor-alpha-induced apoptosis in human tumor cells via suppression of nuclear factor-kappaB. Cancer Lett. 2010;293:109-16, doi: 10.1016/j.canlet.2010.01.002.
- 47. Surh YJ, Chun KS, Cha HH, Han SS, Keum YS, Park KK, et al. Molecular mechanisms underlying chemopreventive activities of anti-inflammatory phytochemicals: down-regulation of COX-2 and iNOS through suppression of NF-kappa B activation. Mutat Res. 2001;480: 243-68.
- 48. Li-Weber M. Targeting apoptosis pathways in cancer by Chinese medicine. Cancer Lett. 2010 Aug 2. [Epub ahead of print]
- 49. von Brandenstein MG, Ngum Abety A, Depping R, Roth T, Koehler M, Dienes HP, et al. A p38-p65 transcription complex induced by endothelin-1 mediates signal transduction in cancer cells, Biochim. Biophys. Acta. 2008;1783:1613-22, doi: 10.1016/j.bbamcr.2008.04.003.
- 50. Mao JD, Wu P, Xia XH, Hu JQ, Huang WB, Xu GQ. Correlation between expression of gastrin, somatostatin and cell apoptosis regulation gene bcl-2/bax in large intestine carcinoma. World J Gastroenterol. 2005;11:721-5.
- 51. Wyllie AH. Apoptosis (the 1992 Frank Rose Memorial Lecture). Br J Cancer. 1993;67:205-8, doi: 10.1038/bjc.1993.40.
- 52. Wyllie AH. Apoptosis and carcinogenesis. Eur J Cell Biol. 1997; 73:18997.
Chrysin reduces proliferation and induces apoptosis in the human prostate cancer cell line pc-3
Publication Dates
-
Publication in this collection
21 July 2011 -
Date of issue
2011
History
-
Received
23 Dec 2010 -
Accepted
10 Mar 2011 -
Reviewed
18 Jan 2011