ABSTRACT:
Avian pathogenic Escherichia coli (APEC) strains are isolated from lesions of poultry presenting colibacillosis, which is a disease that causes either systemic or localized clinical signs. Such strains share many characteristics with E. coli strains that cause extra-intestinal illness in humans. There is not a consensus on how to define the APEC pathotype with regard to the presence of virulence traits. On the other hand, in the past few years, five minimal predictors for APEC detection were proposed. The E. coli isolates in this work were tested through polymerase chain reaction (PCR) to the five proposed minimal predictors and cva C. The strains presenting them were categorized as potential APEC. The APEC and non-APEC categories showed high resistance (> 50%) to cephalotin, erythromycin, streptomycin, sulphametoxazol/trimethoprim, ampicillin, and amoxicillin. Potential APEC strains were significantly more resistant to cephalotin (p < 0.05) and neomcycin (p < 0.01) than non-APEC. These latter were significantly more resistant to tetracycline (p < 0.01) than the potential APEC strains. These results demonstrate that feces of poultry present E. coli strains with resistant features, showing or not the potential of causing colibacillosis in poultry. Because APEC and extra-intestinal illness in humans may be similar, these resistant strains are of interest to public health.
KEYWORDS:
Escherichia coli; avian pathogenic Escherichia coli; antimicrobial resistance, broiler chickens
RESUMO:
Cepas de Escherichia coli patogênica para aves (APEC) estão isoladas das lesões de frangos com colibacilose, uma doença que causa sinais clínicos sistêmicos ou localizados. As APEC compartilham algumas características com as cepas de Escherichia coli que produzem doenças extraintestinais nos seres humanos. Ainda não há um consenso sobre a definição de patotipos das cepas de APEC, no que diz respeito à presença das características de virulência. Entretanto, nos últimos anos, foram definidos cinco indicadores mínimos para a identificação de patotipos das cepas de APEC. Os isolados de E. coli utilizados neste trabalho foram testados por meio de reação em cadeia de polimerase (PCR) para os cinco indicadores mínimos e para cva C. Os isolados que possuíam os cinco indicadores mínimos foram definidos como potenciais cepas de APEC. As categorias APEC e não APEC apresentaram alta resistência (> 50%) à cefalotina, eritromicina, estreptomicina, sulfametoxazol mais trimetoprim, ampicilina e amoxicilina. Possíveis cepas de APEC foram significativamente mais resistentes à cefalotina (p < 0,05) e neomicina (p < 0,01) do que as cepas não-APEC. Estas foram significativamente mais resistentes à tetraciclina (p < 0,01) do que as possíveis cepas de APEC. Esses resultados demonstram que as fezes dos frangos de corte albergam cepas de E.coli com características de resistência, apresentando ou não potencialidade de causar colibacilose. Em função das características de similaridade entre APEC e doenças extraintestinais nos seres humanos, estas cepas resistentes são de interesse à saúde pública.
PALAVRAS-CHAVE:
Escherichia coli; Escherichia coli patogênica para aves; resistência a antimicrobianos; frangos
INTRODUCTION
Avian pathogenic Escherichia coli (APEC) is a large infectious agent present in the modern poultry industry worldwide. Every year, economic losses, in order of millions, due to APEC in the poultry chain (Alekshun; Levy, 1997ALEKSHUN, M.N.; LEVY, S.B. Regulation of chromosomally mediated multiple antibiotic resistance: the mar regulon. Antimicrobial Agents Chemotherapy , v.41, p.2067-2075, 1997.). The development of diseases caused by E. coli in chickens depends on the agent's interaction with the environment and the host. The relevant virulence factors of APEC include the resistance to components of the complement system, adhesins, and the ability to sequester the iron in the host blood and tissues.
Some environmental situations such as high concentrations of ammonia in the shed, poor ventilation, extreme temperatures, wet poultry litter, high density of animals, and inefficient disinfection can act as predisposing factors (Ferreira et al., 2009Ferreira, A.J.P.; Revolledo, L.; Ferreira, C.S.A. Colibacilose. In: REVOLLEDO, L. (Ed.)Patologia Aviária.Barueri: Manole, 2009. p.67-74.). APEC belongs to the extra-intestinal pathogenic E. coli category and is associated with colibacillosis. This term refers to colisepticemia, peritonitis, pneumonia, pleuropneumonia, sacculitis, pericarditis, cellulitis, swollen head syndrome, among others diseases in poultry (Barnes et al., 2008Barnes, H.J.; Nolan, L.K.; Vaillancourt, J.F. Colibacillosis. IIn: Saif, Y.M. (Ed.) Diseases of Poultry . 12th. ed. Arres, IA: Blackwell Publishing, 2008. p.691-732.). Transmission in poultry can occur either by horizontal via (contact with other birds, feces, food, air, and water) or by vertical via (during laying and salpingitis) (Nakazato et al., 2009Nakazato, G.; Campos, T.A.; Stheling, E.G.; Brocchi, M.; Silveira, W.D. Virulence factors of avian pathogenic Escherichia coli (APEC). Pesquisa Veterinária Brasileira ,v.29, p.479-486, 2009.). There is a continuous excretion of E. coli with the potential of causing diseases in birds by poultry faces, contaminating the environment and consequently other birds. Therefore, the main reservoir of E. coli. is the poultry's intestine (Rodriguez-Siek et al., 2005Rodriguez-siek, K.E.; Giddings, C.W.; Doetkott, C.; Johnson, T.J.; Nolan, L.K. Characterizing the APEC pathotype. Veterinary Research , v.36, p.241-256, 2005.).
There is not a consensus on how to define the APEC pathotype regarding its virulence traits. However, a very compressive work has shown the presence of five genes that act as minimal predictors (iut A, hly F, iss , iro N, and omp T), and can indicate if an E. coli strain has a strong potential of causing extra-intestinal diseases in birds (Johnson et al., 2008Johnson, T.J.; Wannemuehler, Y.; Doetkott, C.; Johnson, S.J.; Rosenberger, S.C.; Nolan, L.K. Identification of minimal predictors of avian pathogenic Escherichia coli virulence for use as a rapid diagnostic tool. Journal of Clinical Microbiology , v.46, p.3987-3996, 2008.). Thus, in the present work, we classified the strains containing these five traits as potential APEC. Many of them also harbor colicins (encoded by the gene cva ), which are proteins expressed by E. coli that inhibit the bacterial growth from the same or related species (Nakazato et al., 2009Nakazato, G.; Campos, T.A.; Stheling, E.G.; Brocchi, M.; Silveira, W.D. Virulence factors of avian pathogenic Escherichia coli (APEC). Pesquisa Veterinária Brasileira ,v.29, p.479-486, 2009.).
Recent studies have indicated a close relationship between APEC and E. coli isolates associated with extra-intestinal disease in humans, then a better understanding of APEC can yield benefits not only for animals' health, but also for humans' health (Johnson et al., 2008Johnson, T.J.; Wannemuehler, Y.; Doetkott, C.; Johnson, S.J.; Rosenberger, S.C.; Nolan, L.K. Identification of minimal predictors of avian pathogenic Escherichia coli virulence for use as a rapid diagnostic tool. Journal of Clinical Microbiology , v.46, p.3987-3996, 2008.). Through the continuous antimicrobial selection, microorganisms that are resistant not only to the ingested drugs, but also to other structurally unrelated drugs, may colonize the intestinal microbiota. In addition, in poultry, multidrug-resistant microorganisms can emerge after the application of sub-therapeutic amounts of tetracycline (growth promoter) in their feed (Levy; Marshall, 2004Levy, S.B.; Marshall, B. Antibacterial resistance worldwide: causes, challenges and responses. Nature Medicine Review , v.10, p.22-29, 2004.). Populations of E. coli in the gut of poultry play an important role in the colonization and spread of pathogenicity characteristics and antimicrobials resistance. Thus, this work aimed at detecting E. coli in the feces of healthy broilers, and further at characterizing the isolates, regarding their antimicrobials resistance and genetic profiles.
MATERIAL AND METHODS
Samples were collected from 80 Cobb broilers aged 21 days that represented 25% of a poultry flock. After sampling, the cloacal swabs were immediately placed in tubes containing peptone water. In the laboratory, 0.5 mL of peptone water from each tube was put in tubes with brilliant green broth (HiMedia Laboratories, India) and incubated for 24 hours at 37°C. After incubation, the content was streaked out on MacConkey agar (Oxoid, United Kingdom). Then, it was incubated for 24 hours at 37°C. The colonies (five per plate) with typical E. coli characteristics (lactose positive) were biochemically identified through the indole production, methyl red and Voges-Proskauer reactions, citrate utilization, production of urease and hydrogen sulfide (H2S) after incubation for 24 and 72 hours at 37°C (Koneman et al., 2001Koneman, E.W.; Allen, S.D.; Janda, W.M.; Schreckenberger, P.C.; Winn Jr, W.C. Diagnóstico Microbiológico . 5th. ed. Rio de Janeiro: Medsi, 2001.).
The microbial DNA template was obtained based on a thermal lysis technique (Keskimaki et al., 2001Keskimaki, M.; Ekulund, M.; Personen, H.; Heiskanen, T.; Siitonen, A. EPEC, EAEC and STEC in stool specimens: Prevalence and molecular epidemiology of isolates. Diagnostic Microbiologicand Infectious Disease , v.40, p.151-156, 2001.). Isolates identified as E. coli were subjected to polymerase chain reaction (PCR) for detection of iut A, hly F, omp T, and cva C (Johnson; Stell, 2000Johnson, J.R.; Stell, A.L. Extended virulence genotypes of Escherichia coli strains from patients with uropepsis in relation to phylogeny and host compromiso. The Journal of Infectious Diseases , v.181, p.261-272, 2000.). Furthermore, iss and iro N were detected using primers and the conditions previously described (Rodriguez-Siek et al., 2005Rodriguez-siek, K.E.; Giddings, C.W.; Doetkott, C.; Johnson, T.J.; Nolan, L.K. Characterizing the APEC pathotype. Veterinary Research , v.36, p.241-256, 2005.). The isolates were tested for resistance to ampicillin (10 µg), amoxicillin (20 µg), cephalothin (30 µg), enrofloxacin (5 µg), erythromycin (15 µg), streptomycin (10 µg), gentamicin (10 µg), neomycin (30 µg), tetracycline (30 µg), nitrofurantoin (300 µg), and sulfamethoxazole + trimethoprim (1.25 + 23.75 µg). The method used was disk diffusion described by the National Committee for Clinical Laboratory Standards (NCCLS, 2003National Committee for Clinical Laboratory Standards. Performance standards for antimicrobial disk susceptibility tests . Approved standards. Document M2-A8. Wayne: NCCLS, 2003.). Fisher's exact test was employed to compare the antimicrobial resistance of potential APEC versus non-APEC.
RESULTS AND DISCUSSION
From the 80 sampled animals, it was possible to detect E. coli. in 48 (60%) of them. Five isolates of each positive animal were tested through biochemical reaction, yielding 91 E. coli strains. Table 1 presents that more than half of the isolates harbored at least one of the targeted genes. The most frequent profile found was iro N + omp T + iss + iut A + hly F + cva C, followed by omp T + iut A + hly F and omp T + hly F. Individually, the five most frequent minimal predictor genes detected was hly F (49.4%) followed by omp T (46%), iut A (43%), iss (31.7%), and iro N (22%). The individual prevalence of cva C was 18.7%. We identified 15 (16.5%) APEC isolates with at least five predictors (either with or without the additional gene cva C) in 10 animals.
Overall, the potential APEC isolates were slightly more resistant to the tested antimicrobials than the remaining isolates (non-APEC). There was absolute resistance (100%) in potential APEC isolates to cephalothin, erythromycin, streptomycin, neomycin, ampicillin, and amoxicillin while, for non-APEC, the resistance for these antimicrobials was 73.7, 96.1, 88.2, 42.1, 81.6, and 80.3% respectively. For sulphametoxazol plus trimethoprim, the potential APEC showed a resistance level of 86.7% while the non-APEC indicated 64.5%. However, for the antimicrobials enrofloxacin, nitrofurantoin, tetracycline and gentamicin, the non-APEC isolates were more resistant (27.6, 5.3, 77.6, and 10.5%, respectively) than the potential APEC (6.7, 0, 13.3, and 6.7%, respectively). Potential APEC strains were significantly more resistant to cephalotin (p < 0.05) and neomcycin (p < 0.01) than non-APEC strains. The non-APEC strains were significantly more resistant to tetracycline (p < 0.01) than the potential APEC strains.
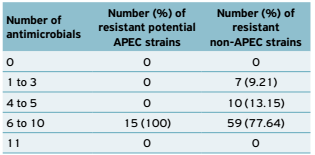
In general, all isolates were less resistant to the antimicrobials nitrofurantoin (APEC: 0%; non-APEC: 5.13%) and gentamicin (APEC: 6.7%; non-APEC: 10.5%). More differences between groups were observed for antimicrobials neomycin (APEC: 100%; non-APEC: 42.1%) and tetracycline (APEC: 13.3%; non-APEC: 77.6%). Regarding multiresistance, no isolate was sensitive or resistant to the 11 tested antimicrobials. The potential APEC showed higher multiresistance levels than non-APEC (Table 2).
In this work, it was possible to detect E. coli strains in poultry feces presenting the minimal predictors for the APEC patothype. All these strains were multiresistant to antimicrobials. Many strains without the five minimal APEC predictors were also multiresistant.
The gene related to aerobactin (iut A), which encodes a ferric aerobactin receptor, was present in 43% of the E. coli detected in this work. In a previous investigation (Wooley et al., 1992Wooley, R.E.; Spears, K.R.; Brown, J.; Nolan, L.K.; Fletcher, O.J. Relationship of complement resistance and selected virulence factors in pathogenic avian Escherichia coli Avian Disease, v.36, p.679-684, 1992.), the presence of aerobactin was reported in 95% of E. coli isolates from chickens with colisepticemia, demonstrating the probable role of such gene on this disease pathogenesis. The ability to capture iron from the environment is associated with the pathogenic ability of strains. The genes iut A and iro N (other iron uptake related gene) present in 43 and 22% of the isolates of this investigation reinforce this role, since a previous work (Silveira et al., 2002Silveira, W.D.; Ferreira, A.; Brocchi, M.; Hollanda, L.M.; Castro, A.F.P.; Yamada, A.T.; Lancellotti, M. Biological characteristics and pathogenicity of avian Escherichia coli strains. Veterinary Microbiology ,v.85, p.47-53, 2002.) showed that APEC strains may express iron sequester system, while non-pathogenic may not.
The increased serum survival (ISS) factor is associated with the pathogenicity of APEC, because it is more frequently found among pathogenic strains than in nonpathogenic ones (Pfaff-McDonough et al., 2000Pfaff-McDonough, S.J.; Horne, S.H.; Giddings, C.W.; Ebert, J.O.; Doetkott, C.; Smith, M.H.; Nolan, L.K. Complement resistance related traits among Escherichia coli isolates from apparently healthy birds and birds with colibacillosis. Avian Diseases , v.44, p.23-33, 2000.). Our results indicate the presence of iss gene in 31.7% of the isolates. Its occurrence in the plasmid ColV suggests that there is a relation between the factor ISS and APEC pathogenicity, as previously suggested (Mellata et al., 2003Mellata, M.; Dho-Moulin, M.; Dozois, C.M.; Curtiss III, R.; Lehoux, B.; Fairbrother, J.M. Role of avian pathogenic Escherichia coli virulence factors in bacterial interaction with chicken heterophils and macrophages. Infection and Immunity ,v.71, p.494-503, 2003.).
The most common genes in the strains were hly F (49.4%) and omp T (46%), which are linked to hemolytic capacity and production of proteolytic enzymes, respectively. These are important features for the colonization and establishment of ecological niches. The majority of APEC strains have a number of genes such as cvaC , iro N, iss , and iut A. Many of them are located on a plasmid known as pTJ100.
It has been suggested this group of genes might be useful in defining the APEC pathotype (Rodriguez-siek et al., 2005Rodriguez-siek, K.E.; Giddings, C.W.; Doetkott, C.; Johnson, T.J.; Nolan, L.K. Characterizing the APEC pathotype. Veterinary Research , v.36, p.241-256, 2005.). Clearly, the pathogenicity of APEC isolates reflects a coordinated action of multiple genes, frequently present in plasmids, which facilitates the ecological dispersion of pathogenic characteristics.
No single factor can be related unequivocally to APEC pathogenicity (Kariyawasam et al., 2006Kariyawasam, S.; Johnson, T.J.; Nolan, L.K. Unique DNA sequences of avian pathogenic Escherichia coli isolates as determined by genomic suppression subtractive hybridization. FEMS Microbiology Letters ,v.262, p.193-200, 2006.). Therefore, it is essential to identify microbial populations that potentially may cause intestinal imbalances and diseases affecting the health and the weight gain of poultry. Increasing resistance to first-line antimicrobial agents among E. coli isolates represents a potential threat both to animal's and human's health (Diarra et al., 2007Diarra, M.S.; Silversides, F.G.; Diarrasouba, F.; Pritchard, J.; Masson, L.; Brousseau, R.; Bonnet, C.; Delaquis, P.; Bach, S.; Skura, B.J.; Topp, E. Impact of feed supplementation with antimicrobial agents on growth performance of broiler chickens, Clostridium perfringens and Enterococcus counts, and antibiotic resistance phenotypes and distribution of antimicrobial resistance determinants in Escherichia coli isolates. Applied Environmental Microbiology , v.73, p.6566-6576, 2007.). In this study, antimicrobial resistance was more generalized among potential APEC isolates; however, there were variations, and for some antimicrobials, such as enrofloxacin, gentamicin and nitrofurantoin, the non-APEC isolates were more resistant. Furthermore, for tetracycline, the non-APEC isolates showed to be statistically more resistant than the potential APEC. The high resistance found among non-APEC isolates in this work is in accordance with a previous report (Bonnet et al., 2009Bonnet, C.; Diarrassouba, F.; Brousseau, R.; Masson, L; Topp, E.; Diarra, M.S. Pathotype and Antibiotic Resistance Gene Distributions of Escherichia coli Isolates from Broiler Chickens Raised on Antimicrobial-Supplemented Diets. Applied Environmental Microbiology ,v.75, p.6955-6962, 2009.), in which although potentially virulent E. coli isolates tended to carry few antibiotic resistance genes, broiler chickens act as a reservoir for commensal E. coli strains carrying large numbers of antibiotic resistance genes. It is noteworthy that all potential APEC strains showed multiresistance, as well as many non-APEC strains. This reinforces the possibility of E. coli of avian origin to be a reservoir of resistant genes, raising a concern in public health.
The impact of the drug selection process can be largely confined to the individual taking the antibiotic if widespread antibiotic usage is absent. After therapy, the selected resistant commensal strains will eventually be 'diluted out' and their growth will be suppressed by the return of drug-susceptible and natural competitors (Levy; Marshall, 2004Levy, S.B.; Marshall, B. Antibacterial resistance worldwide: causes, challenges and responses. Nature Medicine Review , v.10, p.22-29, 2004.). The problem of widespread resistance among bacterial populations in poultry is that they continually receive antibiotics in feed as growth promoters. If, however, whole populations are treated with the same class of antibiotic, susceptible strains will have little opportunity to recolonize their niche, and resistant strains will acquire an important advantage (Levy; Marshall, 2004Levy, S.B.; Marshall, B. Antibacterial resistance worldwide: causes, challenges and responses. Nature Medicine Review , v.10, p.22-29, 2004.). The resulting ecological imbalance produces a potentially serious environmental pool of resistant genes (Alekshun; Levy, 1997ALEKSHUN, M.N.; LEVY, S.B. Regulation of chromosomally mediated multiple antibiotic resistance: the mar regulon. Antimicrobial Agents Chemotherapy , v.41, p.2067-2075, 1997.), which eventually can be used by APEC strains.
Nonetheless, some studies have tracked a decline in resistance frequencies when an antibiotic is removed (Barbosa; Levy, 2000Barbosa, T.M.; Levy, S.B. The impact of antibiotic use on resistance development and persistence. Drug Resistant Updates , v.3, p.303-311, 2000.). Replacement by susceptible flora represents a chief contribution to a decrease in resistant strains (Levy; Marshall, 2004Levy, S.B.; Marshall, B. Antibacterial resistance worldwide: causes, challenges and responses. Nature Medicine Review , v.10, p.22-29, 2004.). Thus, one way to reduce the number of resistant strains would be to mix poultry-resistant flora with those who have sensitive flora, causing a dilution of resistant strains among populations. The findings suggest that the fastest way to eliminate resistant strains is to outnumber them with susceptible strains (Levy; Marshall, 2004Levy, S.B.; Marshall, B. Antibacterial resistance worldwide: causes, challenges and responses. Nature Medicine Review , v.10, p.22-29, 2004.).
CONCLUSIONS
The strains characterized in this work reflect a resistant and potential pathogenic population that can be spread on the environment by poultry feces, contaminating agricultural workers, agribusiness, and foods. Finally, the profile of these multiresistant isolates is of concern because APEC and human extra-intestinal illness may be similar, and so the zoonotic risk of APEC is increased when the strains show multiresistance.
ACKNOWLEDGMENTS
R.P.M. (grant number 2008/00417-0) and C.A.B. received fellowships from Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP) during the development of this work, which was funded by the same institution (grant number 2011/15050-8).
REFERENCES
- ALEKSHUN, M.N.; LEVY, S.B. Regulation of chromosomally mediated multiple antibiotic resistance: the mar regulon. Antimicrobial Agents Chemotherapy , v.41, p.2067-2075, 1997.
- Barbosa, T.M.; Levy, S.B. The impact of antibiotic use on resistance development and persistence. Drug Resistant Updates , v.3, p.303-311, 2000.
- Barnes, H.J.; Nolan, L.K.; Vaillancourt, J.F. Colibacillosis. IIn: Saif, Y.M. (Ed.) Diseases of Poultry . 12th. ed. Arres, IA: Blackwell Publishing, 2008. p.691-732.
- Bonnet, C.; Diarrassouba, F.; Brousseau, R.; Masson, L; Topp, E.; Diarra, M.S. Pathotype and Antibiotic Resistance Gene Distributions of Escherichia coli Isolates from Broiler Chickens Raised on Antimicrobial-Supplemented Diets. Applied Environmental Microbiology ,v.75, p.6955-6962, 2009.
- Diarra, M.S.; Silversides, F.G.; Diarrasouba, F.; Pritchard, J.; Masson, L.; Brousseau, R.; Bonnet, C.; Delaquis, P.; Bach, S.; Skura, B.J.; Topp, E. Impact of feed supplementation with antimicrobial agents on growth performance of broiler chickens, Clostridium perfringens and Enterococcus counts, and antibiotic resistance phenotypes and distribution of antimicrobial resistance determinants in Escherichia coli isolates. Applied Environmental Microbiology , v.73, p.6566-6576, 2007.
- Ferreira, A.J.P.; Revolledo, L.; Ferreira, C.S.A. Colibacilose. In: REVOLLEDO, L. (Ed.)Patologia Aviária.Barueri: Manole, 2009. p.67-74.
- Johnson, J.R.; Stell, A.L. Extended virulence genotypes of Escherichia coli strains from patients with uropepsis in relation to phylogeny and host compromiso. The Journal of Infectious Diseases , v.181, p.261-272, 2000.
- Johnson, T.J.; Wannemuehler, Y.; Doetkott, C.; Johnson, S.J.; Rosenberger, S.C.; Nolan, L.K. Identification of minimal predictors of avian pathogenic Escherichia coli virulence for use as a rapid diagnostic tool. Journal of Clinical Microbiology , v.46, p.3987-3996, 2008.
- Kariyawasam, S.; Johnson, T.J.; Nolan, L.K. Unique DNA sequences of avian pathogenic Escherichia coli isolates as determined by genomic suppression subtractive hybridization. FEMS Microbiology Letters ,v.262, p.193-200, 2006.
- Keskimaki, M.; Ekulund, M.; Personen, H.; Heiskanen, T.; Siitonen, A. EPEC, EAEC and STEC in stool specimens: Prevalence and molecular epidemiology of isolates. Diagnostic Microbiologicand Infectious Disease , v.40, p.151-156, 2001.
- Koneman, E.W.; Allen, S.D.; Janda, W.M.; Schreckenberger, P.C.; Winn Jr, W.C. Diagnóstico Microbiológico . 5th. ed. Rio de Janeiro: Medsi, 2001.
- Levy, S.B.; Marshall, B. Antibacterial resistance worldwide: causes, challenges and responses. Nature Medicine Review , v.10, p.22-29, 2004.
- Mellata, M.; Dho-Moulin, M.; Dozois, C.M.; Curtiss III, R.; Lehoux, B.; Fairbrother, J.M. Role of avian pathogenic Escherichia coli virulence factors in bacterial interaction with chicken heterophils and macrophages. Infection and Immunity ,v.71, p.494-503, 2003.
- Nakazato, G.; Campos, T.A.; Stheling, E.G.; Brocchi, M.; Silveira, W.D. Virulence factors of avian pathogenic Escherichia coli (APEC). Pesquisa Veterinária Brasileira ,v.29, p.479-486, 2009.
- National Committee for Clinical Laboratory Standards. Performance standards for antimicrobial disk susceptibility tests . Approved standards. Document M2-A8. Wayne: NCCLS, 2003.
- Pfaff-McDonough, S.J.; Horne, S.H.; Giddings, C.W.; Ebert, J.O.; Doetkott, C.; Smith, M.H.; Nolan, L.K. Complement resistance related traits among Escherichia coli isolates from apparently healthy birds and birds with colibacillosis. Avian Diseases , v.44, p.23-33, 2000.
- Rodriguez-siek, K.E.; Giddings, C.W.; Doetkott, C.; Johnson, T.J.; Nolan, L.K. Characterizing the APEC pathotype. Veterinary Research , v.36, p.241-256, 2005.
- Silveira, W.D.; Ferreira, A.; Brocchi, M.; Hollanda, L.M.; Castro, A.F.P.; Yamada, A.T.; Lancellotti, M. Biological characteristics and pathogenicity of avian Escherichia coli strains. Veterinary Microbiology ,v.85, p.47-53, 2002.
- Wooley, R.E.; Spears, K.R.; Brown, J.; Nolan, L.K.; Fletcher, O.J. Relationship of complement resistance and selected virulence factors in pathogenic avian Escherichia coli Avian Disease, v.36, p.679-684, 1992.
Publication Dates
-
Publication in this collection
2016
History
-
Received
02 Apr 2014 -
Accepted
31 Mar 2016