Resumo
O desmatamento elevado no Bioma Caatinga vem gerando processos de desertificação em diversas áreas, alterando diretamente a biota, o microclima e os solos, sendo fundamental o desenvolvimento de técnicas de pesquisa capazes de incorporar informações que identifiquem o estado dos recursos naturais, apontando os seus relacionamentos e alguns caminhos a serem tomados para uma intervenção eficiente que gere a recuperação e o aproveitamento sustentável das terras nesse ambiente. O MTLFC constitui-se em um conjunto de técnicas utilizado pela primeira vez no Brasil para analisar áreas submetidas à desertificação, apresentando indicadores biogeográficos, climatológicos, geomorfológicos e hidrológicos. Através da aplicação do método em uma área do município de São Domingos do Cariri (PB), foi identificada uma diversidade vegetal muito baixa (13 espécies e 489 indivíduos), dominância de poucas espécies (5) e baixa abundância, com maior número de indivíduos nos estratos arbustivo alto e arbustivo, tendo como causa a retirada excessiva de vegetação, as queimadas e o uso contínuo pelo gado caprino. O Balanço Hídrico apresentou-se negativo, havendo indicação de que a atividade vegetativa depende da precipitação oculta. Os solos apresentaram crosta superficial de origem antrópica, tornando a aridez edáfica ainda mais acentuada, associada aos baixos níveis de fertilidade, particularmente o potássio e a matéria orgânica.
Palavras-chave:
Desertificação; MTLFC; Indicadores naturais; Uso dos solos
Abstract
High deforestation in the Caatinga Biome has been generating processes of desertification in several areas, directly altering the biota, the microclimate and the soils, being essential the development of research techniques able to incorporate information which identify the state of natural resources, pointing their relationships and some paths to be taken for an effective intervention which may generate the recovery and sustainable use of land in this environment. The MTLFC consists of a set of techniques used in Brazil for the first time to analyze areas subjected to desertification, presenting biogeographic, climatological, geomorphological and hydrological indicators. By applying the method in an area of the city of São Domingos do Cariri (PB), it has been identified a very low plant diversity (13 species and 489 individuals), dominance of a few species (5) and low abundance, with the largest number of individuals in tall shrub and shrub strata, caused by the excessive removal of vegetation, fires and continuous use by the goats. The Water Balance was negative so that it indicated that the vegetative activity depends on hidden precipitation. The soils have presented surface crust of an anthropic origin, making edaphic aridity even more pronounced, associated with low fertility levels, particularly potassium and organic matter.
Key words:
Desertification; MTLFC; Natural indicators; Land Use
Resumen
La deforestación en el Bioma Caatinga ha generado procesos de desertificación en varias áreas, alterando directamente la biota, el microclima y el suelo, siendo fundamental el desarrollo de técnicas de investigación capaces de incorporar informaciones que identifiquen el estado de los recursos naturales, señalando sus relaciones y algunos caminos a seguir para una intervención efectiva que genere la recuperación y el uso sostenible de las tierras en este ambiente. El MTLFC consiste en un conjunto de técnicas utilizado por primera vez en Brasil para analizar areas desertificadas, presentando indicadores biogeográficos, climáticos, geomorfológicos e hidrológicos. Al aplicar el método en un área de São Domingos do Cariri (PB) se identificó una diversidad vegetal muy baja (13 especies y 489 individuos), el dominio de unas pocas especies (5) y baja abundancia, con mayor numero de individuos en los estratos arbustivo alto y arbustivo, teniendo como causa la eliminación excessiva de la vegetación, los incendios y el uso continuo del ganado caprino. El balance hídrico fué negativo, havendo indicios de que la actividad vegetativa depende de la precipitación oculta. Los suelos presentaron costra superficial de origen antropogénico, lo que hace la aridez edáfica aún más pronunciada asociada con los niveles de fecundidad bajos, sobre todo potasio y materia orgánica.
Palabras-clave:
Desertificación; MTLFC; Indicadores naturales; Uso del suelo
INTRODUÇÃO
O Bioma Caatinga tem ocorrência exclusiva no Brasil, sendo sua área principal localizada na Região Nordeste, ocorrendo também em um pequeno trecho da Região Sudeste (norte do Estado de Minas Gerais), onde domina o clima semiárido (menos de 800mm de precipitação/ano), totalizando 734 mil km2 (SILVA et al., 2004SILVA, J. M. C.; TABARELLI, M.; FONSECA, M. T.; LINS, L. V. (org). Biodiversidade da Caatinga: áreas e ações prioritárias para a conservação. Brasília (DF): MMA/UFPE/Conservation International – Biodiversitas – Embrapa Semi-árido, 2004. 382p.), o que equivale a cerca de 10% do território nacional. Oficialmente, é classificada como Savana Estépica (VELOSO et al., 1991VELOSO, H. P.; RANGEL-FILHO, A. L. R.; LIMA, J. C. A. Classificação da vegetação brasileira, adaptada a um sistema universal. Rio de Janeiro (RJ): IBGE, 1991. 124p.), embora mais recentemente, em nível internacional, esse Bioma venha sendo considerado como parte das Florestas Tropicais Sazonalmente Secas - STDF (OLIVEIRA FILHO et al., 2006OLIVEIRA FILHO, A. T.; JARENKOV, J. A.; RODAL, M. J. N. Floristic relationships of seasonally dry forests of eastern South America based on tree species distribution pattern. In: PENNINGTON, R. T.; LEWIS, G. P.; RATTER, J. A. Neotropical savannas and seasonally dry forests. Boca Raton (EUA): CRC Press, p. 159-190, 2006.; PENNINGTON et al., 2000PENNINGTON, R. T.; PRADO, D. A.; PENDRY, C. Neotropical seasonally dry forests and Pleistocene vegetation changes. J. Biogeogr., v. 27, p. 261-273, 2000.; PRADO, 2000PRADO, D. E. Seasonally dry forests of tropical South America: from forgotten ecosystems to a new phytogeographic unit. Edinburgo (UK): J. Bot., v. 57, p. 437-461, 2000.).
Quanto às alterações provocadas pelo desmatamento, a Caatinga é o terceiro Bioma mais degradado do Brasil, perdendo apenas para a Floresta Atlântica e o Cerrado (MYERS et al, 2000MYERS, N; MITTERMEIER, R. A; MITTERMEIER, C. G; FONSECA, G. A. B; KENT, J. Biodiversity hotspots for conservation priorities. Nature, n 403, p.853-859, 2000.). Estima-se que 80% da vegetação encontre-se completamente modificada, devido ao extrativismo e a agropecuária, apresentando-se a maioria dessas áreas em estádios iniciais ou intermediários de sucessão ecológica (ARAÚJO FILHO, 1996ARAÚJO FILHO, J. A. Desenvolvimento sustentável da caatinga. Sobral (CE): Ministério da Agricultura/EMBRAPA/CNPC, 1996. 45p.).
Como consequência das profundas alterações pelas quais vêm passando, a Caatinga apresenta grandes extensões onde a desertificação já se encontra instalada, existindo uma relação estreita entre este tipo de degradação, a vegetação e os solos, sendo o seu desenvolvimento iniciado com as modificações que venham a diminuir a presença da cobertura vegetal por períodos prolongados, aumentando os processos erosivos e deteriorando as propriedades físicas, químicas, biológicas e econômicas do solo CCD (1995)CCD. Convenção das Nações Unidas de Combate à Desertificação. Tradução: Delegação de Portugal. Lisboa (PT): Instituto de Promoção Ambiental, 1995. 55p..
Para o Brasil, de acordo com os dados oficiais, a área susceptível a ocorrência da desertificação abrange 1.338.076km2 e 1.482 municípios, sendo habitada por mais de 30 milhões de pessoas (BRASIL, 2004BRASIL. Programa de Ação Nacional de Combate à Desertificação e Mitigação dos Efeitos da Seca/PAN-Brasil. Brasília (DF): Ministério do Meio Ambiente/Secretaria de Recursos Hídricos, 2004. 214p.).
No caso do Estado da Paraíba, conforme BRASIL (2004)BRASIL. Programa de Ação Nacional de Combate à Desertificação e Mitigação dos Efeitos da Seca/PAN-Brasil. Brasília (DF): Ministério do Meio Ambiente/Secretaria de Recursos Hídricos, 2004. 214p., dos 223 municípios existentes, 208 são considerados susceptíveis a esse tipo de degradação (46.004km2), destacando-se a região dos Cariris Velhos (também conhecida como Cariri Paraibano ou Cariri) por apresentar elevados níveis de desertificação, em virtude da rarefação ou ausência completa de cobertura vegetal (SOUZA, et al., 2011SOUZA, B. I.; SUERTEGARAY, D. M. A.; LIMA, E. R. V. Evolução da desertificação no Cariri paraibano a partir da análise das modificações na vegetação. Rio Claro (SP): Geografia, v. 36, n. 1, p. 193-205, jan./abr. 2011.).
Paralelo aos problemas gerados pela desertificação, a Paraíba é considerada como um dos Estados que melhor representariam a flora da Caatinga, uma vez que grande parte do seu território é dominado pelo clima semiárido e abrange principalmente terrenos do complexo cristalino (ARAÚJO et al., 2005ARAÚJO, F. S.; RODAL, M. J. N.; BARBOSA, M. R. V.; MARTINS, F. R. Repartição da flora lenhosa no domínio da Caatinga. In: ARAÚJO, F. S.; RODAL, M. J. N.; BARBOSA, M. R. V. (orgs.). Análise das variações da biodiversidade do Bioma Caatinga. Suporte a estratégias regionais de conservação. Brasília (DF): Ministério do meio Ambiente, 2005. p. 15-33.), características físicas principais que condicionam o aparecimento desse tipo de vegetação, ainda que os levantamentos até agora efetuados sejam insuficientes para revelar a totalidade da riqueza existente.
Para realizar o levantamento da vegetação nas áreas de Caatinga, grande parte dos trabalhos desenvolvidos tem como base o uso das técnicas de fitossociologia, as quais vêm fornecendo informações importantes. Muitas dessas técnicas foram e continuar a ser utilizadas também nos estudos sobre desertificação, pois até certo ponto são capazes de gerar dados valiosos na análise desse tipo de degradação.
Entretanto, nos estudos que envolvem vegetação no Brasil, conforme Andrade et al. (2011)ANDRADE, L. A.; FABRICANTE, J. R.; ARAÚJO, E. L. Estudos de fitossociologia em vegetação de caatinga. In: FELFILI, J. M.; EISENLOHR, P. V.; NELO, M. M. R. F.; ANDRADE, L. A.; MEIRA NETO, J. A. A. (orgs.). Fitossociologia no Brasil. Métodos e estudos de caso. Viçosa (MG): Ed. UFV, 2011. p. 339-371., uma série de problemas em relação ao uso das técnicas de fitossociologia dificultam a obtenção de mais informações necessárias ao avanço das análises, deixando ainda muitas lacunas pendentes. Dentre as dificuldades mencionadas por esses autores, destacam-se a carência de levantamentos detalhados de solos e a falta ou deficiência de dados climáticos.
Sobre o comentário anteriormente exposto, destacamos que, mesmo que essas informações estivessem melhor disponibilizadas, pelo fato dos softwares que realizam esses tratamentos estatísticos não contemplarem uma forma de relacionar a vegetação aos elementos abióticos e também às intervenções antrópicas, as respostas obtidas ainda continuariam reducionistas.
No que diz respeito as discussões e contribuições da Geografia brasileira sobre essas temáticas, como regra elas vem sendo tratadas dentro da Geografia Física, principalmente enquanto parte da Questão Ambiental, para a qual diversos pesquisadores tem oferecido importantes informações. Ainda assim, se os avanços teóricos são expressivos, existe muito a ser feito na parte das técnicas utilizadas, já que em geral como essas foram desenvolvidas por outras ciências, ao serem adotadas na Geografia, a qual desde a sua origem buscou a relação entre Natureza e Sociedade, é comum que não se consiga apresentar satisfatoriamente as respostas que os profissionais dessa área procuram ou pelo menos apontar caminhos capazes de satisfazer parte das relações discutidas na teoria. Desse modo, é cada vez mais fundamental que os geógrafos apresentem sua contribuição também nessa parte do desenvolvimento científico.
Esse artigo foi desenvolvido com o objetivo de apresentar a aplicação de um método que preenche grande parte das lacunas nos estudos sobre levantamento e dinâmica da vegetação, ao mesmo tempo em que fornece informações fundamentais para os estudos de desertificação, inclusive apontando para as razões sociais que vêm interferindo na área estudada e gerando a degradação em questão.
O Método de Transecto Linear para Fanerófitos e Caméfitos - MTLFC, foi desenvolvido por Cámara e Díaz del Olmo (2004)CÁMARA, R. e DÍAZ DEL OLMO, F. Directrices y Gestión para la Conservación y Desarrollo Integral de un Humedal Centroamericano: Golfo de Montijo (Litoral del Pacífico, Panamá). Panamá: Embajada de España en Panamá, 2004. 311p., professores da Universidad de Sevilla (España)/Facultad de Geografía e Historia/Departamento de Geografia Física y Análisis Geográfico Regional. Para a sua criação, esses pesquisadores apoiaram-se em observações efetuadas por Gentry (1982GENTRY, A. H. Patterns of Neotropical plant species diversity. Evolutionary Biology 15:1-84, 1982. e 1996GENTRY, A. H. Diversity and floristic composition of Neotropical dry forest. In: BULLOCK, S., MEDINA, E. H. A.; MOONEY, H. A (eds.). Tropical deciduous forest ecosystems. Cambridge (UK): Cambridge University Press, 1996. p. 153-188.), incorporando indicadores geomorfológicos, climatológicos, hidrológicos e biogeográficos.
O MTLFC foi testado pela primeira vez no Brasil no Bioma Caatinga, principalmente na região dos Cariris Velhos, através do trabalho de Porto de Lima (2012)PORTO DE LIMA, V. R. Caracterización biogeográfica del bioma Caatinga en el sector semiárido de la cuenca del Río Paraíba – Noreste de Brasil: Propuesta de Ordenación y Gestión de um medio semiárido tropical. 2012. 355f. Tesis (Doctorado en Geografía Física y Análisis Geográfico Regional). Sevilla (ESP): Universidad de Sevilla, 2012., obtendo resultados inéditos, embora o seu uso voltado á análise da degradação em questão esteja sendo realizado pela primeira vez.
PROCEDIMENTOS METODOLÓGICOS
Na aplicação do MTLFC são utilizados transectos que estão relacionados com os perfis de solos, atributos texturais e profundidade das raízes, com a caracterização das formações superficiais e a capacidade de campo associada, bem como os processos hídricos das águas subsuperficiais, estabelecendo peculiaridades que em conjunto resultam nas sinúsias de cada formação vegetal.
Este trabalho foi realizado nas seguintes etapas:
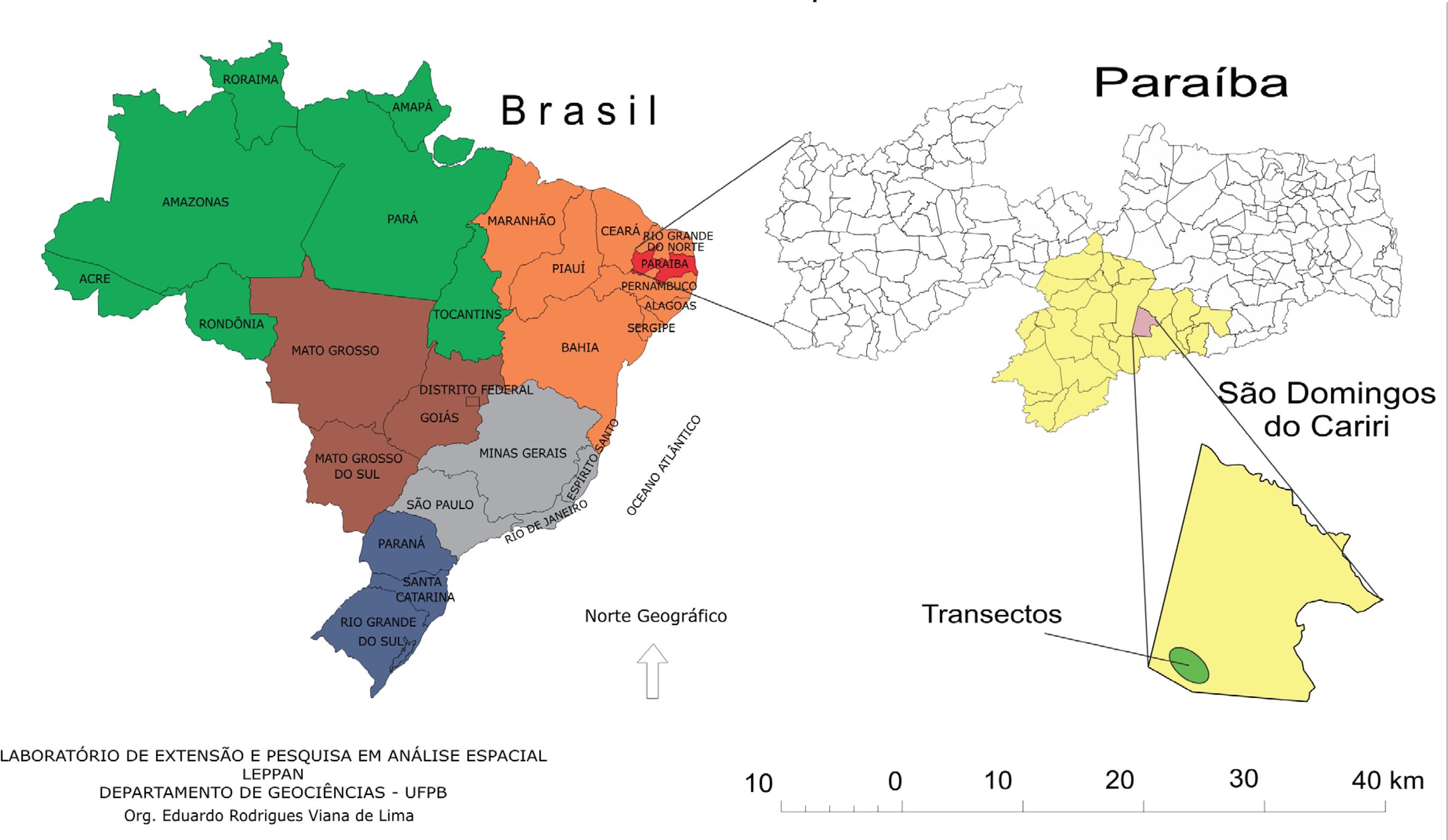
1. Seleção da área para coletas de dados: Realizada no município de São Domingos do Cariri, localidade de Salgadinho (figura 1). A escolha dessa área foi fundamentada no critério fisionômico da paisagem, situada em um pediplano, com topografia suavemente ondulada, vegetação aberta com pouca variação de espécies, sinais visíveis de corte e queimada e solos do tipo Luvissolo Crômico órtico vértico (EMBRAPA, 2006EMBRAPA. Sistema brasileiro de classificação de solos. Rio de Janeiro (RJ): Embrapa/Centro nacional de Pesquisa de Solos, 2006. 306p.) com erosão superficial aparente, resultando em um conjunto representativo de grande parte da região dos Cariris Velhos;
2. Levantamento dos dados em campo: Iniciado a partir da definição da unidade básica de amostragem em um censo de plantas lenhosas arbustivas e arbóreas em uma área de 0,1ha. tratada como uma parcela, onde foram definidos 10 transectos lineares, cada um compreendendo uma área de 50m x 2m, delimitados por uma fita métrica. Os indivíduos recenseados foram aqueles situados dentro da distância de 1m de qualquer lado da fita métrica. Foi considerada também neste inventário a posição do indivíduo identificado e medido, tanto na distância longitudinal como a sua separação à direita ou esquerda da fita métrica. Para estabelecer a estrutura vertical e horizontal das espécies, se o indivíduo possuísse Diâmetro na Altura do Peito - DAP igual ou inferior a 2cm, foi medida a sua altura, o diâmetro maior e menor. Se possuísse DAP igual ou superior a 2cm, foi medida a altura, o raio maior e menor da copa;
3. Tratamento analítico dos dados em planilhas desenvolvidas no software Excel: Teve a finalidade de apresentar os atributos das unidades amostradas e seus elementos em relação às características estruturais da formação vegetal;
4. Identificação do Índice de Valor de Importância – IVI e dos Índices de Diversidade, com base no software Past: Foram calculados os índices Alfa de Simpson e Berger-Parker, para identificar a dominância, e Shannon, para identificar a equidade, uma vez que se trata de uma área degradada, portanto de habitat homogêneo. Além disso, estes são índices amplamente utilizados para análise de diversidade (MORENO, 2001MORENO, C. E. Métodos para medir la Biodiversidad. Zaragoza (ESP.): SEA, Manuales & Tesis, v. 1, 2001. 84p.), inclusive em se tratando de estudos sobre Caatinga.
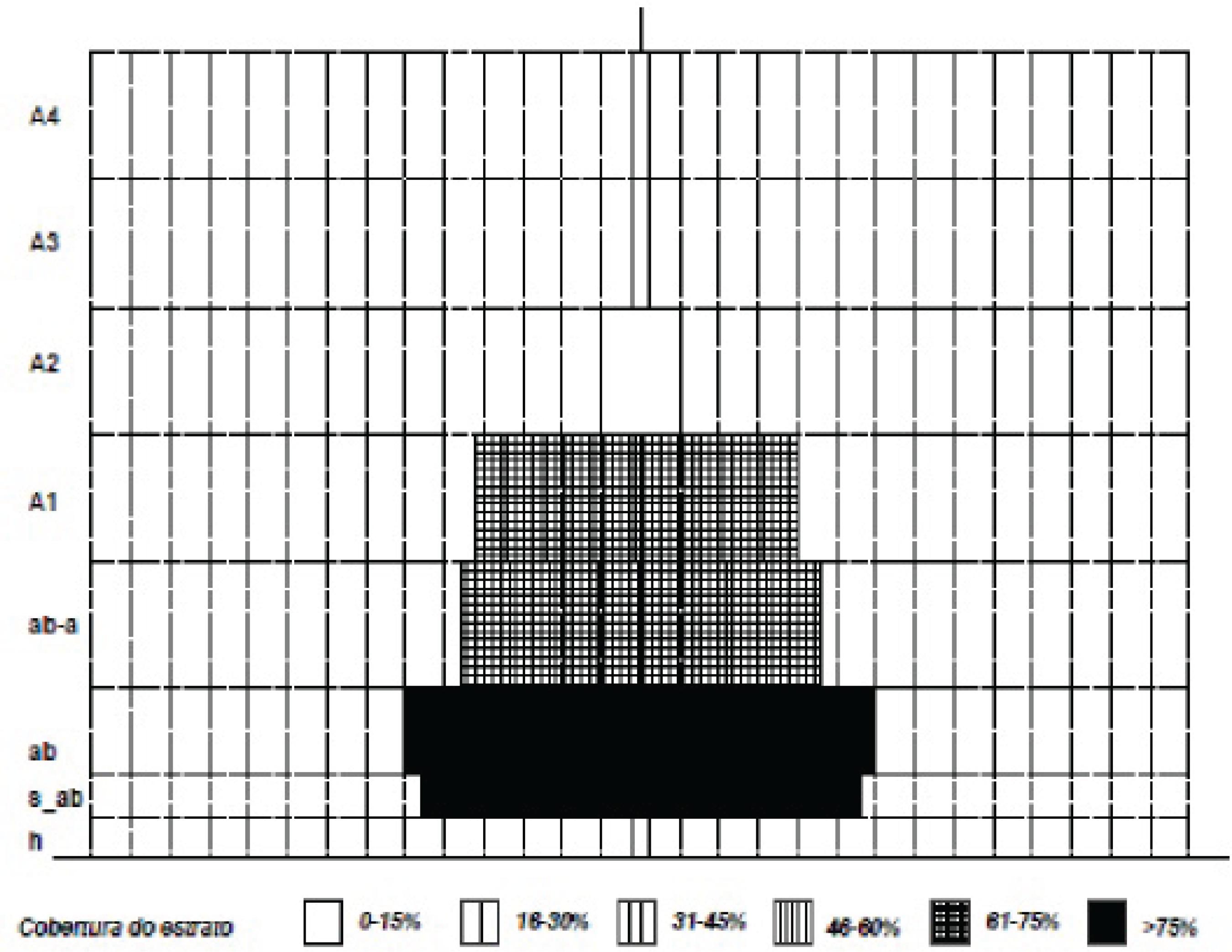
5. Identificação da estrutura vertical da formação, desenvolvida a partir do software OpenOffice.org.Draw: Objetivou estabelecer a dinâmica das formações vegetais amostradas, a qual fica expressa no Diagrama Ecodinámico de Riqueza – Estrutura – Cobertura (DEREC);
6. Identificação da estrutura horizontal da formação, através do Diagrama de Bolhas, desenvolvido no software Excel;
7. Aplicação do Método de Regimes Ecodinámicos - MEREC: Fundamenta-se na análise de dados paramétricos expressos pelo Balanço Hídrico, cuja base de cálculo foi desenvolvida originalmente por Thorntwaite e Matter (1955THORNTHWAITE, C. W. e MATHER, J. R. The Water Balance. Drexel (GERM): Institute of Technology, Laboratory of Climatology. Climatology, 8: 1-104, 1955. e 1957)THORNTHWAITE, C. W. e MATHER, J. R. Instructions and Tables for Computing Potential Evapotranspiration and the Water Balance. Drexel Institute of Technology, Laboratory of Climatology. Climatology, 10:181-311, 1957., e Balanço Bioclimático, com base em Monteiro de Burgos e Rebollar (1974)MONTEIRO DE BURGOS, J. L. e REBOLLAR, J. L. G. Diagramas bioclimáticos. Madrid (ESP): Ministerio de Agricultura/ICONA, 1974. 328p., disponibilizados em um aplicativo para o cálculo automático desenvolvido por Martínez Battle (2002)MARTÍNEZ BATLLE, J. R. Sabanas de la República Dominicana: análisis ecodinámico de patrones tipológicos y sus ecotonos. 2002. 354f. Tesis (Doctorado en Geografía Física y Análisis Geográfico Regional). Sevilla (ESP): Universidad de Sevilla, 2002. e Cámara (2004)CÁMARA, R. e DÍAZ DEL OLMO, F. Directrices y Gestión para la Conservación y Desarrollo Integral de un Humedal Centroamericano: Golfo de Montijo (Litoral del Pacífico, Panamá). Panamá: Embajada de España en Panamá, 2004. 311p. (http://www.geografiafisica.org/2012/09/17/plantilla-para-el-calculo-del-balance-hidrico-y-el-diagrama-bioclimatico/);
8. Análise de solos: Foram observados em campo padrões de estrutura, textura e porosidade, através de análise de perfil em trincheiras cavadas na parcela. Ainda que não prevista no método original, em função da preocupação principal desse trabalho estar baseada na análise da desertificação, foram coletadas 2 amostras de solos com o objetivo de identificar os níveis químicos e de fertilidade do material extraído. Para a realização desse tipo de coleta foi utilizado um trado, sendo o material retirado a 10cm de profundidade em 2 pontos distintos, acondicionado em sacos plásticos e depois enviado para análise no Laboratório de Fertilidade de Solos da Universidade Federal da Paraíba/Campus II.
RESULTADOS E DISCUSSÃO
Espécies, Famílias e Indivíduos
Em toda a parcela foram identificadas 13 espécies, distribuídas em 6 Famílias, contabilizando um total de 489 indivíduos, conforme o quadro 1, a seguir.
C. sonderianus, C. echioides, A. pyrifolium, P. pyramidalis e J. molíssima sobressaíram nesse levantamento em relação as demais espécies, com 423 indivíduos, o que corresponde a 86,5% do total identificado. Dessas espécies, todas pioneiras, C. sonderianus, A. pyrifolium e P. pyramidalis se destacam quanto ao número de indivíduos na maioria dos trabalhos de levantamento fitossociológico desenvolvidos em Caatinga (SAMPAIO, 1996SAMPAIO, E. V. S. B. Fitossociologia. In: SAMPAIO, E. V. S. B.; MAYO, S. J.; BARBOSA, M. R. V. (Eds.). Pesquisa botânica nordestina: progresso e perspectivas. Recife (PE): SBB, p. 203-230, 1996.).
Quanto às Famílias, a maior diversidade encontrada neste levantamento esteve presente em Fabácea, Euphorbiácea e Cactácea, com três espécies cada uma. Essas Famílias também ocupam papel de destaque em outras formações xerófilas na América do Sul (CABRERA e WILLINK, 1973CABRERA, A. L. e WILLINK, A. Biogeografia de América Latina. 2ª ed. Washington (EUA): OEA, 1973. 325p.; SARMIENTO, 1975SARMIENTO, G. The dry plant formations of South America and their forest connections. Journal of Biogeography, v. 2, p. 233-251, 1975.).
Fabácea é considerada a Família mais diversa na Caatinga, possivelmente pela longa história de diversificação em vegetações de clima seco e a estabilidade florística dessas áreas, associadas às adaptações morfológicas que ocorreram em suas espécies quando da adaptação à escassez hídrica, altas temperaturas e baixa umidade (CARDOSO e QUEIROZ, 2010CARDOSO, D. B. O. S. e QUEIROZ, L. P. Caatinga no contexto de uma metacomunidade: evidências da biogeografia, padrões filogenéticos e abundância de espécies em Leguminosas. In: CARVALHO, C. J. B e ALMEIDA, E. A. B. (orgs.). Biogeografia da América do Sul. Padrões e processos. São Paulo (SP): Roca, 2010. p. 241-260.).
Sobre Euphorbiácea, além da sua elevada resistência a falta de água, importante papel de dispersão é desempenhado pela zoocoria, especialmente as formigas, pois em sua alimentação, ao retirarem a polpa das sementes, ocorre diminuição do ataque de fungos, aumentando as taxas de germinação no local onde os propágulos estão depositados. No caso de conduzirem a semente ao formigueiro para a extração do material, temos a diminuição do processo de competição intraespecífica das plântulas e também a predação das sementes embaixo da planta-mãe. Logo que a parte utilizada é subtraída, as sementes são depositadas nas lixeiras, ambientes mais ricos em matéria orgânica que os solos que estão no entorno da colônia, beneficiando-se também da maior permeabilidade, aeração, profundidade e granulometria dessas áreas, intensificando-se assim o sucesso reprodutivo e consequentemente a estrutura espacial das populações vegetais (LEAL, 2005LEAL, I. R. Dispersão de sementes por formigas na Caatinga. In: LEAL, I. R.; TABARELLI, M.; SILVA, J. M. C. (eds.). Ecologia e conservação da Caatinga. Recife (PE): Ed. Universitária da UFPE, 2005. p. 593-624.).
Particularmente sobre C. sonderianus, o papel de outro animal também é muito importante para explicar a sua elevada presença nessas áreas. Trata-se de Zenaida auriculata, espécie de pássaro que costuma se concentrar ás centenas nos locais onde existem essa planta em busca das suas sementes para alimento, o que ocorre logo após sucederem algumas chuvas que proporcionem o seu aparecimento (AGUIRRE, 1976AGUIRRE, A. Nidificação da Zenaida auriculata (des Murs). Brasília (BR): Revista Brasil Florestal 3 (6): 14-18, 1972.; ANTAS, 1986ANTAS, P. T. Z. A nidificação da avoante, Zenaida auriculata, no Nordeste do Brasil, relacionada com o substrato fornecido pela vegetação. Curitiba (PR): Revista Brasileira de Zoologia, v. 3, n. 7, 1986.).
Em área degradada próxima onde ocorreram as coletas do presente trabalho e com características parecidas, Barbosa et al. (2007)BARBOSA, M. R. V.; LIMA, I. B.; LIMA, J. R. CUNHA, J. P.; AGRA, M. F.; THOMAS, W. W. Vegetação e flora no Cariri Paraibano. Rio de Janeiro (RJ): Oecol. Bras., 11 (3): 313-322, 2007.identificaram somente 12 espécies e 6 Famílias, sendo que P. Pyramidalis ocupou a primeira posição em todos os parâmetros fitossociológicos analisados, seguida de C. sonderianus e Combretum leprosum, resultados semelhantes ao do presente trabalho, excetuando a última espécie.
Já em diversas áreas preservadas nessa mesma região, Barbosa et al. (2007)BARBOSA, M. R. V.; LIMA, I. B.; LIMA, J. R. CUNHA, J. P.; AGRA, M. F.; THOMAS, W. W. Vegetação e flora no Cariri Paraibano. Rio de Janeiro (RJ): Oecol. Bras., 11 (3): 313-322, 2007. identificaram um total de 396 espécies, distribuídas em 90 Famílias, dominando Fabácea com 71 espécies. Esses dados demonstram a elevada riqueza existente nos remanescentes de Caatinga melhor preservados do Cariri, em contraste com os que estão degradados.
Ressalta-se ainda que nessas poucas áreas onde existem situações de baixa alteração da Caatinga, a diversidade encontrada é superior a muitos outros remanescentes preservados de STDF existentes no mundo, a exemplo do México, país que como o Brasil é reconhecido em nível internacional como um dos de maior megadiversidade. Nesse caso, em condições de altitude, precipitação e preservação parecidas com as que foram identificadas por Barbosa et al. (2007)BARBOSA, M. R. V.; LIMA, I. B.; LIMA, J. R. CUNHA, J. P.; AGRA, M. F.; THOMAS, W. W. Vegetação e flora no Cariri Paraibano. Rio de Janeiro (RJ): Oecol. Bras., 11 (3): 313-322, 2007. no Cariri, Trejo (2005)TREJO. I. Análisis de la diversidad de la Selva Baja Caducifolia en México. In: HALFFTER, G.; SOBERÓN, J.; KOLEFF, P.; MELIC, A. (eds.). Sobre Diversidad Biológica: el significado de las Diversidades alfa, beta y gamma. Zaragoza (ESP.): SEA/CONABIO/Grupo Diversitas-México/CONACYT, 2005. p. 111-122. só conseguiu identificar na Selva Baixa Caducifólia um número máximo de 115 espécies, incluindo-se aí as lianas.
IVI e Índices de Diversidade
Analisadas sob a perspectiva do IVI, tanto em relação a cobertura como ao DAP, P. pyramidalis, A. pyrifolium, C. sonderianus, C. echioides e J. mollissima se confirmam como as mais importantes na parcela (quadro 2), ao passo que os baixos valores encontrados para as espécies restantes denotam a predominância de indivíduos de pequeno porte e em pequena quantidade.
Os índices de diversidade Alfa apresentaram os seguintes resultados: Simpson - 0.8334; Berger-Parker - 0.2556 e Shannon - 2.002. Com base nos dados obtidos, confirma-se que entre as espécies existentes na parcela amostrada existe uma diversidade baixa (13, no total, conforme visto anteriormente), dominância de algumas poucas espécies e baixa abundância.
Dos índices obtidos, a título de comparação com os resultados de outros trabalhos na Caatinga, o de diversidade de Shannon é o mais utilizado. Entre os levantamentos realizados no mesmo município, em áreas próximas aos efetuados nesse trabalho, com características físicas e de uso do solo parecidas, utilizando o mesmo método, Porto de Lima (2012)PORTO DE LIMA, V. R. Caracterización biogeográfica del bioma Caatinga en el sector semiárido de la cuenca del Río Paraíba – Noreste de Brasil: Propuesta de Ordenación y Gestión de um medio semiárido tropical. 2012. 355f. Tesis (Doctorado en Geografía Física y Análisis Geográfico Regional). Sevilla (ESP): Universidad de Sevilla, 2012. identificou valores de 1,52 e 1,97, portanto muito semelhantes a presente investigação.
Comparando esses levantamentos com aqueles desenvolvidos em condições físicas e de uso do solo parecidas em área próxima, ainda que utilizando outra metodologia, destaca-se o de Andrade et al. (2005)ANDRADE, L. A.; PEREIRA, I. M.; LEITE, U. T.; BARBOSA, M. R. V. Análise da cobertura de duas fitofisionomias de caatinga, com diferentes históricos de uso, no município de São João do Cariri, Estado da Paraíba. Lavras (MG): Cerne, v. 11, n. 3, p. 253-262, jul/set 2005., os quais identificaram valores de 1,43 e 1,51, demonstrando que nessa região existem condições ainda piores que as encontradas no presente levantamento e nos de Porto de Lima (2012)PORTO DE LIMA, V. R. Caracterización biogeográfica del bioma Caatinga en el sector semiárido de la cuenca del Río Paraíba – Noreste de Brasil: Propuesta de Ordenación y Gestión de um medio semiárido tropical. 2012. 355f. Tesis (Doctorado en Geografía Física y Análisis Geográfico Regional). Sevilla (ESP): Universidad de Sevilla, 2012..
Existem outros trabalhos relacionados aos índices de diversidade desse Bioma, entretanto, como não foram apresentadas de forma mais detalhada aspectos relativos às características pluviométricas, pedológicas e de uso dos solos encontradas nas áreas onde ocorreram os levantamentos, tendo em vista a importância dessas variantes no estabelecimento das fitofisionomias da Caatinga, qualquer tentativa de comparação desses números com os que foram expostos neste trabalho levaria a conclusões precipitadas.
ESTRATOS OCUPADOS PELAS ESPÉCIES, COBERTURA E INDIVÍDUOS
Questões como processos de sucessão ecológica e dinâmica podem encontrar explicações através da análise da estrutura vertical e horizontal de um bosque (FINOL, 1971FINOL, U. H. Nuevos parâmetros a considerarse en el análise estrutural de las selvas virgenes tropicales. Caracas (VEN.): Rev. For., v. 14, n. 21, p 24-42, 1971.). Neste sentido, em relação a estrutura vertical em estratos e cobertura da parcela, obteve-se como resultado o gráfico DEREC, expresso na figura 2, o qual mostra a distribuição das espécies identificadas entre os estratos arbustivo (ab: 0,6 – 1,5m), com 12 representantes (92% do total); subarbustivo (s_ab: 0,3 – 0,6m), com 11 representantes (85% do total); arbustivo alto (ab_a: 1,5 – 3,5m), com 9 representantes (69% do total); arbóreo baixo (A1: 3,5 – 5m), com 8 representantes (61% do total), e arbóreo intermediário (A2: 5 – 10m), com 2 representantes (15% do total).
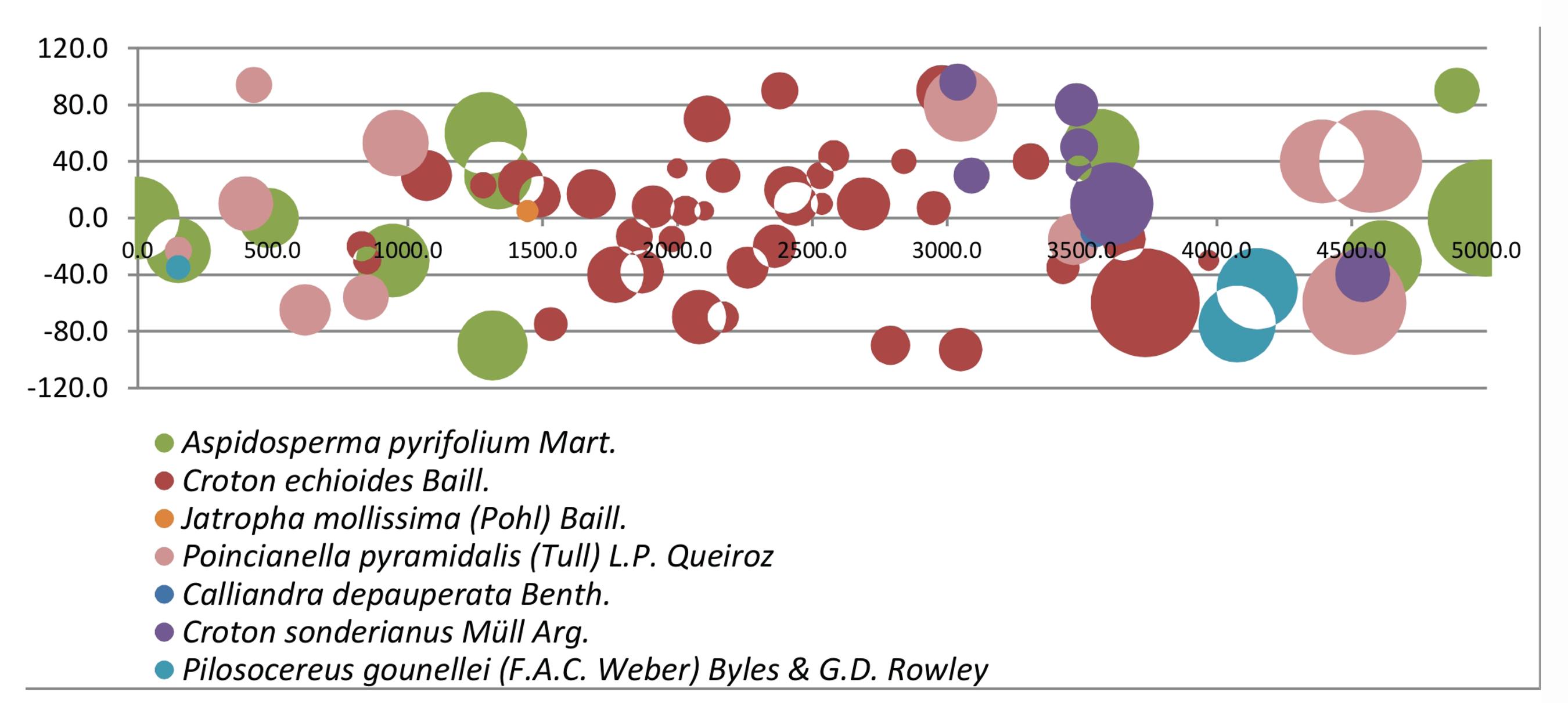
A estrutura horizontal da vegetação, por sua vez, apresenta a distribuição e a superfície coberta pelas espécies e indivíduos em cada transecto, também levando em consideração a área basal e a dominância relativa. Reflete assim os efeitos da história geológica, do clima, das associações bióticas e das intervenções efetuadas pelos homens no manejo do solo, tal como preconizado por Dansereau (1968)DANSEREAU, P. Les structures de végétation. Lisboa (PORT.): Finisterre, v. 3: 147-174, 1968.. A título de exemplo, segue na figura 3, abaixo, um dos Diagramas de Bolhas resultantes da aplicação do MTLFC em um dos 10 transectos feitos nesse trabalho.
Na figura 3 constam 7 das 13 espécies identificadas em toda a parcela, ao mesmo tempo que apresenta como os indivíduos das mesmas se distribuem ao longo dos 50 metros lineares do transecto, 1 metro à esquerda e 1 metro á direita da fita. Refletindo o que foi observado de forma geral na parcela, percebemos nesse exemplo uma estrutura horizontal onde os indivíduos estão organizados de forma esparsada e pouco adensada, tal como pode ser visualizado na figura 4, onde temos uma paisagem mais parecida á de um campo aberto que uma floresta, sendo esta última a fitofisionomia característica da Caatinga em áreas de vegetação preservada.
Diagrama de Bolhas, representando a organização da cobertura horizontal da vegetação em um transecto na parcela.
Em relação a percentagem de cobertura, destacaram-se os estratos arbóreo baixo (53,21% da cobertura) e arbustivo alto (47,67% da cobertura), seguidos dos estratos arbustivo (10,66% da cobertura), subarbustivo (1,43% da cobertura) e arbóreo intermediário (1,26% da cobertura) seguir.
O domínio dos estratos arbóreo baixo e arbustivo alto em relação a cobertura revelam a elevada importância das copas das espécies A. pyrifolium e P. pyramidalis na parcela, as quais também se destacaram por apresentar os únicos indivíduos com altura superior a 5 metros, conforme pode ser observado mais adiante, no quadro 3.
Quanto ao estrato ocupado pelos indivíduos, o maior número destes ocorreu no arbustivo alto (209 - 42,7% do total) e arbustivo (138 - 28,2% do total). As espécies com maior número de indivíduos que ocorreram nesses estratos foram C. sonderianus (125), C. echioides (96), A. pyrifolium (78), P. pyramidalis (78) e J. molíssima (46), como pode ser visualizado no quadro 3, a seguir.
O elevado número de indivíduos de C. sonderianus no estrato arbustivo alto indica, em princípio, que a área estaria em estágio inicial de sucessão ecológica, idéia que poderia ser ratificada pela grande presença de A. pyrifolium no estrato arbustivo e P. pyramidalis no estrato arbustivo alto, sinalizando nesses dois últimos casos forte presença de plantas jovens (em idade adulta as duas últimas espécies ocupam, respectivamente, os estratos arbóreo baixo e arbóreo intermediário). Entretanto, a presença de marcas de corte na maioria dos indivíduos dessas espécies, além de queimadas na parcela, revelam que se trata de uma situação de rebrota, a qual possivelmente vem se repetindo a vários anos.
De acordo com Travassos (2012)TRAVASSOS, I. S. “Florestas brancas” do semiárido nordestino: desmatamento e desertificação no Cariri paraibano. 2012. 155f. Dissertação (Mestrado em Geografia) – UFPB, João Pessoa, 2012., entre 1970 até a primeira metade de 1990 essa parte do Cariri foi uma grande produtora de lenha, cujo maior consumidor era a área polarizada pela cidade de Campina Grande, onde existem muitas indústrias. Após esse período, em função da diminuição da vegetação, a produção caiu substancialmente, o que poderia favorecer a recuperação de uma série de espécies. Entretanto, levando em consideração os impactos ambientais gerados e o fato da população dessa região utilizar tradicionalmente a madeira para vários outros usos, a pressão sobre a vegetação não cessou com a queda da extração de lenha destinada ao Setor Secundário.
Quanto às queimadas, essa prática no manejo do solo é tradicional nessa região, sendo ainda hoje a base de grande parte do aproveitamento agropecuário existente, servindo também para produzir carvão utilizado como fonte energética. Apesar de antiga, o auge dessa atividade ocorreu entre o final do século XIX e a década de 1950, motivada pela expansão da cotonicultura. Nesse período, em função do mercado favorável ao consumo da fibra, esse cultivo ocupou tanto solos menos frágeis a essa prática como outros não apropriados a um uso mais intenso, resultando em áreas que até hoje tem dificuldade em se recuperar (SOUZA et al., 2010SOUZA, B. I.; SUERTEGARAY, D. M. A.; LIMA, E. R. V. Políticas Públicas, uso do solo e desertificação nos Cariris Velhos (PB/Brasil). Barcelona (ESP.): Scripta Nova, v. XIV, n. 311, enero de 2010. Disponível em: www.ub.edu/geocrit/sn/sn-311.htm.
www.ub.edu/geocrit/sn/sn-311.htm...
), a exemplo de onde foram realizados os levantamentos para esse trabalho.
Sobre o efeito dessas ações nas características das plantas da Caatinga, a maioria das espécies desse Bioma rebrota após o corte (SAMPAIO et al., 1998SAMPAIO, E. V. S. B.; ARAÚJO, E. L.; SALCEDO, I. H.; TIESSEN, H. Regeneração da vegetação de caatinga após corte e queima, em Serra Talhada, PE. Brasília (DF): Pesquisa Agropecuária Brasileira, 33: 621-632, 1998.), apresentando, em alguns casos já analisados, taxas de sobrevivência maiores que 50%, independente do tipo de corte aplicado (FIGUEIRÔA et al., 2006FIGUEIRÔA, J. M.; PAREYN, F. G. C.; ARAÚJO, E. L.; SILVA, C. E.; SANTOS, V. F.; CUTLER, D. F.; BARACAT, A.; GASSON, P. Effects of cutting regimes in the dry and wet season on survival and sprouting of woody species from the semi-arid caatinga of northeast Brazil. Forest Ecology and Management, 229, p: 294-303, 2006.). Entretanto, se este é seguido de queima, a rebrota diminui de forma progressiva, na medida em que ocorre o aumento na intensidade de combustão, além dos efeitos persistirem por mais de seis anos na biomassa em relação a densidade e área basal.
Também o uso da madeira de C. sonderianus, A. pyrifolium e P. pyramidalis na construção de cercas é outro fator importante quanto a interferência em sua estratificação. Nesse caso, como as cercas normalmente são renovadas em um período máximo de 8 anos (SOUZA, 2008SOUZA, B. I. Cariri Paraibano: do silêncio do lugar à desertificação. 2008. 198f. Tese (Doutorado em Geografia). Porto Alegre (RS): UFRGS, 2008.), o uso constante dessas espécies impede que as mesmas possam se desenvolver até o estrato máximo.
Além do que já foi relatado, dificultando ainda mais a evolução sucessória em áreas como essa, papel crucial é exercido pela pecuária caprina, atividade também tradicional no Cariri Paraibano e que a partir da década de 1970 vem destacando essa região em todo o Brasil, principalmente devido a grande produção leiteira, onde o Estado exerce um papel fundamental como incentivador e financiador (SOUZA et al., 2010SOUZA, B. I.; SUERTEGARAY, D. M. A.; LIMA, E. R. V. Políticas Públicas, uso do solo e desertificação nos Cariris Velhos (PB/Brasil). Barcelona (ESP.): Scripta Nova, v. XIV, n. 311, enero de 2010. Disponível em: www.ub.edu/geocrit/sn/sn-311.htm.
www.ub.edu/geocrit/sn/sn-311.htm...
).
Os caprinos são pouco seletivos em relação ás plantas que utilizam como alimento, fazendo uso de até 70% das espécies existentes na Caatinga, o que é intensificado por permanecerem junto a vegetação nativa consumindo-a durante o ano inteiro (ARAÚJO FILHO et al., 1996ARAÚJO FILHO, J. A. Desenvolvimento sustentável da caatinga. Sobral (CE): Ministério da Agricultura/EMBRAPA/CNPC, 1996. 45p., 1999aARAÚJO FILHO, J. A.; BARBOSA, T. M. L.; CARVALHO, F. C. Sistema de produção silvopastoril para o semi-árido nordestino. Sobral (CE): Boletim da Embrapa, 29: 1-2, 1999a., 1999bARAÚJO FILHO, J. A.; BARBOSA, T. M. L; CARVALHO, F. C.; CAVALCANTI, A. C. R. Sistema de produção agrossilvopastoril para o semi-árido nordestino. Sobral (CE): Boletim da Embrapa, 30: 1-2, 1999b.), só recebendo alimentação complementar nos casos de estiagens muito prolongadas.
Esses animais constituem importante fator de seleção natural, afetando principalmente arbustos e árvores perenifólias, como Ziziphus joazeiro e Spondias tuberosa, ou espécies decíduas com ciclo reprodutivo parcial ou completo na estação seca, como Myracroduon urundeuva e Commiphora leptophoeos, todas elas clímax na Caatinga (LEAL et al., 2005LEAL, I. R.; VICENTE, A.; TABARELLI, M. Herbivoria por caprinos na caatinga da região de Xingó: uma análise preliminar. In: LEAL, I. R. & SILVA, J. M. C. (Edit.). Ecologia e conservação da caatinga. Recife (PE): Ed. Universitária/UFPE, 2005. p. 695-715.). Além disso, ao darem preferência às folhas e brotos jovens, acabam influenciando negativamente no crescimento das espécies utilizadas, provocando muitas vezes o nanismo destas, enquanto ao consumirem frutos nativos, muitas sementes são totalmente trituradas, impedindo assim a reprodução das plantas.
Dessa maneira, a pecuária caprina, particularmente quando praticada com altas taxas de lotação em áreas de Caatinga e associadas a outras formas de pressão sobre a vegetação, têm o poder de empobrecer e reduzir o porte das plantas, levando à ocorrência de desertificação, o que também já foi constatado em outras regiões semiáridas fora do Brasil (LEAL et al., 2005LEAL, I. R.; VICENTE, A.; TABARELLI, M. Herbivoria por caprinos na caatinga da região de Xingó: uma análise preliminar. In: LEAL, I. R. & SILVA, J. M. C. (Edit.). Ecologia e conservação da caatinga. Recife (PE): Ed. Universitária/UFPE, 2005. p. 695-715.).
Do ponto de vista temporal, análises feitas por Souza et al. (2011)SOUZA, B. I.; SUERTEGARAY, D. M. A.; LIMA, E. R. V. Evolução da desertificação no Cariri paraibano a partir da análise das modificações na vegetação. Rio Claro (SP): Geografia, v. 36, n. 1, p. 193-205, jan./abr. 2011. utilizando imagens de satélite, indicam que a situação encontrada nessa parte do Cariri só tem mudado em relação a intensidade e expansão das condições de degradação. Nesse caso, individualizando o município de São Domingos do Cariri do restante da região dos Cariris Velhos e destacando a parcela onde foram realizados os levantamentos desse trabalho (figura 5), a análise de imagens de satélite permite inferir que no ano de 1989 essa área apresentava desertificação moderada, enquanto em 2005 passou a ser muito grave.
Município de São Domingos do Cariri e área das coletas de dados (transectos) em imagens de satélite de 1989 e 2005.
Tal característica representa uma regra para todo o município em questão, onde em 1989 existiam 164,9km2 de área desertificada, o que correspondia a 68,9% do seu território, enquanto no ano 2005 esse tipo de degradação aumentou, atingindo 187,9km2, o que perfazia 78,5% do seu território (SOUZA et al., 2011SOUZA, B. I.; SUERTEGARAY, D. M. A.; LIMA, E. R. V. Evolução da desertificação no Cariri paraibano a partir da análise das modificações na vegetação. Rio Claro (SP): Geografia, v. 36, n. 1, p. 193-205, jan./abr. 2011.).
BALANÇOS HÍDRICO E BIOCLIMÁTICO
O Balanço Hídrico expressa as condições climáticas e pedológicas dominantes de uma área. Nesse caso, onde está localizada a parcela, há uma precipitação média de chuvas de 385mm/ano, temperaturas médias elevadas (23,8ºC), pequena amplitude térmica (3,4ºC) e déficit hídrico de 1215,79mm/ano. Em relação aos solos, estes apresentam pequena profundidade (90cm), textura franco-argilosa e déficit de água de 830,79mm/ano. Tratados em conjunto, esses dados podem ser parcialmente visualizados na figura 6, a seguir.
Como corre deficiência hídrica tanto do ponto de vista climático como pedológico ao longo de todo ano, outros mecanismos de obtenção de água pela vegetação, não captados pelo cálculo do Balanço Hídrico, devem ser investigados para que se possa entender o desenvolvimento das espécies da Caatinga nessa região.
Parte dos mecanismos de desenvolvimento das plantas pode ser compreendido através do Balanço Bioclimático, o qual vai expressar a atividade vegetativa através das condições dominantes de umidade e temperatura. Neste trabalho identificou-se Intensidade Bioclimática Seca cálida - IBSc de agosto a janeiro, Intensidade Bioclimática Condicionada cálida - IBCc nos meses de fevereiro e março e Intensidade Bioclimática Livre cálida - IBLc de abril a julho, conforme pode ser observado na figura 7.
Com base nos resultados apresentados, de agosto a janeiro (6 meses) temos a estação seca e as espécies não teriam atividade vegetativa; em fevereiro e março (2 meses), com a ocorrência de algumas chuvas, a atividade vegetativa ficaria limitada às espécies que apresentam necessidade mínimas de água; de abril à julho (4 meses), com o estabelecimento da estação chuvosa, a atividade vegetativa não apresentaria limites, podendo ser integralmente aproveitada pelas plantas.
Conforme já foi visto anteriormente, o déficit hídrico está presente nessa parte do Cariri ao longo de todo ano, tanto do ponto de vista climático como pedológico. Logo, a atividade vegetativa, mesmo na estação chuvosa, está condicionada a outros fatores ainda pouco conhecidos para essa parte do Brasil, os quais dizem respeito ás relações planta-planta e planta-solo, condicionando a existência de microclimas.
Essas relações funcionam como atenuadores de situações de estresse vivenciadas pelas plantas, tornado as condições mais favoráveis ao recrutamento e estabelecimento dos indivíduos em áreas de Caatinga, a exemplo da diminuição da insolação excessiva abaixo da copa de arbustos e árvores, o que faz com que, nessa situação, as taxas de evaporação do solo sejam relativamente baixas (SILANS e SILVA, 2007SILANS, A. M. B. P. e SILVA, F. M. Fluxo de calor sensível e evapotranspiração na Caatinga: estudo experimental. Porto Alegre (RS): Revista Brasileira de Recursos Hídricos, v. 12, n. 1, p. 177-188, jan./mar. 2007.).
Além do que já foi relatado anteriormente, é provável que para a atividade vegetativa das espécies dominantes nessa região a participação da precipitação oculta seja decisiva. Nesse caso, embora ainda não comprovada para o Bioma Caatinga, Agan e Berliner (2006)AGAN, N. e BERLINER, P. R. Dew formation and water vapor adsorption in semi-arid environments – A review. Journal of Arid Environments, v. 65, p. 572-590, 2005. têm sugerido que em regiões semiáridas ocorre reumidificação da camada superficial do solo através do orvalho ou da absorção do vapor de água do ar.
Conforme os dados da Estação Climatológica de São João do Cariri, as maiores amplitudes térmicas anuais para os Cariris Velhos ocorrem entre os meses de abril à julho, período chuvoso, sendo superiores a 5ºC, o que favorece a intensificação da formação de orvalho. Ao mesmo tempo, também nesse período ocorrem os maiores percentuais de umidade relativa do ar, ultrapassando o valor de 80%. Nesse caso, a observação dessas informações com o que já se conhece sobre o comportamento fenológico de algumas espécies da Caatinga, indicam que a idéia de Agan e Berliner (2006)AGAN, N. e BERLINER, P. R. Dew formation and water vapor adsorption in semi-arid environments – A review. Journal of Arid Environments, v. 65, p. 572-590, 2005. estaria correta.
Nesse Bioma, conforme Machado et al. (1997)MACHADO, I. C. S.; BARROS, L. M.; SAMPAIO, E. V. S. B. Phenology of the caatinga species at Serra Talhada, PE, Northeasthern Brazil. Biotropica, v. 29, p. 57-68, 1997., a fenologia da floração da maioria das espécies lenhosas é sincronizada com o início da estação chuvosa, padrão também observado em outras áreas de STDF neotropicais (BULLOCK, 1995BULLOCK, S. H. Plant reproduction in neotropical dry forests. In: BULLOCK, S.; MOONEY, H.; MEDINA, E. (eds.). Seasonnally Dry Tropical Forests. Cambridge (UK): Cambridge University Press, 1995. p. 277-297.). Além disso, a sazonalidade climática exerce influência sobre o número de sementes no solo, sendo esse efeito mais forte na profundidade de 0-5cm (SANTOS et al., 2010SAMPAIO, E. V. S. B.; ARAÚJO, E. L.; SALCEDO, I. H.; TIESSEN, H. Regeneração da vegetação de caatinga após corte e queima, em Serra Talhada, PE. Brasília (DF): Pesquisa Agropecuária Brasileira, 33: 621-632, 1998.) as quais apresentam, para a maioria das espécies, da mesma forma que na floração, germinação no início das chuvas (COSTA e ARAÚJO, 2003COSTA, R. C. e ARAÚJO, F. S. Densidade, germinação e flora do banco de sementes no solo, no final da estação seca, em uma área de caatinga, Quixadá, CE. São Paulo (SP): Acta Botânica Brasílica, v. 17, n. 2, p. 1-6, Abr./Jun. 2003.).
Sobre C. sonderianus, P. pyramidalis, S. galheirensis e A. pyrifolium, espécies de elevado grau de xerofilia e que mais se destacaram no levantamento realizado para esse trabalho, dados levantados por Parente (2009)PARENTE, H. N. Avaliação da vegetação e do solo em áreas de caatinga sob pastejo caprino no Cariri da Paraíba. Tese (Doutorado Integrado em Zootecnia). Areia (PB): UFPB, 2009. no município de São João do Cariri revelam que o seu período de rebrotamento tem início logo após os primeiros eventos de chuvas, o que confirma os resultados observados em relação ao ciclo vegetativo obtido através do Balanço Bioclimático para os meses de fevereiro e março. Quanto à floração e frutificação, estas ocorrem em meados da estação úmida, enquanto a queda de folhas acontece logo após o término das chuvas.
ANÁLISE DOS SOLOS
Sobre os solos, as observações em campo, juntamente com a consulta a bibliografia (EMBRAPA, 2006EMBRAPA. Sistema brasileiro de classificação de solos. Rio de Janeiro (RJ): Embrapa/Centro nacional de Pesquisa de Solos, 2006. 306p.), permitiram identifica-los como Luvissolo Crômico, apresentando revestimento pedregoso na superfície (pavimento desértico), crosta superficial de 5 a 10mm de espessura, profundidade rasa (cerca de 90cm), com horizonte A de 15cm, seguido de um horizonte B textural com 25cm e horizonte C com 50cm.
Acrescentamos que esse tipo de solo também apresenta argilas de alta atividade, caracterizando-se como moderadamente ácido a neutro e com elevado potencial nutricional quando não degradado, em virtude das altas quantidades de nutrientes disponíveis às plantas e de minerais primários facilmente intemperizáveis, especialmente o potássio (EMBRAPA, 2006EMBRAPA. Sistema brasileiro de classificação de solos. Rio de Janeiro (RJ): Embrapa/Centro nacional de Pesquisa de Solos, 2006. 306p.).
Os resultados das análises laboratoriais na parcela encontram-se apresentados no quadro 4, a seguir. Para efeito de comparação e discussão sobre as respostas dos elementos analisados nesse trabalho, destacamos no mesmo quadro os resultados de outras análises efetuadas por Souza (2008)SOUZA, B. I. Cariri Paraibano: do silêncio do lugar à desertificação. 2008. 198f. Tese (Doutorado em Geografia). Porto Alegre (RS): UFRGS, 2008., em áreas com vegetação preservada situadas em lugares próximos de onde foram realizadas essas coletas.
Com base nos resultados expostos, observa-se uma grande semelhança entre os números encontrados para os elementos químicos analisados nas amostras da área degradada, a exceção do fósforo (P), o qual se mostrou mais presente na primeira situação apresentada. Sabe-se que em situações em que existe forte proximidade com a rocha os valores desse elemento tendem a aumentar substancialmente. Nesse caso, como esse tipo de solo apresenta pouca profundidade, a situação apresentada é comum, o que explica a diferença encontrada. Destaca-se que o fósforo (P) é um elemento fundamental para o desenvolvimento e produção da vegetação (JORGE, 1972aJORGE, J. A. Fósforo. In: MONIZ, A. C (Coord.). Elementos de Pedologia. São Paulo (SP): Polígono/Ed. da Universidade de São Paulo, 1972a. p. 191-197.), sendo em geral deficiente em áreas de clima semiárido (SAMPAIO et al., 2005SAMPAIO, E. V. S. B.; ARAÚJO, M. S. B.; SAMPAIO, Y. S. B. Impactos ambientais da agricultura no processo de desertificação no Nordeste do Brasil. Revista de Geografia do Deptº de Ciências Geográficas, v. 22, nº 01. Recife (PE): UFPE, p. 93-113, 2005.; SILVEIRA et al., 2006SILVEIRA, M. M. L.; ARAÚJO, M. S. B.; SAMPAIO, E. V. S. B. Distribuição de fósforo em diferentes ordens de solo do semi-árido da Paraíba e de Pernambuco. Viçosa (MG): Revista Brasileira de Ciência do Solo, v. 30, n. 2, p. 1-13, 2006.), salvo na ocorrência da situação descrita anteriormente.
A comparação dos dados presentes no quadro 4 mostra situações que em geral são bastante distintas para as áreas degradadas e preservadas. Neste sentido, destaca-se particularmente os resultados dos elementos que mais influenciam a fertilidade: potássio (K), atuante no crescimento, conformação e qualidade dos frutos (MESSIAS et al., 2008MESSIAS, A. S.; SILVA, D. J.; FREIRE, F. J.; SILVA, M. C. L. Fertilizantes. In: CAVALCANTI, F. J. A. (coord.). Recomendações de adubação para o Estado de Pernambuco: 2ª aproximação. Recife (PE): IPA, 2008. p. 89-103.) e matéria orgânica (M.O), a qual melhora as propriedades físicas e químicas do solo, servindo de fonte de elementos minerais para as plantas (JORGE, 1972bJORGE, J. A. Matéria Orgânica. In: MONIZ, A. C. (Coord.). Elementos de Pedologia. São Paulo (SP): Polígono/Ed. da Universidade de São Paulo, 1972b. p. 169-178.), além de estabilizar a estrutura do solo, influenciando diretamente na resistência à erosão (GALINDO, 2007GALINDO, I. C. L. Relação solo-vegetação em áreas sob processo de desertificação no Estado de Pernambuco. 2007. 255f. Tese (Doutorado em Ciência do Solo). Recife (PE): UFRPE, 2007.). A estes dois é acrescentado o fósforo (P), sobre o qual já foram tecidos comentários.
Com base nos resultados apresentados no quadro 4, tanto o potássio (K) como a matéria orgânica (M.O) ocorreram em maiores quantidades na área preservada, ao passo que os níveis de fósforo (P) são fortemente influenciados pela maior ou menor distância da rocha, fazendo com que o seu valor na primeira situação de área degradada se encontre próximo ou até superior às existentes na área preservada.
Os resultados apresentados demonstram que existe uma forte relação entre a vegetação, os processos erosivos e os diferentes níveis de fertilidade presentes. Nesse caso, na área degradada, a rarefação da vegetação favorece a ação das chuvas torrenciais típicas da região e consequentemente a lixiviação e a intensificação da destruição dos agregados do solo. Além disso, como a Caatinga está adaptada a um padrão mais elevado de fertilidade dos solos, é possível que uma série de espécies encontre dificuldade para voltar a colonizar novamente esses ambientes, enquanto não forem restabelecidas melhores condições pedológicas.
Como o processo anteriormente destacado é dinâmico, temos também uma influência direta do desmatamento na modificação da estrutura original dos solos afetados, os quais apresentarão aumento de densidade e redução da porosidade, compactação e formação de crostas nas camadas superficiais, aumento da resistência à penetração e a consequente diminuição da infiltração da água, o que leva a intensificação dos efeitos da seca, aspectos já detectados em outras áreas do semiárido brasileiro (NUNES et al., 2006NUNES, L. A. P. L.; ARAÚJO FILHO, J. A.; MENEZES, R. I. Q. Impacto da queimada e do pousio sobre a qualidade de um solo sob caatinga no semiárido nordestino. Mossoró (RN): Revista Caatinga, v. 19, n. 2, p. 200-208, abril/junho 2006.; SOUZA e MARTINS, 2012SOUZA, B. I. e MARTINS, V. C. Infiltração da água em solos de zona semiárida e sua relação com os processos de desertificação. Porto Alegre (RS): Boletim Gaúcho de Geografia, v. 39, p. 25-39, 2012.) e também na África (ALBERGEL et al., 1992ALBERGEL, J.; CASENAVE, A.; RIBSTEIN, P.; VALENTIN, C. Aridité climatic, aridité édaphique. Étude des conditions de l’infiltrabilité em Afrique Tropical Séche. In: LE FLOC’H, E.; GROUZIS, M.; CORNET, A.; BILLE, J. C (eds.). L’aridité: une contrainte au développement. Caractérisation, réponses biologiques, strategies des societés. Paris (FR): ORSTOM, 1992. p. 123-130.).
Trazendo essa discussão á luz dos processos que podem desencadear a desertificação, esses são fortemente influenciados pelas condições pedológicas ligadas a capacidade de retenção de água no perfil dos solos (VERHEYE, 1990VERHEYE, W. H. Une approche agropédologique de la desertification. Paris (FR): Secheresse, n. 2, vol. 1, juillet, p. 94-97, 1990.). Neste caso, a degradação progressiva da cobertura vegetal é um fator decisivo nessa questão, a qual acaba interferindo diretamente na sucessão ecológica dos ambientes afetados.
Com base nas considerações efetuadas, temos a diminuição da possibilidade de estabelecimento e crescimento de algumas plântulas, facilitando o domínio de espécies que apresentam requerimentos hídricos menores (SHACHAK et al., 1998SHACHAK, M.; SACHS, M.; MOSHE, I. Ecosystem management of desertfield shrublands in Israel. Ecosystems 1: 475-483, 1998.; VIDIELLA e ARMESTO, 1989MARTÍNEZ BATLLE, J. R. Sabanas de la República Dominicana: análisis ecodinámico de patrones tipológicos y sus ecotonos. 2002. 354f. Tesis (Doctorado en Geografía Física y Análisis Geográfico Regional). Sevilla (ESP): Universidad de Sevilla, 2002.), tal como foi observado no que diz respeito ás 5 espécies mais importantes do levantamento realizado nesse trabalho, dentre as quais Silva et al. (2004)SILVA, J. M. C.; TABARELLI, M.; FONSECA, M. T.; LINS, L. V. (org). Biodiversidade da Caatinga: áreas e ações prioritárias para a conservação. Brasília (DF): MMA/UFPE/Conservation International – Biodiversitas – Embrapa Semi-árido, 2004. 382p.destacam J. mollissima, P. pyramidalis e C. echioides como as de maior resistência às secas na Caatinga.
Ressalta-se, porém, que para serem obtidos mais dados relacionados a influência das alterações da infiltração da água em solos do semiárido e a sua relação com a recolonização vegetal nos ambientes atingidos por desmatamento excessivo, é necessário um número maior de estudos que façam uma interface entre os dois elementos destacados e destes com o comportamento do banco de sementes presente nessas áreas, uma das principais estratégias de sobrevivência a longo prazo das comunidades vegetais nas regiões de climas secos no mundo (BASKIN e BASKIN, 1998BASKIN, C. C. e BASKIN, J. M. Seeds, ecology, biogeography and evolution of dormancy and germination. New York (USA): Academic Press, 1998. 342p.; KEMP, 1989KEMP. P. R. Seed Banks and vegetation process in deserts. In: LECK, M. A.; PARKER, V. T.; SIMPSON, R. L. (eds.). Ecology of soil seed Banks. New York (USA): Academic Press, 1989. p. 257-280.), aspecto ainda pouco conhecido para o Bioma Caatinga (COSTA e ARAÚJO, 2003COSTA, R. C. e ARAÚJO, F. S. Densidade, germinação e flora do banco de sementes no solo, no final da estação seca, em uma área de caatinga, Quixadá, CE. São Paulo (SP): Acta Botânica Brasílica, v. 17, n. 2, p. 1-6, Abr./Jun. 2003.).
CONSIDERAÇÕES FINAIS
Os resultados apresentados demonstraram a eficiência do MTLFC nos estudos sobre levantamento de vegetação e na identificação das razões do estabelecimento das espécies nas áreas analisadas, apresentando neste último caso uma integração com diversos elementos abióticos que, tratados em conjunto e associados a outros dados, tornam esse método um instrumento poderoso para uso em pesquisas que exijam uma análise interdisciplinar, a exemplo da desertificação.
De forma mais específica, na área analisada esse trabalho demonstrou que:
1. Em toda a parcela ocorreram 13 espécies divididas em 6 Famílias, totalizando 489 indivíduos, dentre os quais destacaram-se C. sonderianus (125), C. echioides (96), A. pyrifolium (78), P. pyramidalis (78) e J. molíssima (46), representando 86,5% de todo o levantamento e os maiores IVIs encontrados;
2. Fabácea, Euphorbiácea e Cactácea destacaram-se com o maior número de espécies (3 para cada Família), enquanto Euphorbiácea foi dominante quanto ao número de indivíduos (267, o que equivale a 54,6% do total levantado);
3. Os índices de diversidade Alfa encontrados demonstraram que existe baixa diversidade, dominância de algumas poucas espécies e baixa abundância: Simpson – 0.8334, Berger-Parker – 0.2556 e Shannon – 2.002;
4. Os estratos arbustivo e subarbustivo sobressaíram em relação a distribuição do número de espécies (12 e 11, respectivamente). Os estratos arbóreo baixo e arbustivo apresentaram-se dominantes em relação a percentagem na cobertura (53,21% e 47,67%), em função das copas de A. pyrifolium e P. pyramidalis. Os estratos arbustivo alto e arbustivo destacaram-se quanto ao número de indivíduos (209 – 42,7% do total e 138 – 28,2% do total);
5. Foi detectada elevada presença de marcas de corte e queimadas na vegetação da parcela, além do uso pelos caprinos, o que indica interferência direta na diversidade e estrutura encontradas;
6. O Balanço Hídrico mostrou-se deficitário ao longo de todo o ano, enquanto o Balanço Bioclimático demonstrou que existem 6 meses sem atividade vegetativa, 2 meses com atividade vegetativa limitada às espécies com baixas necessidades hídricas e 4 meses sem restrição a atividade vegetativa;
7. Existem indicativos de que a atividade vegetativa seria condicionada pela presença do orvalho e do vapor d’água e que sob a presença de biomassa os teores de umidade na camada superficial dos solos são mais elevados;
8. Os solos apresentam crosta superficial nas áreas sem vegetação o que, juntamente com a diferença textural entre os horizontes A e Bt, pode estar influenciando diretamente na infiltração da água, tornando a carência hídrica mais acentuada e fazendo com que as poucas espécies vegetais existentes na área sejam aquelas que apresentam o máximo de adaptação a essa condição;
9. A fertilidade dos solos encontra-se muito alterada, apresentando principalmente baixos níveis de potássio e matéria orgânica, o que também pode influenciar negativamente em relação ao estabelecimento de espécies mais exigentes quanto a esses elementos.
Em função dos resultados encontrados, áreas como essa precisam de uma intervenção direta para serem recuperadas, processo que deve começar pela diminuição da pressão sobre a vegetação ainda presente, seguido de um reflorestamento para o qual as espécies vegetais pioneiras identificadas mostram elevada aptidão para essa fase inicial, em decorrência da sua grande resistência a esse tipo de ambiente. Ao mesmo tempo, formas de uso sustentáveis do solo devem ser incentivadas nos ambientes de clima seco do Brasil, evitando a criação e a expansão de áreas com processos de desertificação. Neste sentido, a adoção dos sistemas agrosilvopastoris deve ter os seus efeitos melhor investigados.
AGRADECIMENTOS
Á Coordenação de Aperfeiçoamento de Pessoal de Nível Superior – CAPES, pela concessão de bolsa de estudos ao primeiro autor deste artigo para realização de Pós-Doutorado na Universidade de Sevilla/Espanha (Processo nº 5338-12-3).
REFERÊNCIA BIBLIOGRÁFICA
- AGAN, N. e BERLINER, P. R. Dew formation and water vapor adsorption in semi-arid environments – A review. Journal of Arid Environments, v. 65, p. 572-590, 2005.
- AGUIRRE, A. Nidificação da Zenaida auriculata (des Murs). Brasília (BR): Revista Brasil Florestal 3 (6): 14-18, 1972.
- ALBERGEL, J.; CASENAVE, A.; RIBSTEIN, P.; VALENTIN, C. Aridité climatic, aridité édaphique. Étude des conditions de l’infiltrabilité em Afrique Tropical Séche. In: LE FLOC’H, E.; GROUZIS, M.; CORNET, A.; BILLE, J. C (eds.). L’aridité: une contrainte au développement. Caractérisation, réponses biologiques, strategies des societés. Paris (FR): ORSTOM, 1992. p. 123-130.
- ANDRADE, L. A.; PEREIRA, I. M.; LEITE, U. T.; BARBOSA, M. R. V. Análise da cobertura de duas fitofisionomias de caatinga, com diferentes históricos de uso, no município de São João do Cariri, Estado da Paraíba. Lavras (MG): Cerne, v. 11, n. 3, p. 253-262, jul/set 2005.
- ANDRADE, L. A.; FABRICANTE, J. R.; ARAÚJO, E. L. Estudos de fitossociologia em vegetação de caatinga. In: FELFILI, J. M.; EISENLOHR, P. V.; NELO, M. M. R. F.; ANDRADE, L. A.; MEIRA NETO, J. A. A. (orgs.). Fitossociologia no Brasil Métodos e estudos de caso. Viçosa (MG): Ed. UFV, 2011. p. 339-371.
- ANTAS, P. T. Z. A nidificação da avoante, Zenaida auriculata, no Nordeste do Brasil, relacionada com o substrato fornecido pela vegetação. Curitiba (PR): Revista Brasileira de Zoologia, v. 3, n. 7, 1986.
- ARAÚJO, F. S.; RODAL, M. J. N.; BARBOSA, M. R. V.; MARTINS, F. R. Repartição da flora lenhosa no domínio da Caatinga. In: ARAÚJO, F. S.; RODAL, M. J. N.; BARBOSA, M. R. V. (orgs.). Análise das variações da biodiversidade do Bioma Caatinga Suporte a estratégias regionais de conservação. Brasília (DF): Ministério do meio Ambiente, 2005. p. 15-33.
- ARAÚJO FILHO, J. A. Desenvolvimento sustentável da caatinga. Sobral (CE): Ministério da Agricultura/EMBRAPA/CNPC, 1996. 45p.
- ARAÚJO FILHO, J. A.; GADELHA, E. R.; LEITE, P. Z.; SOUZA, S. M. A.; CRISPIM, M. C. R. Composição botânica e química da dieta de ovinos e caprinos em pastoreio combinado na região dos Inhamuns, Ceará. Revista da Sociedade Brasileira de Zootecnia Viçosa (MG): UFV, 25: 383-395, 1996.
- ARAÚJO FILHO, J. A.; BARBOSA, T. M. L.; CARVALHO, F. C. Sistema de produção silvopastoril para o semi-árido nordestino. Sobral (CE): Boletim da Embrapa, 29: 1-2, 1999a.
- ARAÚJO FILHO, J. A.; BARBOSA, T. M. L; CARVALHO, F. C.; CAVALCANTI, A. C. R. Sistema de produção agrossilvopastoril para o semi-árido nordestino. Sobral (CE): Boletim da Embrapa, 30: 1-2, 1999b.
- BARBOSA, M. R. V.; LIMA, I. B.; LIMA, J. R. CUNHA, J. P.; AGRA, M. F.; THOMAS, W. W. Vegetação e flora no Cariri Paraibano. Rio de Janeiro (RJ): Oecol. Bras., 11 (3): 313-322, 2007.
- BASKIN, C. C. e BASKIN, J. M. Seeds, ecology, biogeography and evolution of dormancy and germination. New York (USA): Academic Press, 1998. 342p.
- BRASIL. Programa de Ação Nacional de Combate à Desertificação e Mitigação dos Efeitos da Seca/PAN-Brasil Brasília (DF): Ministério do Meio Ambiente/Secretaria de Recursos Hídricos, 2004. 214p.
- BULLOCK, S. H. Plant reproduction in neotropical dry forests. In: BULLOCK, S.; MOONEY, H.; MEDINA, E. (eds.). Seasonnally Dry Tropical Forests Cambridge (UK): Cambridge University Press, 1995. p. 277-297.
- CABRERA, A. L. e WILLINK, A. Biogeografia de América Latina 2ª ed. Washington (EUA): OEA, 1973. 325p.
- CAMARA, R. Escalonamiento Bioclimático, Regímenes Ecodinámicos y Formaciones Vegetales de la Isla de la Española en República Dominicana Estudios em Biogeografía. Girona (ESP): Terrassa/Servei de Publications de la Universitat de Girona, p. 39-58, 2004.
- CÁMARA, R. e DÍAZ DEL OLMO, F. Directrices y Gestión para la Conservación y Desarrollo Integral de un Humedal Centroamericano: Golfo de Montijo (Litoral del Pacífico, Panamá). Panamá: Embajada de España en Panamá, 2004. 311p.
- CARDOSO, D. B. O. S. e QUEIROZ, L. P. Caatinga no contexto de uma metacomunidade: evidências da biogeografia, padrões filogenéticos e abundância de espécies em Leguminosas. In: CARVALHO, C. J. B e ALMEIDA, E. A. B. (orgs.). Biogeografia da América do Sul Padrões e processos. São Paulo (SP): Roca, 2010. p. 241-260.
- CCD. Convenção das Nações Unidas de Combate à Desertificação Tradução: Delegação de Portugal. Lisboa (PT): Instituto de Promoção Ambiental, 1995. 55p.
- COSTA, R. C. e ARAÚJO, F. S. Densidade, germinação e flora do banco de sementes no solo, no final da estação seca, em uma área de caatinga, Quixadá, CE. São Paulo (SP): Acta Botânica Brasílica, v. 17, n. 2, p. 1-6, Abr./Jun. 2003.
- DANSEREAU, P. Les structures de végétation. Lisboa (PORT.): Finisterre, v. 3: 147-174, 1968.
- EMBRAPA. Sistema brasileiro de classificação de solos Rio de Janeiro (RJ): Embrapa/Centro nacional de Pesquisa de Solos, 2006. 306p.
- FIGUEIRÔA, J. M.; PAREYN, F. G. C.; ARAÚJO, E. L.; SILVA, C. E.; SANTOS, V. F.; CUTLER, D. F.; BARACAT, A.; GASSON, P. Effects of cutting regimes in the dry and wet season on survival and sprouting of woody species from the semi-arid caatinga of northeast Brazil Forest Ecology and Management, 229, p: 294-303, 2006.
- FINOL, U. H. Nuevos parâmetros a considerarse en el análise estrutural de las selvas virgenes tropicales. Caracas (VEN.): Rev. For., v. 14, n. 21, p 24-42, 1971.
- GALINDO, I. C. L. Relação solo-vegetação em áreas sob processo de desertificação no Estado de Pernambuco 2007. 255f. Tese (Doutorado em Ciência do Solo). Recife (PE): UFRPE, 2007.
- GENTRY, A. H. Patterns of Neotropical plant species diversity. Evolutionary Biology 15:1-84, 1982.
- GENTRY, A. H. Diversity and floristic composition of Neotropical dry forest. In: BULLOCK, S., MEDINA, E. H. A.; MOONEY, H. A (eds.). Tropical deciduous forest ecosystems Cambridge (UK): Cambridge University Press, 1996. p. 153-188.
- JORGE, J. A. Fósforo. In: MONIZ, A. C (Coord.). Elementos de Pedologia São Paulo (SP): Polígono/Ed. da Universidade de São Paulo, 1972a. p. 191-197.
- JORGE, J. A. Matéria Orgânica. In: MONIZ, A. C. (Coord.). Elementos de Pedologia São Paulo (SP): Polígono/Ed. da Universidade de São Paulo, 1972b. p. 169-178.
- KEMP. P. R. Seed Banks and vegetation process in deserts. In: LECK, M. A.; PARKER, V. T.; SIMPSON, R. L. (eds.). Ecology of soil seed Banks New York (USA): Academic Press, 1989. p. 257-280.
- LEAL, I. R. Dispersão de sementes por formigas na Caatinga. In: LEAL, I. R.; TABARELLI, M.; SILVA, J. M. C. (eds.). Ecologia e conservação da Caatinga Recife (PE): Ed. Universitária da UFPE, 2005. p. 593-624.
- LEAL, I. R.; VICENTE, A.; TABARELLI, M. Herbivoria por caprinos na caatinga da região de Xingó: uma análise preliminar. In: LEAL, I. R. & SILVA, J. M. C. (Edit.). Ecologia e conservação da caatinga Recife (PE): Ed. Universitária/UFPE, 2005. p. 695-715.
- MACHADO, I. C. S.; BARROS, L. M.; SAMPAIO, E. V. S. B. Phenology of the caatinga species at Serra Talhada, PE, Northeasthern Brazil. Biotropica, v. 29, p. 57-68, 1997.
- MARTÍNEZ BATLLE, J. R. Sabanas de la República Dominicana: análisis ecodinámico de patrones tipológicos y sus ecotonos 2002. 354f. Tesis (Doctorado en Geografía Física y Análisis Geográfico Regional). Sevilla (ESP): Universidad de Sevilla, 2002.
- MESSIAS, A. S.; SILVA, D. J.; FREIRE, F. J.; SILVA, M. C. L. Fertilizantes. In: CAVALCANTI, F. J. A. (coord.). Recomendações de adubação para o Estado de Pernambuco: 2ª aproximação. Recife (PE): IPA, 2008. p. 89-103.
- MONTEIRO DE BURGOS, J. L. e REBOLLAR, J. L. G. Diagramas bioclimáticos Madrid (ESP): Ministerio de Agricultura/ICONA, 1974. 328p.
- MORENO, C. E. Métodos para medir la Biodiversidad Zaragoza (ESP.): SEA, Manuales & Tesis, v. 1, 2001. 84p.
- MYERS, N; MITTERMEIER, R. A; MITTERMEIER, C. G; FONSECA, G. A. B; KENT, J. Biodiversity hotspots for conservation priorities. Nature, n 403, p.853-859, 2000.
- NUNES, L. A. P. L.; ARAÚJO FILHO, J. A.; MENEZES, R. I. Q. Impacto da queimada e do pousio sobre a qualidade de um solo sob caatinga no semiárido nordestino. Mossoró (RN): Revista Caatinga, v. 19, n. 2, p. 200-208, abril/junho 2006.
- OLIVEIRA FILHO, A. T.; JARENKOV, J. A.; RODAL, M. J. N. Floristic relationships of seasonally dry forests of eastern South America based on tree species distribution pattern. In: PENNINGTON, R. T.; LEWIS, G. P.; RATTER, J. A. Neotropical savannas and seasonally dry forests Boca Raton (EUA): CRC Press, p. 159-190, 2006.
- PARENTE, H. N. Avaliação da vegetação e do solo em áreas de caatinga sob pastejo caprino no Cariri da Paraíba Tese (Doutorado Integrado em Zootecnia). Areia (PB): UFPB, 2009.
- PENNINGTON, R. T.; PRADO, D. A.; PENDRY, C. Neotropical seasonally dry forests and Pleistocene vegetation changes. J. Biogeogr, v. 27, p. 261-273, 2000.
- PORTO DE LIMA, V. R. Caracterización biogeográfica del bioma Caatinga en el sector semiárido de la cuenca del Río Paraíba – Noreste de Brasil: Propuesta de Ordenación y Gestión de um medio semiárido tropical. 2012. 355f. Tesis (Doctorado en Geografía Física y Análisis Geográfico Regional). Sevilla (ESP): Universidad de Sevilla, 2012.
- PRADO, D. E. Seasonally dry forests of tropical South America: from forgotten ecosystems to a new phytogeographic unit. Edinburgo (UK): J. Bot., v. 57, p. 437-461, 2000.
- SAMPAIO, E. V. S. B. Fitossociologia. In: SAMPAIO, E. V. S. B.; MAYO, S. J.; BARBOSA, M. R. V. (Eds.). Pesquisa botânica nordestina: progresso e perspectivas. Recife (PE): SBB, p. 203-230, 1996.
- SAMPAIO, E. V. S. B.; ARAÚJO, E. L.; SALCEDO, I. H.; TIESSEN, H. Regeneração da vegetação de caatinga após corte e queima, em Serra Talhada, PE. Brasília (DF): Pesquisa Agropecuária Brasileira, 33: 621-632, 1998.
- SAMPAIO, E. V. S. B.; ARAÚJO, M. S. B.; SAMPAIO, Y. S. B. Impactos ambientais da agricultura no processo de desertificação no Nordeste do Brasil. Revista de Geografia do Deptº de Ciências Geográficas, v. 22, nº 01. Recife (PE): UFPE, p. 93-113, 2005.
- SANTOS, D. M.; SILVA, K. A.; SANTOS, J. M. F. F.; LOPES, C. G. R.; PIMENTEL, R. M. M.; ARAÚJO, E. L. Variação espaço-temporal do banco de sementes em uma área de Floresta Tropical Seca (Caatinga) – Pernambuco. Recife (PE): Revista de Geografia, UFPE, v. 27, n. 1, Jan/Abr. 2010.
- SARMIENTO, G. The dry plant formations of South America and their forest connections. Journal of Biogeography, v. 2, p. 233-251, 1975.
- SHACHAK, M.; SACHS, M.; MOSHE, I. Ecosystem management of desertfield shrublands in Israel. Ecosystems 1: 475-483, 1998.
- SILANS, A. M. B. P. e SILVA, F. M. Fluxo de calor sensível e evapotranspiração na Caatinga: estudo experimental. Porto Alegre (RS): Revista Brasileira de Recursos Hídricos, v. 12, n. 1, p. 177-188, jan./mar. 2007.
- SILVA, J. M. C.; TABARELLI, M.; FONSECA, M. T.; LINS, L. V. (org). Biodiversidade da Caatinga: áreas e ações prioritárias para a conservação Brasília (DF): MMA/UFPE/Conservation International – Biodiversitas – Embrapa Semi-árido, 2004. 382p.
- SILVEIRA, M. M. L.; ARAÚJO, M. S. B.; SAMPAIO, E. V. S. B. Distribuição de fósforo em diferentes ordens de solo do semi-árido da Paraíba e de Pernambuco. Viçosa (MG): Revista Brasileira de Ciência do Solo, v. 30, n. 2, p. 1-13, 2006.
- SOUZA, B. I. Cariri Paraibano: do silêncio do lugar à desertificação. 2008. 198f. Tese (Doutorado em Geografia). Porto Alegre (RS): UFRGS, 2008.
- SOUZA, B. I. e MARTINS, V. C. Infiltração da água em solos de zona semiárida e sua relação com os processos de desertificação. Porto Alegre (RS): Boletim Gaúcho de Geografia, v. 39, p. 25-39, 2012.
- SOUZA, B. I.; SUERTEGARAY, D. M. A.; LIMA, E. R. V. Políticas Públicas, uso do solo e desertificação nos Cariris Velhos (PB/Brasil). Barcelona (ESP.): Scripta Nova, v. XIV, n. 311, enero de 2010. Disponível em: www.ub.edu/geocrit/sn/sn-311.htm.
» www.ub.edu/geocrit/sn/sn-311.htm - SOUZA, B. I.; SUERTEGARAY, D. M. A.; LIMA, E. R. V. Evolução da desertificação no Cariri paraibano a partir da análise das modificações na vegetação. Rio Claro (SP): Geografia, v. 36, n. 1, p. 193-205, jan./abr. 2011.
- THORNTHWAITE, C. W. e MATHER, J. R. The Water Balance. Drexel (GERM): Institute of Technology, Laboratory of Climatology. Climatology, 8: 1-104, 1955.
- THORNTHWAITE, C. W. e MATHER, J. R. Instructions and Tables for Computing Potential Evapotranspiration and the Water Balance. Drexel Institute of Technology, Laboratory of Climatology. Climatology, 10:181-311, 1957.
- TRAVASSOS, I. S. “Florestas brancas” do semiárido nordestino: desmatamento e desertificação no Cariri paraibano. 2012. 155f. Dissertação (Mestrado em Geografia) – UFPB, João Pessoa, 2012.
- TREJO. I. Análisis de la diversidad de la Selva Baja Caducifolia en México. In: HALFFTER, G.; SOBERÓN, J.; KOLEFF, P.; MELIC, A. (eds.). Sobre Diversidad Biológica: el significado de las Diversidades alfa, beta y gamma. Zaragoza (ESP.): SEA/CONABIO/Grupo Diversitas-México/CONACYT, 2005. p. 111-122.
- VELOSO, H. P.; RANGEL-FILHO, A. L. R.; LIMA, J. C. A. Classificação da vegetação brasileira, adaptada a um sistema universal Rio de Janeiro (RJ): IBGE, 1991. 124p.
- VERHEYE, W. H. Une approche agropédologique de la desertification. Paris (FR): Secheresse, n. 2, vol. 1, juillet, p. 94-97, 1990.
- VIDIELLA, P. E. e ARMESTO, J. J. Emergence of ephemeral plant species from soil samples of the chilean coastal desert in reponse to experimental irrigation. Revista Chilena de Historia Natural, 66: 99-107, 1989.
Datas de Publicação
-
Publicação nesta coleção
Jan-Apr 2015