Resumos
Durante o estudo dos metazoários parasitos de Paroaria dominicana Linnaeus, 1758 (Passeriformes), quatro infrapopulações de Platynosomum illiciens (Brown, 1901) (Trematoda) foram coletadas nos ductos hepáticos, com intensidades parasitárias de 10, 11, 18 e 36 espécimes respectivamente. Diferenças entre os valores morfométricos médios e o número total de ovos das infrapopulações foram estatisticamente testadas. O peso do fígado não se correlacionou ao peso das aves, ao comprimento do corpo das aves e à abundância parasitária. Foram detectadas correlações positivas entre o tamanho das infrapopulações e o número de ovos produzidos e entre o tamanho do corpo dos parasitos e o tamanho dos ovos. Correlações negativas foram observadas entre a intensidade parasitária e o peso do fígado; entre o tamanho das infrapopulações e o tamanho dos indivíduos dessas infrapopulações; entre o número e o tamanho dos ovos e entre o tamanho do corpo dos parasitos e o número de ovos. Os resultados indicaram que as infrapopulações maiores se estabeleceram em aves com fígados menores e que quanto maiores as infrapopulações foram observados menor tamanho corporal e menor tamanho dos ovos, os que se apresentaram em maior quantidade.
Ecologia parasitária; Galo-da-campina; helmintos; efeitos dependentes da densidade
During the study of the metazoan parasites of Paroaria dominicana Linnaeus, 1758 (Passeriformes), four infrapopulations of Platynosomum illiciens (Brown, 1901) (Trematoda), parasitic in the liver ducts were collected, with intensity of infection 10, 11, 18 and 36 specimens respectively. Differences between the means of morphometrical values and the total number of eggs were statistically tested. The liver weigh were not significantly correlated with avian total weigh and length, and with the total parasitic abundance. Positive correlations were observed between the size of infrapopulations and the total number of eggs, and between parasite body size and the eggs. size. Negative correlations were observed between parasitic intensity and liver weigh; between the size of infrapopulations and the parasite body size; between total number and the size of eggs and between parasite body size and total number of eggs. These results suggests that host with smaller liver had greater infrapopulations. These greater infrapopulations showed smaller parasite body size and higher size and number of eggs.
Parasite ecology; Red-cowlled cardinal; helminths; density dependent factors
ARTIGOS
Aspectos quantitativos e morfométricos das infrapopulações de platynosomum illiciens (trematoda: dicrocoeliidae) em paroaria dominicana (passeriformes: emberizidae) do estado da Bahia, Brasil* * Financiado em parte pelo CNPq.
Quantitative and morfometrical aspects in infrapopulations of platynosomum illiciens (trematoda: Dicrocoeliidae) in paroaria dominicana (passeriformes: emberizidae) from the state of Bahia, Brazil
Adriano R. CarvalhoI,* * Financiado em parte pelo CNPq. ; Sueli Souza-LimaII; Erik DaemonII; Luiz E.R. TavaresIII; José L. LuqueIII
ICurso de Pós-Graduação em Ciências Veterinárias, Universidade Federal Rural do Rio de Janeiro (UFRRJ), Km 7 da BR 465, Seropédica, RJ, 23890-000. Bolsista do CNPq. E-mail: arcarvalho@ufrrj.br
IIDepartamento de Zoologia, ICB, UFJF. Bolsista do CNPq
IIIDepartamento de Parasitologia Animal/IV/UFRRJ
RESUMO
Durante o estudo dos metazoários parasitos de Paroaria dominicana Linnaeus, 1758 (Passeriformes), quatro infrapopulações de Platynosomum illiciens (Brown, 1901) (Trematoda) foram coletadas nos ductos hepáticos, com intensidades parasitárias de 10, 11, 18 e 36 espécimes respectivamente. Diferenças entre os valores morfométricos médios e o número total de ovos das infrapopulações foram estatisticamente testadas. O peso do fígado não se correlacionou ao peso das aves, ao comprimento do corpo das aves e à abundância parasitária. Foram detectadas correlações positivas entre o tamanho das infrapopulações e o número de ovos produzidos e entre o tamanho do corpo dos parasitos e o tamanho dos ovos. Correlações negativas foram observadas entre a intensidade parasitária e o peso do fígado; entre o tamanho das infrapopulações e o tamanho dos indivíduos dessas infrapopulações; entre o número e o tamanho dos ovos e entre o tamanho do corpo dos parasitos e o número de ovos. Os resultados indicaram que as infrapopulações maiores se estabeleceram em aves com fígados menores e que quanto maiores as infrapopulações foram observados menor tamanho corporal e menor tamanho dos ovos, os que se apresentaram em maior quantidade.
Palavras-chave: Ecologia parasitária, Galo-da-campina, helmintos, efeitos dependentes da densidade.
ABSTRACT
During the study of the metazoan parasites of Paroaria dominicana Linnaeus, 1758 (Passeriformes), four infrapopulations of Platynosomum illiciens (Brown, 1901) (Trematoda), parasitic in the liver ducts were collected, with intensity of infection 10, 11, 18 and 36 specimens respectively. Differences between the means of morphometrical values and the total number of eggs were statistically tested. The liver weigh were not significantly correlated with avian total weigh and length, and with the total parasitic abundance. Positive correlations were observed between the size of infrapopulations and the total number of eggs, and between parasite body size and the eggs. size. Negative correlations were observed between parasitic intensity and liver weigh; between the size of infrapopulations and the parasite body size; between total number and the size of eggs and between parasite body size and total number of eggs. These results suggests that host with smaller liver had greater infrapopulations. These greater infrapopulations showed smaller parasite body size and higher size and number of eggs.
Key words: Parasite ecology, Red-cowlled cardinal, helminths, density dependent factors.
INTRODUÇÃO
Os mecanismos básicos que promovem a ocorrência de efeitos dependentes da densidade no crescimento e reprodução de helmintos geraram hipóteses que vão desde a competição, por exploração ou interferência, por recursos limitados, à possibilidade dos helmintos estimularem resposta imune mais drástica por parte dos hospedeiros (READ, 1951; HOLMES, 1961; 1962a; 1962b; BUSH; LOTZ, 2000; ROBERTS, 2000; LOWRIE et al., 2004). Outros autores afirmaram que diferenças na capacidade reprodutiva entre os parasitos e a heterogeneidade existente entre hospedeiros podem gerar enganos em relação aos mecanismos dependentes da densidade (KEYMER; SLATER, 1987; POULIN, 1995; 1996). No caso da reprodução, a densidadedependente pode ocorrer através dos efeitos diretos sobre a produção per capta de ovos ou, indiretamente, através da diminuição do crescimento, adiando a maturação e/ou reduzindo o período de vida do parasito (SHOSTAK; SCOTT, 1993). Outro fa-tor importante entre trematódeos co-específicos, é que o tamanho dos ovos varia consideravelmente de acordo com oscilações ambientais e relacionadas ao hospedeiro (POULIN; HAMILTON, 2000; THOMAS; POULIN, 2003).
Platynosomum illiciens (Brown, 1901) Kossack, 1910 tem sido registrado no Brasil, nos estados do Mato Grosso, Pará e Rio de Janeiro, parasitando ductos hepáticos de aves e mamíferos (RODRIGUES, 1963; TRAVASSOS et al., 1969). Estudos sobre aspectos infrapopulacionais de P. illiciens são escassos e importantes uma vez que indícios apontam este parasito como causador de hepatopatia em aves (KAZACOS et al., 1980).
Paroaria dominicana Linnaeus, 1758, conhecido vulgarmente como galo da campina, é um pássaro típico do interior do Nordeste do Brasil, ocorrendo do sul do Maranhão, Piauí e Ceará até o norte de Minas Gerais (RIDGELY; TUDOR, 1989). Vivem na mata baixa, rala e bem ensolarada (SICK, 1997; SILVA et al., 2003). São aves granívoras, de bico grosso e cônico, que forrageiam no solo, podendo ingerir pequenos artrópodes e moluscos (RIDGELY; TUDOR, 1989; SICK, 1997). Por suas qualidades canoras, tanto os machos quanto às fêmeas são disputados no mercado clandestino, o que faz com que sofram grande pressão de caçadores, sendo uma das espécies que mais movimenta o comércio ilegal de aves silvestres (SICK, 1997).
O presente trabalho teve como objetivo avaliar as possíveis implicações do aumento na intensidade do digenético P. illiciens nos ductos hepáticos de P. dominicana, sobre fatores relacionados ao crescimento e fecundidade dos parasitos que constituem as diferentes infrapopulações.
MATERIAL E MÉTODOS
Entre fevereiro e novembro de 2003, foram necropsiados 102 espécimes de P. dominicana, 48 fêmeas e 54 machos, cedidos mortos pelo Instituto do Meio Ambiente e Recursos Naturais Renováveis (IBAMA) de Juiz de Fora, MG, ao Departamento de Zoologia da Universidade Federal de Juiz de Fora, MG, oriundos do tráfico de aves silvestres, e de acordo com informações da apreensão, originários do estado da Bahia.
As aves foram medidas, pesadas e o fígado coletado, pesado em balança de precisão (magnificância = 10-4g) e examinado em microscópio estereoscópico para detecção da presença de digenéticos. Os parasitos encontrados foram coletados e processados segundo as técnicas propostas por Amato et al. (1991). Foi realizada a morfometria de 30% dos espécimes de cada infrapopulação, em microscópio Olympus BX-41, com o auxílio de ocular micrométrica. Na morfometria foram medidos os maiores comprimento e largura do corpo e comprimento e largura dos ovos. O tamanho (área) do corpo dos espécimes foi estimado conforme Poulin e Morand (1997) como o produto do maior comprimento pela largura máxima e o tamanho (área) dos ovos determinado como o produto do comprimento pela largura dos ovos (POULIN, 1997).
Para o estudo da fecundidade, foi realizada a contagem dos ovos em 30% dos indivíduos de cada infrapopulação, através do rompimento do corpo dos espécimes com o auxílio de estilete e posterior digestão tecidual de cada espécime da amostra em placa de Petri contendo 1ml de solução de ácido clorídrico (2%), por cinco minutos. Posteriormente, foi adicionado 3ml de água destilada a solução de HCl. Desta solução, após a homogeneização, foi retirado, com o auxílio de micropipeta volumétrica, 1ml (25%) para a montagem de lâminas, que foram levadas ao microscópio óptico para a contagem dos ovos. O número de ovos resultante da contagem de cada lâmina foi multiplicado por quatro (∑ nº de ovos x 4) e o resultado considerado como o número total de ovos do parasito.
Possíveis diferenças entre os valores médios do tamanho do corpo e a área e o número de ovos entre as infrapopulações de P. illiciens foram avaliadas com o teste de ANOVA um critério e com o teste a posteriori de Tukey para amostras de tamanhos diferentes, com os dados previamente transformados em log10(x+1).
O coeficiente de correlação Pearson (r) foi utilizado para determinar possíveis correlações entre o peso do fígado com o comprimento e o peso das aves; entre o tamanho das infrapopulações (= intensidade parasitária) e o tamanho do corpo, o tamanho e número dos ovos; entre o tamanho do corpo e a área e número de ovos, e entre o número e a área dos ovos (ZAR, 1999). O nível de significância estatística adotado foi P < 5%. A terminologia ecológica utilizada foi à proposta por Bush et al. (1997).
Três espécimes representativos de P. illiciens foram depositados na Coleção Helmintológica do Instituto Oswaldo Cruz (CHIOC), dois espécimes sob o número CHIOC 36648a e um sob o número CHIOC 36648b.
RESULTADOS
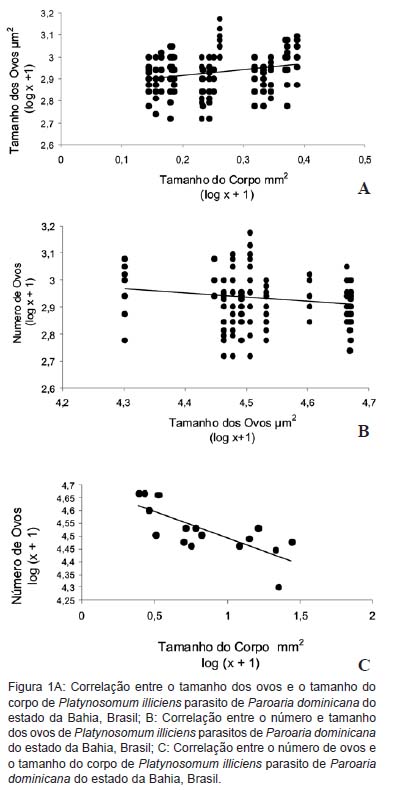
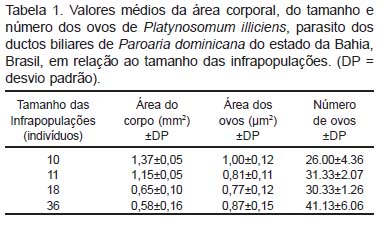
Foram coletadas quatro infrapopulações de P. illiciens parasitando os ductos hepáticos de P. dominicana; o tamanho dessas infrapopulações variou entre dez e 36 parasitos, totalizando 75 espécimes coletados, dos quais 25 foram utilizados para a morfometria. Os valores médios da área do corpo, do tamanho e número de ovos dos indivíduos de cada infrapopulação são apresentados na Tabela 1. Foram observadas diferenças significativas entre o tamanho do corpo e o tamanho e número de ovos dos indivíduos de P. illiciens que constituem as infrapopulações (Tabela 2).
Não foi verificada correlação entre o peso do fígado e o peso das aves (r = 0,12; P = 0,50) e nem entre do peso do fígado e o comprimento do corpo das aves (r = -0,11; P = 0,52). A abundância parasitária de P. illiciens não se correlacionou ao peso do fígado (rs= -0,09; P = 0,58), por outro lado, a intensidade parasitária correlacionou-se negativamente ao peso do fígado (rs = -1,00; P < 0,01). Foram observadas ainda correlações positivas significativas entre o tamanho das infrapopulações e o número de ovos produzidos (r = 0,76; P < 0,01) e entre o tamanho do corpo dos parasitos e o tamanho dos ovos (r = 0,28; P < 0,01); e correlação negativa entre o tamanho das infrapopulações e o tamanho dos indivíduos que formam essas infrapopulações (r = -0,81; P < 0,01), entre o número e o tamanho dos ovos (r = -0,19; P < 0,01) e entre o tamanho do corpo dos parasitos e o número de ovos (r = -0,77; P < 0,01) (Figura 1).
DISCUSSÃO
Os resultados do presente estudo indicam a ocorrência de variações no tamanho e número dos ovos e no tamanho dos indivíduos em função do número de espécimes que constituem as diferentes infrapopulações de P. illiciens nos ductos hepáticos de P. dominicana. Com aumento no número de indivíduos que formam as infrapopulações, ocorreu diminuição no tamanho do corpo, redução do tamanho dos ovos e aumento no número destes ovos. Estes resultados parecem apontar para a ocorrência de efeitos dependentes da densidade, o que para Bush e Lotz (2000), poderia ocorrer em virtude da associação intraespecifica que se estabelece entre os constituintes das infrapopulações, afirmando que a aglomeração de indivíduos num mesmo sítio promove o aumento da resistência ambiental, representado por grande número de efeitos negativos, como a falta de alimento, de espaço para a reprodução, de oxigênio, acúmulo de dejetos metabólicos e a severidade da resposta imune do hospedeiro. Assim, em virtude da limi-tação ambiental, vários podem ser os efeitos dependentes da densidade, principalmente relacionados ao crescimento e a capacidade reprodutiva de helmintos, conforme já reportado para outras espécies de populações naturalmente infectadas (ANDERSON; MAY, 1978; KEYMER, 1982). Além disso, Keymer (1982) sugeriu que os efeitos dependentes da densidade estão relacionados com o padrão agregado de distribuição dos parasitos na população de hospedeiros e que estes devem ser reconhecidos como importantes fatores relacionados à biologia da população de parasitos, prevenindo um aumento exponencial da população pelos efeitos que exerce sobre o estabelecimento, reprodução ou sobrevivência dos parasitos em função do número de outros indivíduos no mesmo hospedeiro.
Em organismos de vida livre, condições de alta densidade favorecem organismos com potencial de crescimento menor, mas com melhores capacidades para utilizar e competir por recursos escassos (ODUM, 1988). Já os parasitos, muitas vezes referidos como exemplos perfeitos de organismos r-selecionados, possuem diversas características finais de estratégia de historia de vida r, tais como período de vida curto, maturidade precoce, corpo pequeno, e prolíficos produtores de ovos pequenos (PIANKA, 1970). Os resultados do presente trabalho apontam para o fato de que aumentando a intensidade de P. illiciens nos ductos biliares ocorreu uma mudança na estratégia reprodutiva destes parasitos, que aumentam a produção de ovos, podendo indicar uma resposta estado-dependente buscando alcançar maior capacidade de infecção. De acordo com Price (1974), a competição entre os parasitos, nas fases de dispersão e residência, é um dos fatores que definem a estratégia na produção de ovos pelos parasitos. Assim, a alta capacidade de produção de ovos, como resultado de pressões seletivas, surge da necessidade de compensar a baixa eficiência de transmissão, relacionada ao ciclo de vida dos helmintos (PRICE, 1974; KEYMER, 1982), onde a probabilidade de sobrevivência e de infecção do hospedeiro durante as fases de transmissão do ciclo de vida, e a disponibilidade de nutrientes ou outros recursos para a reprodução dos adultos, são suspeitos de terem dirigido a evolução de estratégias reprodutivas de parasitos (PRICE, 1974; JENNINGS; CALOW, 1975; CALLOW, 1983).
Com a diminuição do corpo e o aumento do número de ovos, surge o efeito da repartição ótima de recursos entre essas duas características da bionomia do parasito. Assim, em trabalho anterior, realizado com uma espécie de Schistosoma parasito de mamíferos, foi detectada relação inversa entre o número e o tamanho dos ovos (LOKER, 1983). Estudos realizados com Fasciola hepatica de bovinos e ovinos, e com Schistosoma mansoni de humanos observaram que a fecundidade desses parasitos variou inversamente com a densidade dos mesmos no fígado de seus hospedeiros, além de não ter sido demonstrado variação no tamanho dos ovos (SMITH, 1987). Os dados obtidos no presente trabalho demonstraram que com o aumento da densidade dos parasitos foi acompanhado pela diminuição do tamanho do corpo e aumento da fecundidade, porém, com diminuição do número de ovos. Os resultados observados, em concordância com os obtidos em outros trabalhos, parecem indicar a ocorrência de interferências entre as características da história de vida, além de apontar para a diversidade de respostas estado-dependentes de acordo com as pressões seletivas pelas quais as diferentes espécies estiveram submetidas.
Nas aves estudadas, o peso do fígado não se correlacionou ao peso e nem ao comprimento das aves, o que pode indicar que o desenvolvimento do fígado ocorre de forma independente do desenvolvimento corporal.
Os dados obtidos no presente trabalho parecem apontar para a ocorrência de efeitos dependentes da densidade. No entanto, foram observadas apenas quatro infrapopulações, nas quais a intensidade não variou de maneira acentuada. Para resultados mais conclusivos, são necessários estudos experimentais que permitam controlar os níveis de infecção em um grupo de hospedeiros e obter uma grande amplitude de intensidade.
Agradecimentos:-Ao Instituto do Meio Ambiente e Recursos Naturais Renováveis de Juiz de Fora MG, pela concessão das aves. Ao Departamento de Zoologia, ICB, UFJF e ao Departamento de Parasitologia Animal, IV, UFRRJ, pela infra-estrutura disponibilizada.
Recebido em 19 de setembro de 2006
Aceito para publicação em 08 de dezembro de 2007.
- ANDERSON, R. M.; MAY, R. M. Regulation and stability of host-parasite population interactions: I. Regulatory processes. Journal of Animal Ecology, v.47, n. 1, p. 219-248, 1978.
- AMATO, J. F. R; BOEGER, W. A.; AMATO, S. P. Protocolos para Laboratório, coleta e processamento de parasitos de pescado. Seropédica: UFRRJ, Imprensa Universitária, 1991. 81p.
- BUSH, A. O.; LAFFERTY, K. D.; LOTZ, J. M.; SHOSTAK, A. W. Parasitology meets Ecology on its onw Terms: Margolis et al. revisited. Journal of Parasitology, v. 83, n. 4, p. 575-583, 1997.
- BUSH, A. O.; LOTZ, J. M. The ecology of "crowding". Journal of Parasitology, v. 86, n. 2, p. 212-213, 2000.
- CALOW, P. Pattern and paradox in parasite reproduction.Parasitology, v. 86, n. 4, p. 197-207, 1983.
- HOLMES, J. C. Effects of concurrent infections on Hymenolepis diminuta (Cestoda) and Moniliformis dubius (Acanthocephala). I. General effects and comparison with crowding. Journal of Parasitology, v. 47, n. 2, p. 209-216, 1961.
- HOLMES, J. C. Effects of concurrent infections on Hymenolepis diminuta (Cestoda) and Moniliformis dubius (Acanthocephala). II. Effects on growth. Journal of Parasitology, v. 48, n. 1, p. 87-96, 1962a.
- HOLMES, J. C. Effects of concurrent infections on Hymenolepis diminuta (Cestoda) and Moniliformis dubius (Acanthocephala). III. Effects in Hamsters. Journal of Parasitology, v. 48, n. 1, p. 97-100, 1962b.
- JENNINGS, J. B.; CALOW, P. The relationship between high fecundity and the evolution of entoparasitism. Oecologia, v. 21, n. 1, p. 109-115, 1975.
- KAZACOS, K. R.; DHILLON, A. S.; WINTERFIELD, R. W.; THACKER, H. L. Fatal hepatic trematodiasis in cockatoos due to Platynosomum proxillicens Avian Diseases, v. 24, n.3, p. 788-793, 1980.
- KEYMER, A. Density-dependent mechanisms in the regulation of intestinal helminth populations. Parasitology, v. 84, n. 6, p. 573-587, 1982.
- KEYMER, A. E.; SLATER, A. F. G. Helminth fecundity: density dependence or statistical illusion? Parasitology Today, v. 3, n. 2, p. 56-58, 1987.
- LOKER, E.S.A comparative study of the life-histories of mammalian schistosomes. Parasitology, v. 87, n. 3, p. 343369, 1983.
- LOWRIE, F. M.; BEHNKE, J. M.; BARNARD, C. J. Density-dependent effects on the survival and growth of the rodent stomach worm Protospirura muricola in laboratory mice. Journal of Helminthology, v. 78, n. 2, p. 121-128, 2004.
- ODUM, E. P. Ecologia Trad. Christopher J. Tribe. Rio de Janeiro: Guanabara Koogan, 1988. 438 p.
- PIANKA, E. R. On r- and k-selection. American Naturalist, v. 104, n. 927, p. 592-597, 1970.
- POULIN, R. Evolution of parasite life-history traits: myths and reality. Parasitology Today, v. 11, n. 9, p. 342-345, 1995.
- POULIN, R. The evolution of life history strategies in parasitic animals. Advances in Parasitology, v. 37, p. 107-134, 1996.
- POULIN, R. Egg production in adult trematodes: adaptation or constraint? Parasitology, v. 114, n. 2, p. 195-204, 1997.
- POULIN, R.; HAMILTON, W. Egg size variation as a function of environmental variability in parasitic trematodes. Canadian Journal of Zoology, v. 78, n. 4, p. 564-569, 2000.
- POULIN, R.; MORAND, S. Parasite body size distributions: interpreting patterns of skewness. International Journal for Parasitology, v. 27, n. 8, p. 959-964, 1997.
- PRICE, P. W. Strategies for egg production. Evolution, v. 28, n. 1, p. 76-84, 1974.
- READ, C. P. The "crowding effect" in tapeworm infections. Journal of Parasitology, v. 37, n. 1, p. 174-178, 1951.
- RIDGELY, R. S.; TUDOR, G. The Birds of South America. Austin: University of Texas Press, 1989. p. 516.
- ROBERTS, L. S. The crowding effect revisited. Journal of Parasitology, v. 86, n. 2, p. 209-211, 2000.
- RODRIGUES, H. O. Contribuição estudo do gênero Platynosomum Looss, 1907 (Trematoda: Dicrocoeliidae). Memórias do Instituto Oswaldo Cruz, v. 61, n. 3, p. 507515, 1963.
- SHOSTAK, A. W.; SCOTT, M. E. Detection of density-dependent growth and fecundity of helminths in natural infections. Parasitology, v. 106, n. 4, p. 527-539, 1993.
- SICK, H. Ornitologia Brasileira Rio de Janeiro: Nova Fronteira, 1997. p. 912.
- SILVA, J. M. C.; SOUZA, M. A.; BIEBER, A. G. D.; CARLOS, C. J. Aves da caatinga: status, uso do habitat e sensitividade. In: LEAL, I. R.; TABARELLI, M.; SILVA, J. M. C. (Eds), Ecologia e Conservação da Caatinga Recife: Editora Universitária UFPE, 2003. p. 239-274.
- SMITH, G. The regulation of trematode populations by density-dependent processes. International Journal for Parasitology, v. 17, n. 2, p. 541-548, 1987.
- THOMAS, F.; POULIN, R. Egg size variability in trematodes: test of the bet-hedging hypotesis. Journal of Parasitology, v. 89, n. 6, p. 1159-1162, 2003.
- TRAVASSOS, L.; FREITAS, J. F. T.; KOHN, A. Trematódeos do Brasil. Memórias do Instituto Oswaldo Cruz, v. 67, n. 1, p. 1-885, 1969.
- ZAR, J. H. Biostatistical Analysis Upper Sadle River: Prentice-Hall Press, 1999. 663 p.
Datas de Publicação
-
Publicação nesta coleção
12 Mar 2012 -
Data do Fascículo
Dez 2007
Histórico
-
Aceito
08 Dez 2007 -
Recebido
19 Set 2006