Abstracts
During the study of the metazoan parasites of Paroaria dominicana (Linnaeus, 1758), eight infrapopulations of Mediorhynchus emberizae (Rudolphi, 1819), were collected in the medium third of the small intestine, with parasite intensities ranging from one to ten specimens. Differences among the average values of the biomass, volume of the eggs, volume of the testicles and the parasite density of the infrapopulations were statistically tested in order to detect variations of these parameters in function of parasite density. Was observed that the acanthocephalans biomass increased with the volume of the intestine and also that the parasite density increased with the parasite intensity in the infrapopulations. It was verified that the increase of the parasite intensity was accompanied by the decrease of the mean biomass of the parasites. Decrease of the volume of the eggs was verified with the increase of the density and of the parasite intensity. The results of the present work could suggested the occurrence of density-dependent factors and the decrease of testicles volume and the biomass of the males with the increase of the parasite density in the infrapopulations of M. emberizae in the intestine of P dominicana.
Red-cowled cardinals; acanthocephalans; biomass; Brazil
Durante o estudo dos metazoários parasitos de Paroaria dominicana (Linnaeus, 1758), oito infrapopulações de Mediorhynchus emberizae (Rudolphi, 1819), foram coletadas no terço médio do intestino delgado, com intensidades parasitárias variando de um a dez espécimes. Diferenças entre os valores médios da biomassa, volume dos ovos, volume dos testículos e da densidade parasitária das diferentes infrapopulações foram estatisticamente testadas com o objetivo de detectar variações desses parâmetros em função da densidade parasitária. Foi observado que a biomassa de acantocéfalos aumentou com o volume do intestino e também que a densidade dos parasitos aumentou com a intensidade parasitária nas infrapopulações. Verificou-se que o aumento da intensidade parasitária foi acompanhada pela diminuição da biomassa média dos parasitos. Constatou-se diminuição do volume dos ovos com o aumento da densidade e da intensidade parasitária. Os resultados do presente trabalho parecem dar indícios da ocorrência de fatores dependentes da densidade e da diminuição do volume testicular e a biomassa dos machos com o aumento da densidade parasitária nas infrapopulações de M. emberizae no intestino de P. dominicana.
Galo da campina; acantocéfalos; biomassa; Brasil
NOTA DE PESQUISA
Relação entre biomassa e densidade parasitária de Mediorhynchus emberizae (acanthocephala: gigantorhynchidae) parasito de Paroaria dominicana (passeriformes: emberizidae) do estado da Bahia, Brasil
Relationship between biomass and parasite density of Mediorhynchus emberizae (Acanthocephala: Gigantorhynchidae) parasites of Paroaria dominicana (Passeriformes: Emberizidae) of the State of Bahia, Brazil
Adriano R. CarvalhoI, * * Bolsista CNPq ** Pesquisador CNPq. E-mails: lertavares@gmail.com e jlluque@ufrrj.br ; Sueli Souza-LimaII; Luiz E. R. TavaresIII, * * Bolsista CNPq ** Pesquisador CNPq. E-mails: lertavares@gmail.com e jlluque@ufrrj.br ; José L. LuqueIII, ** * Bolsista CNPq ** Pesquisador CNPq. E-mails: lertavares@gmail.com e jlluque@ufrrj.br
ICurso de Pós-Graduação em Ciências Veterinárias, Universidade Federal Rural do Rio de Janeiro (UFRRJ). E-mail: arcarvalho@ufrrj.br
IIDepartamento de Zoologia, Universidade Federal de Juiz de Fora (UFJF). E-mail: souzalima@acessa.com
IIIDepartamento de Parasitologia Animal, Universidade Federal Rural do Rio de Janeiro (UFRRJ), BR 465, Km 7, Caixa Postal 74508, Seropédica, RJ 23890-000
RESUMO
Durante o estudo dos metazoários parasitos de Paroaria dominicana (Linnaeus, 1758), oito infrapopulações de Mediorhynchus emberizae (Rudolphi, 1819), foram coletadas no terço médio do intestino delgado, com intensidades parasitárias variando de um a dez espécimes. Diferenças entre os valores médios da biomassa, volume dos ovos, volume dos testículos e da densidade parasitária das diferentes infrapopulações foram estatisticamente testadas com o objetivo de detectar variações desses parâmetros em função da densidade parasitária. Foi observado que a biomassa de acantocéfalos aumentou com o volume do intestino e também que a densidade dos parasitos aumentou com a intensidade parasitária nas infrapopulações. Verificou-se que o aumento da intensidade parasitária foi acompanhada pela diminuição da biomassa média dos parasitos. Constatou-se diminuição do volume dos ovos com o aumento da densidade e da intensidade parasitária. Os resultados do presente trabalho parecem dar indícios da ocorrência de fatores dependentes da densidade e da diminuição do volume testicular e a biomassa dos machos com o aumento da densidade parasitária nas infrapopulações de M. emberizae no intestino de P. dominicana.
Palavras-chave: Galo da campina, acantocéfalos, biomassa, Brasil.
ABSTRACT
During the study of the metazoan parasites of Paroaria dominicana (Linnaeus, 1758), eight infrapopulations of Mediorhynchus emberizae (Rudolphi, 1819), were collected in the medium third of the small intestine, with parasite intensities ranging from one to ten specimens. Differences among the average values of the biomass, volume of the eggs, volume of the testicles and the parasite density of the infrapopulations were statistically tested in order to detect variations of these parameters in function of parasite density. Was observed that the acanthocephalans biomass increased with the volume of the intestine and also that the parasite density increased with the parasite intensity in the infrapopulations. It was verified that the increase of the parasite intensity was accompanied by the decrease of the mean biomass of the parasites. Decrease of the volume of the eggs was verified with the increase of the density and of the parasite intensity. The results of the present work could suggested the occurrence of density-dependent factors and the decrease of testicles volume and the biomass of the males with the increase of the parasite density in the infrapopulations of M. emberizae in the intestine of P dominicana.
Key words: Red-cowled cardinals, acanthocephalans, biomass, Brazil.
Os mecanismos básicos que promovem a ocorrência de efeitos dependentes da densidade no crescimento e reprodução de helmintos geraram hipóteses que vão desde a competição, por exploração ou interferência, por recursos limitados, à possibilidade dos helmintos estimularem resposta imune mais drástica por parte dos hospedeiros (BUSH; LOTZ, 2000; ROBERTS, 2000; LOWRIE et al., 2004). Diversos trabalhos realizados sobre efeitos dependentes da densidade parasitária têm utilizado a variação da intensidade parasitária nas infrapopulações, relacionada às características do hospedeiro, como peso e comprimento, ou mesmo usando apenas cada hospedeiro como uma unidade amostral independente (ARNEBERG et al., 1998; DEZFULI et al., 2002, LOWRIE et al., 2004). No entanto, a utilização da biomassa de parasitos, em função da área ou volume efetivamente ocupado no hospedeiro tem sido apontada como uma medida mais robusta, visto que o suprimento de energia, espaço físico e efeito do hospedeiro são compartilhados e assim, podem determinar como a biomassa parasitária pode ser sustentada, além de fornecer indícios de como ocorre o gerenciamento da energia entre os diversos aspectos da bionomia dos parasitos, como o crescimento e o tamanho dos ovos (POULIN; GEORGE-NASCIMENTO, 2007).
O presente trabalho teve como objetivo avaliar as possíveis implicações do aumento na densidade do acantocéfalo Mediorhynchus emberizae no intestino delgado de Paroaria dominicana, vulgarmente conhecida como galo da campina, uma ave granívora típica do interior do Nordeste do Brasil, sobre fatores relacionados à biomassa e volume dos ovos produzidos pelas fêmeas e biomassa e massa testicular dos machos, que constituem as diferentes infrapopulações. Entre fevereiro e novembro de 2003, foram necropsiados 102 espécimes de P. dominicana, 48 fêmeas e 54 machos, cedidos mortos pelo Instituto do Meio Ambiente e Recursos Naturais Renováveis (IBAMA) Juiz de Fora, MG ao Departamento de Zoologia da Universidade Federal de Juiz de Fora, MG, apreendidos do tráfico de aves silvestres, e originários do estado da Bahia. Todas as aves foram medidas e pesadas. Todos os órgãos foram observados em microscópio estereoscópico para a coleta de helmintos. O intestino delgado foi dividido em três regiões, terço anterior, terço médio e terço posterior, sendo que para cada uma foram medidos o comprimento e a largura. Os acantocéfalos encontrados foram coletados e processados segundo as técnicas compiladas em Amato et al. (1991). Foi realizada a morfometria de todos dos espécimes de cada infrapopulação, em microscópio Olympus BX-41, com o auxílio de ocular micrométrica. Na morfometria foram medidos o comprimento e largura do corpo e comprimento e largura de 20 ovos de cada fêmea adulta grávida. Os ovos selecionados para serem medidos foram aqueles localizados no interior do sino uterino; quando não era alcançado o número de 20 ovos, foram medidos ovos próximos a esta localização.
O biovolume ou volume corporal, reconhecido como o espaço ocupado pelo corpo dos acantocéfalos, foi calculado para cada um dos espécimes de todas as infrapopulações de M. emberizae em P. dominicana. O volume corporal de cada espécime de parasito e da região do intestino delgado foram aproximados, assumindo a forma de um cilindro perfeito, pela fórmula πCL2/4; o volume dos ovos e dos testículos, de formato elipsóide, foi calculado por πCL2/6, onde C é o comprimento e L a largura dos ovos (POULIN; MORAND, 2000; POULIN et al., 2003). Abiomassa dos parasitos, que no presente trabalho é reconhecida como a quantidade total de material que forma o corpo dos acantocéfalos, foi indicada pelo biovolume, obedecendo a relação 1cm3 = 1g (POULIN; GEORGE-NASCIMENTO, 2007). O volume médio dos ovos é fornecido em x10-2mm3 e a biomassa dos testículos em x10-2g. Abiomassa total de cada infrapopulação foi obtida pelo somatório da biomassa de cada parasito e a densidade parasitária foi obtida pela razão entre a biomassa total da infrapopulação pelo volume do terço do intestino delgado onde se localizavam, e o resultado fornecido em x10-5g/mm3.
As fêmeas de M. emberizae foram classificadas em adultas não grávidas, que apresentam apenas bolas ovarianas, e adultas grávidas, que contém uma mistura de bolas ovarianas e embriões em desenvolvimento e plenamente desenvolvidos (AZNAR et al., 2001). O teste t de "student" foi utilizado para comparar a biomassa das fêmeas adultas não grávidas com o das fêmeas adultas e grávidas, entre as infrapopulações com o mesmo número de fêmeas. Os machos de M. emberizae foram tratados separadamente para a observação do efeito do aumento das infrapopulações sobre a biomassa e a massa testicular. Amassa testicular é representada pela soma das massas dos testículos anterior e posterior (POULIN; MORAND, 2000). A contribuição da massa dos testículos na biomassa dos parasitos foi medida pela entre a massa testicular e a biomassa do corpo. Possíveis diferenças entre os valores médios da biomassa total e o volume dos ovos entre as infrapopulações de M. emberizae foram avaliadas com o teste de ANOVAum critério e com o teste a posteriori de Tukey para amostras de tamanhos diferentes, com os dados previamente transformados em log10(x + 1). O coeficiente de correlação de Pearson (r) foi utilizado para determinar possíveis correlações entre o volume do intestino com o comprimento e o peso das aves; entre o tamanho das infrapopulações e o peso dos hospedeiros, a biomassa total, densidade parasitária, o volume dos ovos; entre a biomassa dos parasitos e o volume intestinal; entre o número de fêmeas na infrapopulação e a biomassa dos machos e entre a densidade parasitária e o volume médio dos ovos (ZAR, 1996). O nível de significância estatística adotado foi P < 0.05. A terminologia ecológica utilizada foi a proposta por Bush et al. (1997). Espécimes de M. emberizae foram depositados na Coleção Helmintológica do Instituto Oswaldo Cruz (CHIOC), CHIOC 36649a macho, e CHIOC 36649b fêmea.
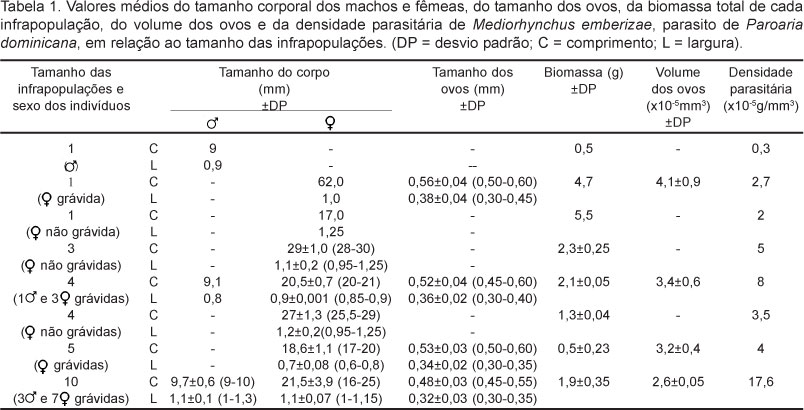
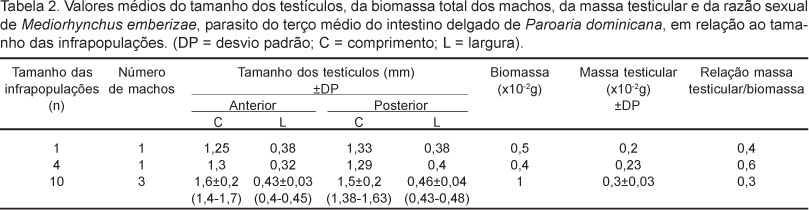
Foram coletadas oito infrapopulações de M. emberizae parasitando o terço médio do intestino delgado de P. dominicana não havendo a co-ocorrência de outra espécie de helminto. O tamanho dessas infrapopulações variou entre um e dez parasi-tos. Foram coletados e medidos um total de 29 espécimes, dos quais 24 fêmeas e cinco machos. Os valores médios do tamanho dos espécimes, da biomassa e do tamanho e volume dos ovos, além da densidade parasitária de cada infrapopulação são apresentados na Tabela 1. O peso médio das aves parasitadas foi 22,89 ± 3,46g (16,40 - 26,70g), o terço médio do intestino delgado mediu 5,4 ± 0,7cm (4,6-6,3cm) de comprimento e 0,53 ± 0,09cm (0,4-0,6cm) e o volume intestinal médio foi 1163,45 ± 380,19mm3 (791,57 - 1441,95mm3). Foram observadas diferenças significativas entre a biomassa total dos indivíduos de M. emberizae que constituem as infrapopulações (F4,455 = 332,65; P < 0,01) e entre o volume dos ovos das fêmeas que constituem as diferentes infrapopulações (F3,296= 403,26; P < 0,01). As fêmeas adultas não grávidas apresentaram biomassa maior do que aquelas adultas grávidas, comparando as infrapopulações com uma e com três fêmeas (t = -6,05; P < 0,01). Foram obtidos machos em três infrapopulações, onde observou-se que os valores da biomassa e da massa testicular aumentaram com o aumento do número de fêmeas das infrapopulaões e que nos machos maiores a contribuição da massa testicular na biomassa total dos parasitos foi menor (Tabela 2). Foi verificado ainda que quanto maior o número de fêmeas na infrapopulação maior foi a biomassa dos machos de M. emberizae (r = 0,90; P = 0,03).
O volume intestinal não foi correlacionado com o peso das aves (r = 0,09; P = 0,89), no entanto, quanto maior o volume intestinal, maior foi a biomassa de parasitos verificada (r = 0,75; P < 0,01). A densidade parasitária aumentou com o tamanho das infrapopulações (r = 0,94; P < 0,01). Verificaram-se correlações negativas entre o tamanho das infrapopulações e a biomassa média dos parasitos (r = -0,15; P < 0,01); entre o volume médio dos ovos e o tamanho das infrapopulações (r = -0,62; P < 0,01), e a densidade parasitária (r = -0,59; P < 0,01).
Os resultados obtidos no presente trabalho apontam para o fato de que o aumento da intensidade parasitária nas infrapopulações de M. emberizae em P. dominicana determinou o aumento da densidade parasitária, tendo como reflexos a diminuição da biomassa dos parasitos e também a diminuição no volume dos ovos. Considerando a biomassa de parasitos no hospedeiro, como um reflexo da sua demanda energética (GEORGE-NASCIMENTO et al., 2002), pode-se levantar a possibilidade da ocorrência de efeitos dependentes da densidade, uma vez que com o aumento da densidade parasitária, passa a ocorrer uma maior competição pelos recursos energéticos não só entre os parasitos, mas também com relação ao hospedeiro. Esse fato é reforçado pela posição dos acantocéfalos adultos na região média do intestino delgado, onde absorvem os nutrientes, resultantes da digestão do hospedeiro, através da superfície do corpo (CROMPTON, 1973), o que pode influenciar negativamente a biomassa do hospedeiro, fato que foi observado no presente trabalho.
Quando comparado a biomassa de fêmeas grávidas e não grávidas, em infrapopulações de um e três espécimes, as primeiras apresentaram-se menores, podendo sugerir uma possível ocorrência de "trade-off' entre algumas características bionômicas, tais como crescimento e fecundidade (POULIN, 2007). Dessa forma, entre os acantocéfalos, as fêmeas grávidas devem fazer o repartimento da energia disponível entre o desenvolvimento corporal, que permite uma maior capacidade de competição e na aquisição de recursos, e a produção de ovos, visto que ovos mais volumosos podem obter maior sucesso em infecções futuras (POULIN et al., 2003). Por outro lado, as fêmeas não grávidas podem alocar grande parte dos recursos para o desenvolvimento corporal, o que poderia gerar espécimes com maior biomassa. Outro fator que poderia estar influenciando esses resultados é o fato de que as fêmeas não grávidas ocorreram em menor densidade parasitária (Tabela 2), o que poderia contribuir para maior biomassa apresentada. Quando considerados isoladamente os machos, os resultados do presente trabalho indicaram que nos machos maiores, a massa tes-ticular teve menor contribuição na biomassa total do helminto, indicando não ocorrer o crescimento alométrico dos testículos em relação ao corpo dos machos. Embora, devido ao tamanho da amostra, esta seja uma observação preliminar, esse fato pode sugerir que o investimento relativo no crescimento dos testículos diminuiu com o aumento do número de fêmeas.
Os resultados do presente trabalho parecem dar indícios da ocorrência de fatores dependentes da densidade e da diminuição da razão entre o volume testicular e a biomassa corporal dos machos com o aumento da densidade parasitária nas infrapopulações de M. emberizae no intestino de P. dominicana.
REFERÊNCIAS BIBLIOGRÁFICAS
AMATO, J.F.R; BOEGER, W.A.; AMATO, S.P. Protocolos para Laboratório, coleta e processamento de parasitos de pescado. Seropedica: UFRRJ, Imprensa Universitária, 1991. 81p.
ARNEBERG, P.; SKORPING, A.; READ, A. F. Parasite abundance, body size, life histories, and the energetic equivalence rule. American Naturalist, v. 151, n. 6, p. 497513, 1998.
AZNAR, F. J.; BUSH, A. O.; BALBUENA, J. A.; RAGA, J. A. Corynosoma cetaceum in the stomach of franciscanas, Pontoporia blainvillei (Cetacea): an exceptional case of habitat selection by an acanthocephalan. Journal of Parasitology, v. 87, n. 3, p. 536-541, 2001.
BUSH, A.O.; LAFFERTY, K.D.; LOTZ, J.M.; SHOSTAK, A.W. Parasitology meets Ecology on its onw Terms: Margolis et al. revisited. Journal of Parasitology, v. 83, n. 4, p. 575-583, 1997.
BUSH, A. O.; LOTZ, J. M. 2000. The ecology of "crowding". Journal of Parasitology, v. 86, n. 2, p. 212-213, 2000. CROMPTON, D. W. T. The sites occupied by some parasitic helminths in the alimentary tract of vertebrates. Biological Reviews of the Cambridge Philosophical Society, v. 48, n. 1, p. 27-83, 1973.
DEZFULI, B. S.; VOLPONI, S.; BELTRAMI, L.; POULIN, R. Intra- and interspecific density-dependent effects on growth in helminth parasite of the cormorant, Phalacrocorax carbo sinensis. Parasitology, v. 124, n. 5, p. 537-544, 2002.
GEORGE-NASCIMENTO, M.; GARCÍAS, F.; MUNOZ, G. Parasite body volume and infracommunity patterns in the southern pomfret Brama australis (Pisces: Bramidae). Revista Chilena de História Natural, v. 75, n. 4, p. 835-839, 2002.
LOWRIE, F. M.; BEHNKE, J. M.; BARNARD, C. J. Densitydependent effects on the survival and growth of the rodent stomach worm Protospirura muricola in laboratory mice. Journal of Helminthology, v. 78, n. 2, p. 121-128, 2004.
POULIN, R. Evolutionary Ecology of Parasites. 2nd. Princeton: Princeton University Press, 2007. 332 p
POULIN, R.; MORAND, S. Testes size, body size and male-male competition in acanthocephalan parasites. Journal of Zoology, v. 250, n. 4, p. 551-558, 2000.
POULIN, R.; WISE, M.; MOORE, J. Acomparative analysis of adult body size and its correlates in acanthocephalan parasites. International Journal for Parasitology, v. 33, n. 8, p. 799-805, 2003.
POULIN, R.; GEORGE-NASCIMENTO, M. The scaling of total parasite biomass with host body mass. International Journal for Parasitology, v. 37, n. 3-4, p. 359-364, 2007.
ROBERTS, L. S. The crowding effect revisited. Journal of Parasitology, v. 86, n. 2, p. 209-211, 2000.
ZAR, J. H. Biostatistical Analysis. Upper Saddle River: Prentice-Hall, 1996. 662 p.
Recebido em 09 de outubro de 2007.
Aceito para publicação em 15 de abril de 2008.
- AMATO, J.F.R; BOEGER, W.A.; AMATO, S.P. Protocolos para Laboratório, coleta e processamento de parasitos de pescado. Seropedica: UFRRJ, Imprensa Universitária, 1991. 81p.
- ARNEBERG, P.; SKORPING, A.; READ, A. F. Parasite abundance, body size, life histories, and the energetic equivalence rule. American Naturalist, v. 151, n. 6, p. 497513, 1998.
- AZNAR, F. J.; BUSH, A. O.; BALBUENA, J. A.; RAGA, J. A. Corynosoma cetaceum in the stomach of franciscanas, Pontoporia blainvillei (Cetacea): an exceptional case of habitat selection by an acanthocephalan. Journal of Parasitology, v. 87, n. 3, p. 536-541, 2001.
- BUSH, A.O.; LAFFERTY, K.D.; LOTZ, J.M.; SHOSTAK, A.W. Parasitology meets Ecology on its onw Terms: Margolis et al. revisited. Journal of Parasitology, v. 83, n. 4, p. 575-583, 1997.
- BUSH, A. O.; LOTZ, J. M. 2000. The ecology of "crowding". Journal of Parasitology, v. 86, n. 2, p. 212-213, 2000.
- CROMPTON, D. W. T. The sites occupied by some parasitic helminths in the alimentary tract of vertebrates. Biological Reviews of the Cambridge Philosophical Society, v. 48, n. 1, p. 27-83, 1973.
- DEZFULI, B. S.; VOLPONI, S.; BELTRAMI, L.; POULIN, R. Intra- and interspecific density-dependent effects on growth in helminth parasite of the cormorant, Phalacrocorax carbo sinensis. Parasitology, v. 124, n. 5, p. 537-544, 2002.
- GEORGE-NASCIMENTO, M.; GARCÍAS, F.; MUNOZ, G. Parasite body volume and infracommunity patterns in the southern pomfret Brama australis (Pisces: Bramidae). Revista Chilena de História Natural, v. 75, n. 4, p. 835-839, 2002.
- LOWRIE, F. M.; BEHNKE, J. M.; BARNARD, C. J. Densitydependent effects on the survival and growth of the rodent stomach worm Protospirura muricola in laboratory mice. Journal of Helminthology, v. 78, n. 2, p. 121-128, 2004.
- POULIN, R. Evolutionary Ecology of Parasites. 2nd. Princeton: Princeton University Press, 2007. 332 p
- POULIN, R.; MORAND, S. Testes size, body size and male-male competition in acanthocephalan parasites. Journal of Zoology, v. 250, n. 4, p. 551-558, 2000.
- POULIN, R.; WISE, M.; MOORE, J. Acomparative analysis of adult body size and its correlates in acanthocephalan parasites. International Journal for Parasitology, v. 33, n. 8, p. 799-805, 2003.
- POULIN, R.; GEORGE-NASCIMENTO, M. The scaling of total parasite biomass with host body mass. International Journal for Parasitology, v. 37, n. 3-4, p. 359-364, 2007.
- ROBERTS, L. S. The crowding effect revisited. Journal of Parasitology, v. 86, n. 2, p. 209-211, 2000.
- ZAR, J. H. Biostatistical Analysis. Upper Saddle River: Prentice-Hall, 1996. 662 p.
Publication Dates
-
Publication in this collection
11 May 2012 -
Date of issue
June 2008
History
-
Accepted
15 Apr 2008 -
Received
09 Oct 2007